Introduzione al sistema endocrino
Lezione 2
Prof. Manasseri.
Sbob. Siciliano Enrico
5/03/2013
Sia il sistema nervoso, che il sistema endocrino, che il sistema immunitario svolgono le loro funzioni di controllo, di integrazione, e
di coordinazione mediante sostanze chimiche. Perché anche se il sistema nervoso dialoga attraverso l’impulso, che è un evento
elettrico, alla fine il segnale verrà trasmesso agli effettori tramite delle sostanze chimiche, che si chiamano neurotrasmettitori.
Innanzitutto parliamo dei neurotrasmettitori. Sono liberati dai neuroni a livello sinaptico (la sinapsi verrà attenzionata
successivamente dalla professoressa Metro). In realtà a livello sinaptico non sono liberati soltanto dei neurotrasmettitori, ma anche
altre sostanze che si chiamano neuromodulatori, sostanze che appunto modulano la neurotrasmissione. Di norma i
neurotrasmettitori sono delle amine biogene, quindi piccole molecole, invece i neuromodulatori di norma sono neuro peptidi; i
neurotrasmettitori liberati a livello sinaptico agiranno o su altre cellule nervose :
sinapsi interneuronali o sinapsi centrali perché si trovano a livello del SNC
sinapsi tra neuroni e degli effettori come per esempio il muscolo scheletrico (la sinapsi neuromuscolare è la placca
motrice)
Ormoni endocrini. Ci riferiamo alle classiche ghiandole endocrine, o cellule specializzate alla sintesi, all’immagazzinamento e alla
liberazione di ormoni. Ormoni che passeranno in circolo e agiranno a distanza.
-Nella secrezione endocrina classica abbiamo le cellule endocrine (che possono essere strutturate in ghiandole endocrine o cellule
endocrine presenti nel contesto di un tessuto (come per gli ormoni gastrointestinali, prodotti a livello gastrico o duodenale, ma non
da ghiandole endocrine, ma da cellule presenti a livello gastrico o intestinale: cellule D per la somatostatina; cellule G per la
gastrina etc.etc. la cellula endocrina produce un ormone che passa in circolo e attraverso la via ematica raggiungerà la cellula
bersaglio (azione a distanza). Ovviamente per poter agire a livello del bersaglio dovrà essere riconosciuto da un recettore.
-Nella secrezione paracrina si parla di
sostanze paracrine, ma in realtà sono
sempre ormoni. La differenza sta nel fatto
che la sostanza chimica, l’ormone, agisce
non a distanza, ma nelle vicinanze della
ghiandola che produce l’ormone senza far
ricorso a l circolo sanguigno, con la semplice
diffusione attraverso i liquidi interstiziali
l’ormone agirà su un’altra cellula che verrà o
stimolata o inibita. Per esempio la cellula D
che produce somatostatina attraverso la via
paracrina agirà sulla cellula G che produce
gastrina e la inibirà.
-Per quanto riguarda gli ormoni
neuroendocrini, sono prodotti da neuroni
che oltre a generare, propagare e
trasmettere impulsi, hanno anche la capacità
di produrre ormoni neuroendocrini. Quindi si
parla di ormoni che verranno sempre liberati
nel sangue e attraverso questo, veicolati
fino agli effettori (che se presentano i
recettori specifici potranno svolgere le loro
funzioni ). Nella secrezione neuroendocrina
il neurone produce l’ormone e questo per
via assonica verrà trasportato a livello del
terminale assonico: i bottoni terminali. Il contatto non avviene tra neurone ed altri neuroni, ma tra neurone ed un sistema di
capillari: l’ormone attraverso il circolo sanguigno giungerà all’effettore. Questa funzione potrà essere svolta da un’unica struttura
nervosa: l’ipotalamo.
-Nelle secrezioni autocrine l’ormone agisce sulla stessa cellula che l’ha prodotto. Di norma questa secrezione è di natura inibitoria
per evitare un’eccessiva secrezione ormonale (una del le caratteristiche dell’attività ormonale è che l’ormone agisce in piccolissime
quantità).
Distinguiamo due modalità di secrezione: di tipo fisiologico (quando
l’ormone agisce in piccole concentrazioni) o azione di tipo
farmacologico (quando l’ormone agisce in quantità più elevate)
Il glucagone è un ormone glicogenolitico (questa attività si attua
mediante livelli modestissimi di glucagone), ma se abbiamo degli
stimoli particolari e si ha un’eccessiva risposta della cellula che
produce glucagone si ha un’azione farmacologica e il glucagone ad
alto dosaggio è capace di provocare un effetto inotropo positivo con
l’aumento della forza di contrazione del miocardio.
-Le citochine sono prodotte dalle cellule del sistema immunitario.
Alcune citochine sono prodotte dal tessuto adiposo bianco. Il tessuto
adiposo ha il compito di accumulare trigliceridi, ma questo è anche
capace di produrre una serie di sostanze , basti pensare che una
buona quota degli estrogeni prodotti dalla donna (anche l’uomo
produce estrogeni) è sintetizzata nel tessuto adiposo. Ne è
testimonianza l’amenorrea nelle atlete in cui si ha avuto un’estrema
perdita del tessuto adiposo. Anche altri prodotti come la leptina, che
è una citochina ed altre sostanze che si chiamano adipochine, hanno
effetti metabolici notevoli, sia positivi che negativi per l’organismo.
La leptina è un fattore (sostanza ormone-simile) capace di indurre sazietà.
Parlando della secrezione ormonale classica; ecco quali sono i principali distretti:
ipofisi: lobo anteriore e lobo posteriore (negli anfibi c’è anche un lobo intermedio; nell’uomo non è possibile separarlo e l’ormone
prodotto nel lobo intermedio negli anfibi viene prodotto nei mammiferi nell’adenoipofisi. Tale ormone è l’ormone melanotropo
stimolante (MSH) che permette la pigmentazione cutanea e nell’uomo è prodotto dal lobo anteriore non dal lobo intermedio).
Tiroide
paratiroidi
timo(la timulina è da stimolo per i linfociti T)
Pancreas endocrino(isole di Langerhans)
surreni(corticale e midollare)
gonadi
Queste sono ghiandole, ma non è che sono costituite solo ed esclusivamente da cellule endocrine, ma sono degli organi veri e
propri in cui la componente morfologica principale sono le cellule endocrine; sono strutture altamente vascolarizzate; presentano
cellule di sostegno e così via.
Non possiamo parlare di ipotalamo come ghiandola endocrina; nell’ipotalamo ci sono neuroni capaci di produrre ormoni e ne
distinguiamo due tipologie: i magnocellulari e i parvicellulari.
I magnocellulari sono situati soprattutto nel nucleo sopraottico e paraventricolare e producono 2 ormoni: ossitocina, vasopressina.
I parvicellulari sono situati soprattutto nel nucleo arcuato e nel nucleo paraventricolare e producono fattori capaci di influenzare
,stimolando o inibendo, l’attività dell’adenoipofisi. Quindi non è una ghiandola endocrina; l’ipotalamo presenta neuroni (il lobo
arcuato è molto importante anche nei meccanismi di fame sazietà)che hanno diversa funzione oltre quella di produrre fattori
ipotalamici capaci di agire sull’ipofisi: le liberine o i releasing factors o possono inibire la produzione di ormoni ipofisari.
Poi abbiamo altri ormoni diffusi, dove le cellule sono
presenti non in ghiandole, ma sparse nei tessuti tra cui:
lo stomaco: gastrina etc.
l’intestino tenue: colecistochinina e secretina (la
pancreozimina è identificabile con la colecistochinina poiché
questa agisce sia sul pancreas che sulla colecisti)
rene: ormone D, eritropoietina (fattore ormone simile), la
renina non è un ormone, è un enzima
la placenta: ha funzione trofica, ma anche endocrina in
quanto produce progesterone.
Il trofoblasto negli stati iniziali produce anche la
gonadotropina corionica che agisce sul corpo luteo gravidico
permettendo la produzione di progesterone, finché non si
sarà formata la placenta. I test di gravidanza consistono
nella ricerca della gonadotropina corionica.
Cute: produce precursori della vitamina D -> ormone D a
livello renale
Qual è il ruolo svolto dagli ormoni?
-Processi di riproduzione: ovaio e testicolo sono gli organi direttamente interessati
-Crescita e sviluppo: bersaglio sono le ossa, i muscoli e i visceri. L’ormone per eccellenza è il GH, ma non solo; una serie di ormoni
sono deputati all’accrescimento e allo sviluppo corporeo come il testosterone e gli ormoni tiroidei. In realtà abbiamo diversi tipi di
ormoni capaci di influenzare la crescita e lo sviluppo
-Mantenimento dell’omeostasi: pH, temperatura, glicemia, volume dei liquidi, osmolarità dei liquidi.
-Produzione, utilizzazione e deposito di energia, in questo caso i bersagli sono i mitocondri oppure il tessuto adiposo (sia quello
bruno che quello bianco). Il bianco ha la funzione di deposito di trigliceridi, il grasso bruno ha invece funzione termogenetica.
Poi, possiamo classificare gli ormoni tenendo conto della natura chimica:
-natura aminoacidica: di norma l’aminoacido è la tirosina, che è precursore di adrenalina, noradrenalina e delle catecolamine; delle
iodotironine: T3 (triiodotironina) e T4 (tiroxina)
-natura proteica: la maggior parte degli ormoni e possono essere proteine semplici o proteine coniugate, come glicoproteine (ad
esempio la gonadotropina corionica, il FSH, l’LH, l’ormone tireotropo , l’insulina)
-natura steroidea: derivano dalla struttura del colesterolo. Sono i glucocorticoidi (come il cortisolo), i mineralcorticoidi (come
l’aldosterone e il corticosterone), il DHEA (deidroepiandrosterone, prodotto a livello della ghiandola surrenale) e gli ormoni sessuali
(prodotti dalle ghiandole ovaio e testicolo: soprattutto estrogeni, testosterone e progesterone)
Un’altra classificazione tiene conto dei rapporti con l’ipofisi, esistono due tipologie:
-ormoni prodotti da ghiandole endocrine che dipendono dall’ipofisi, che a sua volta dipende dall’ipotalamo. Queste ghiandole sono
la tiroide, il surrene, le gonadi. Gli ormoni prodotti sono le iodotironine, il cortisolo gli ormoni sessuali (che poi agiranno sulla cellula
bersaglio dandoci gli effetti propri ormonali. Sono quindi ormoni che agiscono sulla secrezione di altri ormoni. Dobbiamo ricordare
che a livello dell’adenoipofisi vengono prodotti altri due ormoni che non hanno come bersaglio un’altra ghiandola endocrina e sono
la prolattina e l’ormone GH (ormone della crescita).
-ormoni prodotti da ghiandole endocrine non dipendenti dall’adenoipofisi. Sono ormoni che di norma non controllano livelli ematici
di ormoni, ma parametri chimici, ad esempio il paratormone e calcitonina (influenzano l’omeostasi calcica), insulina e glucagone
(hanno come effetto il mantenimento di livelli costanti di glicemia).
Vi ho parlato di APUD ormoni, le cellule APUD (Amine Precursor Uptake and Decarboxylation) contengono amine come
catecolamine e serotonina. Sono capaci di prelevare dal sangue i precursori e hanno la capacità di decarbossilare gli aminoacidi e
producono di norma ormoni di natura amminica, di natura peptidica o di natura glicoproteica. Ecco infatti gli APUD ormoni sono
l’ossitocina, gli ormoni adenoipofisari, la melatonina (prodotta dall’epifisi, riveste importanza nei ritmi circadiani), insulina,
glucagone etc., poi abbiamo le iodotironine che sono il T3 e il T4
Gli ormoni steroidei derivano da un comune precursore: il colesterolo o di origine endogena (la cellula possiede una riserva o può
produrlo a partire da acetato) o di origine esogena (proviene dal sangue attraverso le LDL), verrà trasformato in pregnenolone e
successivamente in progesterone, precursore della maggioranza degli ormoni steroidei.
Ricordate che la vasopressina e ossitocina (ipofisi posteriore), non sono di origine neuro ipofisaria, ma sono di origine ipotalamica.
Caratteristiche dei diversi tipi di ormoni, da cui dipende il meccanismo d’azione.
Solubilità
Sito recettoriale
Sintesi
Peptidi
Catecolamine
( sia proteine semplici
che glicoproteine)
Idrofili : Gli ormoni non attraversano la membrana
Ormoni tiroidei
cellulare
membrana plasmatica
Recettori con localizzazione
nucleare
Livello extracellulare: nella
cavità follicolare è contenuta
la colloide ( contenente
tireoglobuline)
Colloide
Recettori di membrana
RER -> apparato del Golgi
Classica sintesi proteica
citoplasma
Granuli secretori
Immagazzinamento
Esocitosi
Secrezione
Trasporto
Liberati come ormoni
liberi
Ghiandole
interessate alla
sintesi
La maggiore quota
In parte libere in parte
legate alle proteine
plasmatiche
Midollare del surrene
Steroidi
Lipofili: attraversano facilmente la matrice lipidica della
Recettori localizzazione
citoplasmatica
Citoplasma
REL
Mitocondri
Esistono come tali nel
citoplasma: non ci serve
una riserva. Le quantità
pertanto sono modeste.
La sintesi può essere fatta
ex novo. Non occorrono
riserve
Endocitosi dalla colloide
Diffusione semplice
essendo liposolubili
Nel 99% dei casi legati a proteine plasmatiche.
Soprattutto prealbumine e albumine.
Cellule follicolari della tiroide
Non le parafollicolari (o
cellule C) che sintetizzano
tireocalcitonina
Corticale del surrene
Gonadi
Meccanismo di secrezione. Come vengono prodotti gli ormoni?
Gli ormoni peptidici vengono prodotti tutti attraverso un precursore inattivo: il preproormone. Per ogni ormone abbiamo uno o più
geni specifici che codificano per l’ormone. Il DNA permetterà la trascrizione dell’RNA messaggero. La sintesi avviene a livello dei
ribosomi. Mentre i ribosomi presenti a livello del citoplasma
sintetizzano proteine non accumulabili in granuli di secrezione, i
ribosomi del RER sintetizzano proteine accumulate dentro
vescicole: i granuli di secrezione. Un preproormone, in cui è
presente: una sequenza segnale, l’ormone e un fattore (il
copeptide). Il segnale serve a trasferire il preproormone al
reticolo endoplasmatico, nella cavità del reticolo. Quindi nel
reticolo endoplasmatico abbiamo la maturazione a proormone,
che può subire delle modifiche e si accumulerà in granuli grazie
all’attività dell’apparato di Golgi : quindi si formano granuli
rivestiti da vescicole.
Quindi nella biosintesi del preproormone la molecola segnale
permette l’ingresso nel RE, poi la sequenza segnale si stacca: si
forma il proormone. In realtà vengono sintetizzati non solo gli
ormoni, ma anche gli enzimi di clivaggio che trasformeranno il
proormone in ormone. Nei granuli io trovo ancora il pro
ormone ed è nei granuli che avvengono le trasformazioni che
porteranno dal pro ormone all’ ormone che verrà liberato
tramite esocitosi. Ad esempio nella sintesi dell’insulina: le due
catene sono legate con ponti disolfuro, una volta si pensava che
la molecola fosse sintetizzata come due catene separate che un
enzima capace di legare dei ponti disolfuro, avrebbe unito, tale enzima era definito zippasi, perché come in una zip univa le due
catene. Ora invece sappiamo che i ponti disolfuro esistono già quando viene sintetizzata la preproforma, ma la molecola
inizialmente ha un andamento lineare: la pre pro insulina, conterrà il segnale leader, conterrà l’insulina con le due catene e il
peptide C che unisce le due catene. Ciò avviene nel RER, cioè nel ribosoma e la sequenza segnale trasferisce la preproinsulina nel
RER, dove la forma lineare verrà ripiegata a formare due catene legate con il peptide C. devono esserci almeno 2 endopeptidasi in
grado si allontanare il peptide C.
Consideriamo ora la calcitonina, un ormone prodotto nelle
cellule parafollicolari della tiroide, chiamato anche
tireocalcitonina. Vedremo nella via del dolore che esiste un
neuro modulatore che viene chiamato CGRP ed è un peptide
correlato col gene della calcitonina (peptide correlato al
gene della calcitonina), cioè avremo un unico gene, ma
questo stesso gene a livello della cellula C della tiroide
permetterà di trascrivere la calcitonina; invece a livello del
sistema nervoso, lo stesso gene codificherà una calcitonina
simile che però non ha gli effetti ipocalcemizzanti
dell’ormone prodotto nella ghiandola tiroidea, ma questo
neuro peptide fungerà da neuro modulatore. Quindi
esisteranno due forme diverse di molecole. Ci sono delle
molecole come la vasopressina (che conosciamo come un
ormone)che in certi circuiti si comportano da
neuromodulatori. Quando farete la sinapsi; il neuro
trasmettitore verrà accumulato in granuli, in vescicole e
liberato per esocitosi, vedrete che che a livello sinaptico
l’esocitosi è un fenomeno molto complesso.
Come avviene la secrezione? Per via della endo-esocitosi.
L’esocitosi cosa presuppone? Che la membrana della
vescicola che in questo caso contiene l’ormone si fonda con la membrana plasmatica della cellula della ghiandola che dovrà liberare
tale ormone. I granuli secretori sono presenti nel citoplasma e quindi si devono spostare in direzione della membrana plasmatica, in
modo che possa avvenire la fusione, quindi abbiamo bisogno di uno spostamento di granuli. Esiste un sistema di microtubuli e di
microfilamenti, che hanno quindi un’attività contrattile che permetterà lo spostamento dei granuli in modo da consentire la
fusione. Essendo microtubuli e microfilamenti strutture contrattili vediamo interessato lo ione calcio ca++. Le cellule non
dispongono di riserve di calcio, poiché è tossico per la cellula. Quindi il calcio proviene dall’esterno e perché si possa verificare
l’esocitosi e quindi prima lo spostamento e la fusione dei microgranuli è necessario che il calcio si leghi ad una proteina: la
calmodulina (che legando calcio diventa attiva); quindi una serie di secondi messaggeri che agirà per mezzo di proteinchinasi,
tramite processi di
fosforilazione e di
defosforilazione che alla fine
porteranno allo
spostamento dei granuli, alla
fusione e quindi alla
secrezione. Sono dunque
necessarie calmodulina e
ioni calcio.
Per quanto riguarda gli
ormoni steroidei: la sintesi
ha origine da un precursore
comune che è il colesterolo;
questo perde la catena
laterale per mezzo di una
desmolasi. La via biosintetica
è illustrata.
Se ci sono dei polimorfismi
che portano ad una mancata
sintesi a certi livelli, si
formano
pseudoermafroditismi,cioè si
hanno conseguenze sullo
sviluppo dell’embrione.
La via metabolica sicuramente verrà dimenticata, ma è necessario non dimenticare i danni che porteranno ad una anomalia
dell’attività metabolica con delle ripercussioni, soprattutto per quanto riguarda gli ormoni sessuali ovviamente sulla maturazione e
così via degli organi sessuali. Il primo effetto è il distacco della catena laterale (e possiamo definire per mezzo di un enzima
limitante che è la desmolasi NADPH dipendente), poi abbiamo gli altri processi che interessano la steroido-genesi.
Per quanto riguarda la sintesi delle catecolamine: tirosina, diossifenilalanina (DOPA), dopamina, tutto a livello citoplasmatico, il
passaggio della dopamina a livello del granulo secretorio e qui verrà trasformata in norepinefrina; se vogliamo sintetizzare
adrenalina, la noradrenalina passa di nuovo nel citoplasma e un enzima metilante formerà adrenalina.
Nelle sinapsi noradrenergiche il neurotrasmettitore per eccellenza è la noradrenalina. L’adrenalina è invece l’ormone della
midollare del surrene, però vi sono circuiti nel Sistema nervoso centrale adrenergici, quindi sempre adrenalina e non
noradrenalina. Ecco come dicevamo prima perché non abbiamo bisogno di immagazzinare gli ormoni steroidei in granuli, perché li
posso sintetizzare a partire dal colesterolo e li posso depositare come esteri del colesterolo in vacuoli, o possono derivare dalle HDL
o dalle LDL. Abbiamo bisogno di 2 ormoni l’ACTH nella corticale del surrene e l’LH per gli ormoni delle gonadi. La sintesi avviene
inizialmente a livello del mitocondrio: il distacco avviene a livello periferico citoplasmatico, poi abbiamo il colesterolo che forma il
pregnenolone, passa nel REL formando nelle gonadi gli ormoni sessuali.
Per quanto riguarda gli ormoni steroidei e gli ormoni tiroidei, per la tiroxina e la triiodotironina la percentuale legata alle proteine
plasmatiche (prealbumine o albumine) è elevata, circa il 99%; anche per gli altri vale lo stesso: cortisolo 94% legato ad albumine,
testosterone 89% etc. essendo lipofili la proteina vettrice è l’albumina o la prealbumina o altre famiglie di globuline. Invece la
percentuale degli ormoni proteici legata a proteine è insignificante.
Questo schema molto semplice descrive i tipi di recettori. Di norma un recettore è costituito da 2 subunità: una subunità recettrice
ed una subunità effettrice.
Gli ormoni steroidei e gli ormoni tiroidei attraversano la membrana citoplasmatica e si legano a dei recettori citoplasmatici per
quanto riguarda gli ormoni steroidei e poi il complesso
recettore-ormone si trasferisce all’interno del nucleo, si legherà
al DNA e quindi avverrà la trascrizione genica, sintesi delle
proteine responsabili dell’azione dell’ormone. Quindi per gli
ormoni steroidei il sito recettoriale è a livello citoplasmatico.
Diverso è per gli ormoni tiroidei; il recettore sarà nucleare, si
legheranno a questo e quindi avremo gli effetti tipici degli
ormoni tiroidei, che consistono sempre nella trascrizione di
mRNA e la sintesi di proteine con diversa natura funzionale. Per
gli ormoni idrofili le cose sono un po’ diverse: non attraversano
la membrana lipidica quindi troveranno gli enzimi a livello della
membrana plasmatica. Per esempio l’insulina riconosce un
recettore di membrana costituito da una sub unità recettrice e
da una effettrice; una volta che si ha avuto il legame
dell’ormone col recettore si potranno avere gli effetti
intracellulari specifici. I recettori per l’insulina, ma anche altri vengono chiamati recettori con attività enzimatica, invece per
esempio le catecolamine e il glucagone agiscono attraverso un secondo messaggero e quindi l’effetto è mediato dalla proteina G.
Un recettore per le catecolamine è costituito da una
subunità recettrice, da una subunità d’accoppiamento
che sarebbe la proteina G, e la subunità effettrice che
sarebbe il secondo messaggero a cui saranno dovuti
effetti metabolici o delle catecolamine o del glucagone.
Analizziamo i recettori di membrana:
abbiamo recettori con attività enzimatica, come i
recettori per l’insulina e i fattori di crescita come quello
prodotto dal GH a livello epatico ( IGF1 chiamata tempo
fa somatomedina), questi recettori vengono chiamati
recettori tirosinachinasici perché hanno la capacità di
catalizzare la fosforilazione di residui di tirosina in
proteine che contengono questo aminoacido.
I recettori serina-treonina-chinasi sono sempre recettori
con attività enzimatica e sono tipici dei recettori per i fattori
di degradazione e per i fattori di inibizione
I recettori guanilatociclasici portano alla sintesi di cGMP
(GMP ciclico) attraverso l’intervento della proteina G,
recettori per esempio del fattore natriuretico atriale. Tra gli
organi endocrini non dobbiamo dimenticare il miocita
atriale. Questi contengono granuli che producono una
miscela di fattori natriuretici atriali che inibiscono il
riassorbimento di sodio a livello tubulare. Stiamo sempre
parlando di recettori con attività enzimatica.
Salto all’ultimo, perché anche questo recettore delle
citochine sono recettori ad attività enzimatica, esempio di
recettore per le citochine, recettori per l’ormone della
crescita, della prolattina e della leptina.
Poi abbiamo recettori accoppiati alle proteine G: sono
recettori per le catecolamine, recettori per gli ormoni
glicoproteici, per il glucagone, per il paratormone, anche
per i colinergici muscarinici (l’acetilcolina reagisce con i recettori muscarinici).
Per quanto riguarda il recettore per l’insulina, è una proteina trans
membrana e attraversa tutta la membrana. Abbiamo 3 domini
funzionali:
-un dominio extracellulare
-un dominio transmembrana
-un dominio intracellulare
Sono legati da ponti disolfuro. La componente extracellulare ha la
capacità di legare o l’insulina, o il fattore di crescita. Una volta che si
ha avuto questo legame verrà attivata la componente citoplasmatica
che ha una attività enzimatica di tipo tirosina-chinasica, capace di
auto fosforilare il dominio intracellulare; se il legame col recettore
non avviene, non c’è il processo di autofosforilazione. Questo
processo è fondamentale, perché se io fosforilo il dominio
citoplasmatico intracellulare, questa componente del recettore entra
in attività e assume anch’esso un’attività tirosina-chinasica, capace di
fosforilare substrati citoplasmatici anche a livello nucleare. Ecco per
esempio, una volta che si ha avuto l’autofosforilazione della
componente del dominio citoplasmatico, esso verrà trasformato in
tirosina-chinasi che potrà fosforilare substrati citoplasmatici o meno,
a cui poi permetteranno le funzioni tipiche, come la traslocazione di
trasportatori, dal citoplasma alla membrana, trasportatori non
soltanto di glucosio, ma anche di aminoacidi. L’insulina non è un
ormone che limita i suoi effetti solamente al metabolismo glicidico, è
panmetabolico: aumenta anche la sintesi proteica,favorendo anche
l’ingresso di aminoacidi dall’esterno, oppure verranno attivati enzimi
già pronti che se fosforilati verranno inattivati , se defosforilati
passeranno alla forma attiva. Quindi enzimi già presenti all’interno
della cellula. Ma vedete che questo fattore di crescita è capace di
agire anche a livello nucleare, infatti verranno attivati dei fattori di trascrizione capaci di modulare l’espressione genica
permettendo la sintesi ex-novo di nuove proteine. In questo caso io attivo proteine a funzione enzimatica già presenti, che devono
essere fosforilati per essere attivi o che invece ha bisogno di sintesi ex novo. L’insulina si lega all’unità recettoriale. Questa è l’unità
catalitica che viene fosforilata ed avrà attività tirosina chinasica che avrà come bersaglio i substrati dei recettori insulinico che di
norma sono proteine che potranno fosforilare enzimi quali quelli per la sintesi proteica, biosintesi degli acidi grassi, sintesi di
glicogeno e crescita ed espressione genica e trasporto della vescicola che contiene glucosio. In realtà i substrati del recettore
insulinico sono moltissimi ad esempio se io attivo questa molecola che si chiama AKT, avviene la traslocazione della vescicola che
contiene i recettori per i canali del glucosio: i GLUT, a livello di membrana plasmatica . La MAP chinasi, fattore di trascrizione
responsabile dell’espressione genica, è un fattore di trascrizione che consente la trascrizione dell’ mRNA. Invece questa che stiamo
per descrivere è molto simile alla precedente: è un recettore per la prolattina, per la leptina e per il GH : abbiamo un dominio
citoplasmatico (intracellulare), uno transmembrana e uno extracellulare. La differenza sta nel fatto che il dominio intracellulare non
ha attività enzimatica tirosina-chinasica, quindi non è capace di autofosforilarsi. Quando l’ormone si lega al sito recettoriale il
dominio intracellulare viene attivato: quindi è capace di agire. Questa attivazione determina l’ attrazione e ancoraggio alla frazione
citoplasmatica di un enzima con attività tirosina-chinasica: quindi l’attivazione dell’unità intracellulare in seguito al legame
dell’ormone, inizialmente permetterà l’attrazione e l’ancoraggio di una tirosina-chinasi citoplasmatica: di conseguenza questa
tirosina-chinasi verrà attivata e quindi verranno fosforilati dei
fattori di trascrizione che permetteranno la modulazione
dell’espressione genica. Nel caso dell’insulina il fattore di
trascrizione veniva chiamato MAP chinasi, qui invece le
tirosina chinasi attivate prendono il nome di Jak 2 (chinasi
associate a Janus), questa una volta fosforilata attiverà enzimi
dandoci effetti fisiologici; oppure verrà attivato un fattore di
trascrizione che si chiama CREB, trasferito a livello del nucleo
e avrà come bersaglio un gene specifico e quindi verrà indotta
la sintesi di un messaggero. Quindi avremo come per
l’insulina, effetti citoplasmatici immediati o effetti nucleari
tardivi, perché ho bisogno che la cellula venga attivata,
trasferita a livello nucleare e lì svolgere le sue funzioni; quindi
avremo effetti a breve termine se attivo enzimi già presenti o
effetti a lungo termine se io produco un nuovo enzima in
questo caso legata a questo fattore di crescita. Per quanto
riguarda il recettore correlato con l’ormone per esempio
natriuretico , recettore cosiddetto guanilatociclasi, anche
questo come i precedenti ha un dominio intracellulare, una
parte trans membrana e una parte extracellulare. Cambia l’attività che è guanilatociclasica e porterà alla sintesi di cGMP e avremo
l’effetto ; un altro effettore è l’ossido di azoto, che è un vasodilatatore che può essere attivato in una struttura periferica.
Descriviamo ora la famiglia dei recettori accoppiati alle proteine G e qui dobbiamo discutere di effetti a breve termine e effetti a
lungo termine, che presuppone sempre la sintesi ex-novo di proteine, l’intervento del nucleo, dei fattori di trascrizione …
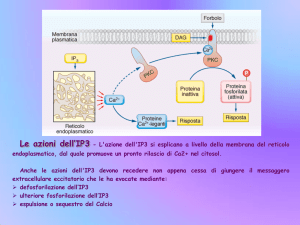
L’ormone si lega al sito recettoriale accoppiato alla proteina G, questa viene attivata, attiva i due secondi messaggeri, che sono a
livello ormonale e sono correlati: l’AMP ciclico (cAMP), l’inositolo trifosfato (IP3) e il diacilglicerolo (DAG).
Quindi è la proteina g che attiva l’adenilatociclasi, portando alla formazione dell’ cAMP. L’ cAMP attiva una chinasi specifica, la
chinasi A, cAMP dipendente, questa attivata fosforilerà delle proteine enzimatiche, di membrana e così via per avere l’effetto tipico
dell’ormone. Oltre ad attivare enzimi già preformati, verrà anche attivato un fattore di trascrizione che dipende dall’ cAMP ed è il
CREB, raggiungerà il nucleo, attiverà il gene, avvierà la trascrizione del messaggero, quindi la sintesi proteica.
L’NGF è un fattore di crescita che troviamo nel sistema nervoso, scoperto da Rita Levi Montalcini, porterà alla sintesi, all’attivazione
di una proteina RAS, che attiva una chinasi che a sua volta attiverà una MAP. La MAP chinasi permetterà la sintesi di nuove proteine
oppure attiverà una fosfolipasi C , quindi avremo IP3 e diacilglicerolo (siamo partiti da un fosfolipide di membrana), a livello
nervoso, la sintesi di questi fattori porterà alla crescita dell’assone e al differenziamento cellulare. Invece se viene attivata la
fosfatocreatinasi, verrà attivata l’HT chinasi che permetteva la traslocazione di un canale nell’insulina, in questo caso necessaria alla
sopravvivenza della cellula. Oppure il fattore di crescita con
un fattore tirosina-chinasico attiva la MAP chinasi che
riconoscerà un sito nel gene. Se invece si parla dell’ cAMP
nella trasduzione legata ad una proteina G la proteina-chinasi
attiverà questo fattore di trascrizione permettendo la
trascrizione genica.
Se abbiamo recettore, proteina G, la proteina effettrice
l’enzima, il secondo messaggero che è l’ cAMP , questo
porterà alla fosforilazione di proteine che interesseranno gli
effetti ormonali.
Alla fine una volta fosforilata una proteina avremo la risposta
cellulare che consisterà in spostamento di materia ( se agisce
su proteine motrici), attività enzimatica, traslocazione,
attivazione gene e sintesi proteica come effetti a lungo
termine.
Facciamo un esempio: prendiamo una catecolamina: la
epinefrina, o anche la noradrenalina. Sappiamo che esistono
due classi di recettori diversi: gli α-recettori e i β-recettori.
Questi ultimi si dividono in β1 e in β2. Esistono anche α1 e
α2, ma gli α2 sono auto recettori. Quando l’adrenalina agisce
su un effettore β, verrà attivata una proteina G, che stimolerà
l’enzima effettore con la sintesi di cAMP. Quindi la proteina G
è correlata con l’cAMP. Se invece l’ormone si lega con un
recettore α1 (postsinaptico), allora il legame della
noradrenalina con il recettore avrà effetti diversi: verrà
attivata una proteina G, che attiva però la fosfolipasi C, che ci permetterà di ricavare DAG e IP3. Il IP3 permetterà al calcio di essere
liberato dalle riserve di calcio intracellulare, invece il DAG attiva una proteinasi C. Lo stesso ormone potrà avere effetti diversi,
perché è diverso il recettore.
L’ormone tiroideo, invece, attraversa la matrice lipidica, poiché è in parte idrosolubile non ha problemi con l’acqua
citoplasmatica,quindi raggiungerà il nucleo e il recettore per l’ormone tiroideo; una volta che l’ormone tiroideo abbia legato il
recettore nucleare , verrà attivata l’espressione genica etc.etc.
L’ ormone steroideo invece è liposolubile, quindi attraversa la matrice lipidica della membrana e deve essere veicolato da un
recettore citoplasmatico, che lega l’ormone steroideo; il complesso ormone recettore verrà veicolato all’ambiente nucleare dove
avverranno i meccanismi accennati. Il recettore presenta un sito di legame con l’ormone e un rito di legame con il DNA.
La quantità dell’ormone che viene prodotto è modestissima.
Abbiamo due famiglie di ormoni, in base alla quantità:
-ormoni presenti in quantità stabile, come gli ormoni della tiroide
-ormoni con andamento oscillante, come catecolamine, ormone
antidiuretico.
L’entità, la quantità dipende dall’equilibrio tra sintesi e catabolismo.
È necessario però che i livelli di ormone si mantengano sempre a
livelli ottimali; è meglio evitare surplus o difetti. Abbiamo quindi
meccanismi di controllo delle quantità di ormoni circolanti. Tra i
meccanismi di controllo per eccellenza figurano i meccanismi a
feedback, cioè l’effetto agisce sulla causa.
Distinguiamo feedback positivi e feedback negativi.
Esempio di feedback positivo: un aumento della glicemia mi
comporterà un aumento dell’insulina; una riduzione della
ipoglicemia mi porterà ad un aumento del glucagone, una
diminuzione della calcemia, mi porterà ad un aumento del
paratormone. Quindi abbiamo dei feedback positivi, ma la maggior parte sono negativi, cioè l’ormone verrà inibito nella sua
produzione per evitare una eccessiva secrezione. Intanto tutto dipende dall’ipotalamo. L’ipotalamo produce o fattori liberanti o
fattori inibenti l’azione della ghiandola ipofisi. L’ipofisi produrrà ormoni che agiranno sulla cellula bersaglio, può essere una tropina
(ACTH, TSH), che agirà su una seconda ghiandola, o meglio la terza, poiché la prima è l’ipotalamo, la seconda l’ipofisi, la terza è la
ghiandola che riceve il segnale che potrebbe essere il surrene, la tiroide o le gonadi. Quindi feedback significa che l’effetto potrà
agire sulla causa che l’ha determinato, di norma in modo inverso. Immaginiamo che ci sia stato uno stimolo che mi abbia
aumentato la secrezione di fattori di rilascio, nell’ipofisi e quindi alla ghiandola bersaglio. La ghiandola bersaglio produrrà l’ormone
e per evitare un eccesso di produzione da parte dell’adenoipofisi, abbiamo un feedback negativo. L’ormone prodotto anche da una
terza ghiandola potrà agire o sull’ipofisi, quindi un feedback diretto, inibendo, o meglio riducendo la secrezione dell’ormone;
oppure potrà agire sull’ipotalamo , quindi la ghiandola bersaglio produrrà l’ormone, che oltre ad agire a livello ipofisario inibendola,
potrà agire anche a livello di ipotalamo così verrà inibito il fattore di rilascio di quell’ormone. Questo è un feedback indiretto perché
non agisce direttamente sull’ipofisi, ma tramite l’ipotalamo. Questi feedback che dall’organo bersaglio si portano all’ipofisi, o
dall’organo bersaglio di portano all’ipotalamo uno diretto e l’altro indiretto sono feedback lunghi perché l’ormone prodotto dalla
ghiandola bersaglio dovrà passare in circolo e raggiungere o l’ipofisi o l’ipotalamo. Quindi feedback lunghi: il primo diretto, il
secondo indiretto. Poi abbiamo dei feedback ad ansa corta in questo caso è l’ormone prodotto dall’adenoipofisi che agirà
sull’ipotalamo. Quindi avremo un feedback negativo tra ipofisi e ipotalamo ad ansa corta. Poi abbiamo un meccanismo di controllo
paracrino e quindi ad esempio l’ormone prodotto dall’adenoipofisi potrà autolimitarsi, quindi feedback ad ansa ultracorta dove la
tropina agisce sulla cellula stessa che sintetizza la tropina. Un ansa
ultracorta potrà essere nel neurone ipotalamico che produce un
determinato ormone, agirà mediante un meccanismo autocrino sulla
cellula stessa che ha prodotto il fattore di rilascio.
Per capire questo processo è meglio riassumere : i neuroni ipotalamici
secernono il primo ormone che raggiungerà l’adenoipofisi attraverso il
circolo portale ipotalamo-ipofisario; l’adenoipofisi produrrà il secondo
ormone che passerà in circolo e raggiungerà la ghiandola surrenale,
dove verrà prodotto il terzo ormone, che potrà inibire o direttamente o
indirettamente attraverso un’ansa lunga o del fattore adenoipofisario
(l’ormone 3 agisce sull’ormone 2)oppure sull’ipotalamo (l’ormone 3
agisce sull’1); il corto è espresso tra ipofisi anteriore e ipotalamo;
l’ultracorto agirebbe direttamente sulla ghiandola che lo ha sintetizzato.
Se prendiamo in esame l’ormone e in particolare l’ACTH, l’ipotalamo se
opportunamente stimolato rilascerà il fattore di rilascio per la
corticotropina, cioè l’ACTH che raggiungerà l’ipofisi anteriore attraverso
il piccolo circolo portale; l’ipofisi risponderà con la produzione di ACTH
che passerà in circolo giungendo a livello della corticale del surrene,
dove viene sintetizzato per esempio cortisolo; il cortisolo potrà inibire o
l’adenoipofisi a produrre ACTH o l’ipotalamo a produrre il fattore
rilasciante. Questo è un feedback lungo: il primo diretto, il secondo
indiretto. La stessa ACTH potrà agire sull’ipotalamo e inibire il
corrispondente fattore di rilascio ( feedback corto); ma lo stesso
ipotalamo potrà gestire la propria attività attraverso un feedback ultracorto. I meccanismi a feedback sono bivalenti. L’attività
dell’ipotalamo potrà essere influenzata da altre componenti nervose. Per esempio sonno e veglia, possono modificare la risposta
ormonale, il dolore, le emozioni, la paura, la rabbia, gli odori, la visione, la vista,possono modificare la secrezione ormonale. Per
esempio gli odori negli animali inferiori possono modificare la secrezione di ormoni sessuali. Queste influenze deriveranno dunque
dal sistema visivo per la vista,la visione ; dal sistema limbico per le emozioni, la paura, la libido; il dolore dal talamo ; sonno e veglia
da una componente del tronco dell’encefalo che si chiama formazione reticolare tronco-encefalica. Queste componenti (anche la
corteccia) possono agire sulla secrezione ormonale agendo o direttamente sulla ghiandola bersaglio, o a livello dell’ipotalamo.
L’ipotalamo ha diverse funzioni: controlla i comportamenti, umore, affettività, istinto, libido, aggressività ect.
Funzioni fisiologiche, come sonno-veglia, controllo del sistema nervoso autonomo simpatico e parasimpatico; ma l’ipotalamo
produce ormoni che agiranno sull’ipofisi. L’ipotalamo è collegato con la corteccia, il sistema limbico e formazioni reticolari. Stimoli
visivi, stimoli emotivi, sonno e veglia possono modificare l’attività ipotalamica attraverso dei neurotrasmettitori; avremo neuroni
che sono corticali, o del sistema limbico, o della formazione reticolare, che agiranno sull’ipotalamo mediante la liberazione di
specifici neurotrasmettitori: catecolamine, acetilcolina, il GABA e così via.