06/12/2012
Mitocondri
Biotecnologie_2012
http://fffaif.org.au/?p=5249
Teoria endosimbiontica
http://learn.genetics.utah.edu/content/begin/
cells/organelles/
1
06/12/2012
Conversione di energia (1)
*
*l’ambiente in assoluto più ossigenato, se sangue arterioso
http://humanphisiology.wikispaces.com/05‐Hematology
Mediante una serie di reazioni che si svolgono nel citosol, l’energia
derivata dalla parziale ossidazione delle moleole di carboidrati
ricche di energia è utilizzata per formare ATP, la moneta energetica
delle cellule.
Un modo molto più efficace di generazione di energia è comparso
molto precocemente nell’evoluzione. E’ basato sulle membrane, e permette alle cellule di acquisire energia da una gran varietà di sorgenti. Es:
Conversione di energia luminosa in energia chimica nella
fotosintesi
Respirazione aerobica che permette di utilizzare l’ossigeno per produrre grandi quantità di ATP a partire delle molecole di cibo. http://thejns.org/na101/home/literatum/publisher/jns/journals/content/jn
s/1971/jns.1971.35.issue‐
3/jns.1971.35.3.0263/production/images/small/jns_1971_35_3_0263.fig00
7.gif
Conversione di energia (2)
La membrana che viene utilizzata per produrre ATP nei procarioti è la membrana plasmatica.
Negli eucarioti, la membrana plasmatica è riservata per altri processi di trasporto. Vice‐versa, delle membrane specializzate all’interno di organelli in grado di convertire energia, sono utlizzate per produrre ATP:
Mitocondri, presenti nelle cellule di praticamente tutti gli organismi
eucarioti (funghi, animali, piante)
Plastidi, sopratutto cloroplasti, che si trovano nelle piante.
Nelle foto al microscopio elettronico, l’aspetto morfologico più
appariscente dei mitocondri e cloroplasti è la grande quantità di membrane interne che essi contengono.
Accoppiamento chemiosmotico (1)
Meccanismo commune usato dai mitocondri, cloroplasti e procarioti per ricavare energia a scopi energetici.
Il termine riflette un collegamento tra le reazioni per formare legami chimici che generano l’ATP (“chemi”) e i trasporti di membrana (“osmotico”).
Le membrane interne forniscono l’impalcatura per un eleborato insieme di processi di trasporto di elettroni che produce la maggior parte dell’ATP
delle cellule. Accoppiamento chemiosmotico (2)
Produzione di energia nei mitocondri
Il processo di accoppiamento ha luogo in due passi collegati, che si svolgono entrambi mediante complessi proteici inseriti in una membrana.
Passo 1: Elettroni ad elevata energia (derivati dall’ossidazione di molecole di cibo, dall’azione della radiazione solare o da altre sorgenti) sono trasferiti lungo una serie di trasportatori di elettroni inseriti nella membrana. Questi trasferimenti di elettroni rilasciano energia che è utilizzata per pompare protoni (H+) che è ubiquitaria nelle cellule) attraverso le membrane e quindi per generare un gradiente protonico elettrochimico. [Un gradiente ionico attraverso una membrana è una forma di energia immagazzinata, che può essere utilizzata per svolgere lavoro utilie quando gli ioni hanno la possibilità di rifluire attraverso la membrana lungo il loro gradiente elettrochimico.
Passo 2: I protoni H+ fluiscono in dietro lungo il loro gradiente elettrochimico lungo una macchina proteica chiamata ATP sintasi, che catalizza la sintesi energia‐dipendente dell’ATP a partire dall’ADP e fosfato inorganico (Pi). Questo enzima ubiquitario gioca il ruolo di una turbina, permettendo che il gradiente protonico porti alla produzione di ATP.
2
06/12/2012
Membrane mitocondriali
•
I mitocondri sono circondati da due membrane, la membrana mitocondriale esterna e la membrana mitocondriale interna.
La membrana mitocondriale esterna racchiude completamente il mitocondrio e rappresenta il confine con il citosol.
La membrana mitocondriale interna contiene due domini interconnessi:
Membrana delimitante interna: sotto la membrana mitocondriale esterna. Particolarmente ricca di proteine implicate nell’importo di proteine mitocondriali.
Creste mitocondriali: si estendono verso l’interno dell’organello.
La membrana delimitante interna e le creste sono collegate da sottili connessioni – giunzioni delle creste.
Adattato da Karp G.: Biologia Cellulare e Molecolare: Concetti ed Esperimenti, EdiSES, IV ed., 2010; Smith & Wood, Cell Biology,2nd ed., Stanley Thornes, 1999.
Porine
Membrana mitocondriale esterna
E’ formata per il circa 50% di lipidi (lipidi simili a qulli della maggior parte delle altre membrane delle cellule eucariotiche.
Contiene solo il circa 5% della massa totale di proteine dei mitocondri.
Le proteine più abbondanti sono le porine, proteine integrali che formano ampi canali (2‐3 nm) circondati da una palizzata di foglietti β (similmente alla membrana esterna che fa parte della parete cellulare dei batteri Gram‐).
Le porine sono strutture dinamiche in grado di chiudersi reversibilmente a seconda delle condizioni della cellula:
Quando i canali delle porine sono aperti, la membrana esterna è permeabile a molecole con peso molecolare fino a 1000 o meno, in particolare anche l’ATP, il NAD e il coenzima A, che giocano un ruolo chiave nel metabolismo energetico.
Contiene diversi enzimi quali la NADH /citocromo b5 ossido‐reduttasi, la monoamina ossidasi e enzimi coinvolti nell’ossidazione dell’adrenalina, nella degradazione del triptofano e l’allungamento degli acidi grassi.
Membrana mitocondriale interna (1)
Presenti sia nella membrana esterna dei batteri Gram‐ che nella membrana esterna dei mitocondri
http://www.tulane.edu/~biochem/faculty/facfigs
/porin_page.htm
Praticamente priva di colesterolo ma ricca di un lipide inusuale il difosfatidil‐glicerolo, detto anche cardiolipina.
La cardiolipina, che ha una bassa concentrazione nella membrana mitocondriale esterna, conta per circa il 20% del contenuto totale di lipidi della membrana interna.
La membrana interna contiene più di 100 diversi tipi di polipeptidi
Contiene un alto rapporto proteine/lipidi (> 3:1 in peso; circa il 20% del contenuto totale di proteine dei mitocondri)
http://www.ncbi.nlm.nih.gov/books/NBK9898/figure/A1977/
Cardiolipina (1)
La cardiolipina (difosfatidilglicerolo) è un lipide molto concentrato della membrana mitocondriale interna, ma anche nella membrana plasmatica dei batteri, ossia in membrane coinvolte nella trasduzione di energia.
Cardiolipina (2)
La cardiolipina è importante per ridurre la permeabilità ai protoni
dando così origine al gradiente elettrochimico di protoni fra lo spazio intermembranario e la matrice dei mitocondri.
La cardiolipina è un “doppio” fosfolipide inusuale, che contiene quattro catene di acidi grassi e che si trova principalmente nella membrana mitocondriale interna. http://themedicalbiochemistrypage.org/lipid-synthesis.php
http://www.ncbi.nlm.nih.gov/books/NBK9885/figure/A1647/?report=objectonly
3
06/12/2012
Cardiolipina (3)
Nel cuore, tessuto ricco di mitocondri, i fosfolipidi mitocondriali svolgono diversi ruoli nella regolazione dei vari processi mitocondriali che includono l’apoptosi, il trasporto degli elettroni e l’importazione verso i mitocondri di proteine e lipidi. (Hatch GM. Cell biology of cardiac mitochondrial phospholipids. Biochem Cell Biol. 82: 99‐112, 2004).
Le membrane mitocondriali dividono i mitocondri in due compartimenti ripieni di fluido:
Matrice:
Consistenza di gel, dovuta ad elevata concentrazione di proteine idrosolubili (< 500 mg/ml).
Contiene diversi enzimi, in particolare quelli che catalizzano i vari passi del ciclo degli acidi tricarbossilici (ciclo di Krebs)
Contiene ribosomi e parecchie molecole di DNA a doppio filamento, di solito circolari.
Il DNA mitocondriale codifica per un piccolo n°di proteine (circa 13 nell’uomo) coinvolte nel trasporto degli elettroni nelle creste mitocondriali, per due RNA ribosomiali e per una ventina di tRNA.
Membrana mitocondriale interna (2)
E’ la membrana specializzata per la trasduzione di energia ed è impermeabile a quasi tutti i piccoli ioni.
Vi si trovano tre tipi principali di complessi proteici:
Componenti della catena di trasporto di elettroni (catena respiratoria): complessi I, II, II e IV.
L’ATP sintasi
Proteine di trasporto specifiche che mediano il trasporto di metaboliti verso e al di fuori della matrice mitocondriale.
L’architettura e la grande fluidità della membrana mitocondriale interna facilitano le interazioni tra i componenti richiesta per la produzione di ATP e in particolare la diffusione laterale di piccole proteine coinvolte nel trasporto di elettroni della catena di trasporto degli elettroni.
Spazio intermembrane
Proteine coinvolte nella morte cellulare programmata (apoptosi).
MATRICE & MEMBRANA MITOCONDRIALE INTERNA
Reazioni che catalizzano l’ossidazione del piruvato e degli acidi grassi a CO2 e H2O.
Sintesi ad esse accoppiata di ATP a partire da ADP + Pi.
4
06/12/2012
Mitocondri
DNA MITOCONDRIALE
http://www.personal.psu.edu/tcr5031/blogs/portfolio/blog/
DNA mitocondriale (1)
Mappa del DNA mitocondriale umano (mtDNA) che illustra i loci dove possono avere luogo mutazioni che portano a citopatie. Le lettere singole sono le abbreviazioni convenzionali ad una lettera degli aminoacidi.
ND: NADH deidrogenasi
CO: citocromo ossidasi
12S e 16S: RNAs ribosomali
http://www.ncbi.nlm.nih.gov/books/NBK2
1778/figure/A3442/?report=objectonly
Fissione e fusione dei mitocondri
DNA mitocondriale (2)
La catena respiratoria mitocondriale. Le subunità codificate dal DNA‐
nucleare sono in rosso, mentre le subunità codificate dal mtDNA sono in porpora. I protoni (H+) sono pompati dalla matrice verso lo spazio intermembranoso attraverso i complessi I, III e IV. Essi ritornano verso la matrice attraverso il complesso V (ATP sintasi) con la concomitante produzione di ATP. Il citocromo c (Cyt c) è una proteina di trasferimento di elettroni codificata dal DNA nucleare. ND: NADH deidrogenasi
SDH: succinato deidrogenasi
COX: citocromo ossidasi
A6 e A8: subunità del complesso V dell’ATP sintasi
http://www.ncbi.nlm.nih.gov/books/N
BK22005/figure/A3436/
Tre gruppi di reazioni
in regioni distinte dei mitocondri (1)
MATRICE (insieme a proteine della membrana mitocondriale interna rivolte verso la matrice):
Ossidazione del piruvato e degli acidi grassi a CO2
Riduzione dei coenzimi NAD+ e FAD a NADH e FADH2
http://www.ncbi.nlm.nih.gov/books/NBK26924/figure/A2606/?report=objectonly
5
06/12/2012
Tre gruppi di reazioni
in regioni distinte dei mitocondri (2)
MEMBRANA MITOCONDRIALE INTERNA:
Trasporto di elettroni dal NADH e dal FADH2 all’O2
Accoppiato alla creazione di FORZA PROTON‐
MOTRICE (gradiente protonico) attraverso la membrana mitocondriale interna.
COMPLESSO F0F1 SULLA MEMBRANA INTERNA:
Utilizzazione dell’energia accumulata nel gradiente protonico per la sintesi dell’ATP.
Mitocondri
IMPORTAZIONE DELLE PROTEINE NELLA MATRICE
Import of proteins into mitochondria
Proteins are targeted for mitochondria by an amino‐terminal presequence containing positively charged amino acids. Proteins are maintained in a partially unfolded state by association with a cytosolic Hsp70 and are recognized by a receptor on the surface of mitochondria. The unfolded polypeptide chains are then translocated through the Tom complex in the outer membrane and transferred to the Tim complex in the inner membrane. The voltage component of the electrochemical gradient is required for translocation across the inner membrane. The presequence is cleaved by a matrix protease, and a mitochondrial Hsp70 binds the polypeptide chain as it crosses the inner membrane, driving further protein translocation. A mitochondrial Hsp60 then facilitates folding of the imported polypeptide within the matrix.
Mitocondri
METABOLISMO
http://www.ncbi.nlm.nih.gov/books/NBK9896/figur
e/A1633/
6
06/12/2012
MOVIMENTO DEL PIRUVATO DAL CITOSOL
VERSO LA MATRICE MITOCONDRIALE (1)
In condizioni aerobiche il piruvato è trasportato all’interno dei mitocondri in scambio con l’ione OH‐ mediante il trasportatore per il piruvato («pyruvate carrier»), un antiporto.
Metabolismo nei mitocondri
(1) DECARBOSSILAZIONE DEL PIRUVATO
http://www.ncbi.nlm.nih.gov/books/NBK22427/; http://course1.winona.edu/sberg/ILLUST/PyruvateSymporter.jpg
MOVIMENTO DEL PIRUVATO DAL CITOSOL
VERSO LA MATRICE MITOCONDRIALE (2)
Il complesso della piruvato deidrogenasi
Le molecole di piruvato prodotte nella glicolisi sono trasportate
attraverso la membrana mitocondriale interna verso la matrice, dove vengono decarbossilate per formare gruppi acetile, con 2 atomi di carbono (‐CH3COO2).
Il gruppo acetile forma in seguito un complesso con il coenzima A per formare acetil‐CoA:
Piruvato + HS‐CoA + NAD+ Æ acetil CoA + CO2 + NADH + H+
La decarbossilazione del piruvato ed il trasferimento del gruppo acetile al CoA sono catalizzati dal complesso multienzimatico gigante della piruvato deidrogenasi nella matrice mitocondriale.
La struttura dell’importante molecola trasportatrice (“Carrier”) attivata
acetil CoA.
In alto un modello “space-filling” della struttura. Un atomo di zolfo (giallo)
forma un legame tioestere con l’acetato. Dato che questo è un legame ad
elevata energia, che rilascia una grande quantità di energia libera quando è
idrolizzato, la molecola di acetato può essere prontamente trasferita ad
altre molecole.
http://www.ncbi.nlm.nih.gov/books/NBK26838/figure/A277/?report=objectonly
7
06/12/2012
Decarbossilazione ossidativa del piruvato
(matrice mitocondriale)
Decarbossilazione del piruvato: collegamento tra la glicolisi e il ciclo di Krebs
Nella decarbossilazione irreversibile del piruvato viene rilasciato CO2 e sono catturati dal NAD elettroni ad alto potenziale di trasferimento.
Il piruvato viene convertito in CO2 (decarbossilato) e in acetilCoA, in un processo che produce anche una molecola di NADH. La reazione della piruvato deidrogenasi ha molti aspetti in comune con il ciclo di Krebs stesso.
Il coenzima A (CoA‐SH) é un trasportatore generale di gruppi acilici attivati in una grande varietà di reazioni. http://www.ncbi.nlm.nih.gov/books/NBK22427/
I prodotti significativi del ciclo di Krebs
Consegna elettroni alla catena respiratoria sulla m.m. interna
Ciclo di Krebs, degli acidi tricarbossilici o dell’acido citrico
Diffonde via dalla cellula
Consegna elettroni alla catena respiratoria sulla m.m. interna
Il ciclo dell’acido citrico in cui gruppi acetilici trasferiti
dal acetil CoA sono ossidati
a CO2
http://www.ncbi.nlm.nih.gov/books/NBK21624/figure/A435
6/?report=objectonly
8
06/12/2012
Appunti sull’ossidazione del succinato
Il succinato è ossidato a fumarato dalla succinato deidrogenasi.
L’accettore di idrogeno è il FAD invece che il NAD+ che viene usato nelle altre tre reazioni di oasido‐reduzione del ciclo.
Nella succinato deidrogenasi l’anello isoalloxazinico del FAD è legato covalentemente alla catena laterale di un’istidina dell’enzima (E‐FAD)
Il FAD è l’accettore dell’idrogeno in questa reazione in quanto lle modificazioni di energia libera sono insufficienti per ridurre il NAD+.
Il FAD è quasi sempre l’accettore di elettroni nelle ossidazioni che rimuovono due atomi di idrogeno da un substrato.
http://www.ncbi.nlm.nih.gov/books/NBK26882/figure/A305/?report=objectonly
http://www.ncbi.nlm.nih.gov/books/NBK22427/
Appunti sulla succinato deidrogenasi
E’ una proteina che contiene tre differenti tipi di centri ferro‐zolfo (Fe‐S).
Diversamente dagli altri enzimi del ciclo dell’acido citrico in quanto è inserita nella membrana mitocondriale interna.
Di fatto, è associata direttamente alla catena di trasporto degli elettroni, il collegamento tra il ciclo dell’acido citrico e la formazione di ATP.
IL FADH2 formato dall’ossidazione del succinato non si dissocia dall’enzima, al contrario del NADH prodotto in altre reazioni di ossido‐riduzione.
Invece, due elettroni sono trasferito dal FADH2 direttamente ai centri Fe‐S dell’enzima.
L’accettore finale di questi elettroni sarà l’ossigeno molecolare.
http://www.ncbi.nlm.nih.gov/books/NBK26882/figure/A306/?report=objectonly
http://www.ncbi.nlm.nih.gov/books/NBK22427/
Riassunto delle reazioni (1)
Stechiometria del ciclo dell’acido citrico
1. Due atomi di carbono entrano nel ciclo nel processo di condensazione di un’unità acetile (dall’acetil CoA) con l’ossaloacetato. Due atomi di carbono escono dal ciclo sotto forma di CO2 nelle decarbossilazioni successive, catalizzate dalla isocitrato deidrogenasi e α‐chetoglutarato deidrogenasi. •
Dati con marcatura isotopica rivelano che gli atomi di carbono che entrano in ogni ciclo non sono quelli che lo abbandonano.
9
06/12/2012
Riassunto delle reazioni (2)
2. Quatro paia di atomi di idrogeno lasciano il ciclo in quattro reazioni di ossidazione. Due molecole di NAD+ sono ridotte nella decarbossilazione ossidativa dell’isocitrato e del α‐chetoglutarato, una molecola di FAD è ridotta nell’ossidazione del succinato e una molecola di NAD+ è ridotta nell’ossidazione del malato.
3. Un composto ad elevato potenziale di trasferimento del fosfato, di solito il GTP, viene generato nella scissione di un legame tioestere del succinil CoA.
Ruolo dell’ossigeno nel ciclo dell’acido citrico
L’ossigeno molecolare non partecipa direttamente al ciclo dell’acido citrico.
Tuttavia, il ciclo opera soltanto in condizioni aerobiche perchè il NAD+ e il FAD possono venire rigenerati nei mitocondri soltanto mediante il trasferimento di elettroni all’ossigeno molecolare.
Mentre la glicolisi può svolgersi sia in modo aerobico che anerobico, il ciclo dell’acido citrico è strettamente aerobico.
Riassunto delle reazioni (3)
4. Sono consumate due molecole di acqua: una nella sintesi del citrato mediante l’idrolisi dell’citril CoA e l’altra nell’idratazione del fumarato.
L’efficacia del ciclo dell’acido citrico può essere aumentata dalla disposizione degli enzimi costituenti. Ci sono sempre più prove sperimentali che gli enzimi siano associati fisicamente uno all’altro per facilitare l’incanlamento deel substrato fra i siti attivi.
E’ sto suggerito il nome di METABOLONE per definire tali complessi multienzimatici.
Il ciclo dell’acido citrico genera elettroni ad alta energia
Il ciclo dell’acido citrico converte gli atomi di carbonio dell’acetil CoA in CO2 che la cellula elimina come prodotto di scarto.
Il punto molto importante è che tale l’ossidazione genera elettroni ad alta energia, trasportati dalle molecole «carrier» attivate, NADH e FADH2.
Questi elettroni ad alta energia sono in seguito trasferiti alla membrana mitondriale interna dove incontrano la catena di trasporto degli elettroni.
La perdita degli elettroni dal NADH e dal FADH2 rigenera il NAD+ e il FAD, necessari per che il metabolismo ossidativo prossegua.
Metabolisamo
CATENA DI TRASPORTO
DEGLI ELETTRONI
Figure 14-10 Molecular Biology of the Cell (© Garland Science 2008)
10
06/12/2012
La riduzione dell’ossigeno ad acqua
avviene in diversi passi (1)
http://www.ncbi.nlm.nih.gov/books/NBK21063/figure/A2492/?report=objectonly
La reazione energeticamente favorevole H2 + ½ O2 Æ H20 viene fatta svolgere in diversi passi in modo che l’energia rilasciata possa essere immagazzinata invece di venire persa nell’ambiente sotto forma di calore.
Prima gli atomi di idrogeno sono separati in protoni ed elettroni.
Gli elettroni passano attraverso una serie di trasportatori di elettroni presenti sulla membrana mitocondriale interna.
In diverse occasioni i protoni e gli elettroni sono ricombinati transitoriamente.
I protoni ritornano permanentemente alla matrice soltanto alla fine della catena di trasporto degli elettroni; nella matrice possono essere usati per neutralizzare le cariche negative create dall’aggiunta finale di elettroni alla molecola di ossigeno.
http://www.ncbi.nlm.nih.gov/books/NBK26904/#A2539
La riduzione dell’ossigeno ad acqua
avviene in diversi passi (2)
Il processo di trasporto degli elettroni inizia quando l’ione idruro (ione carico negativamente dell’idrogeno: H‐) è rimosso dal NADH (per rigenerare NAD+) ed è convertito in un protone e due elettroni (H‐ ÆH+ + 2e‐).
I due elettroni sono passati al primo di una serie di più di 15 diversi trasportatori di elettroni nella catena respiratoria.
Gli elettroni iniziano con energia molto elevata e progressivamente la perdono mentre passano lungo la catena.
Nella maggior parte dei passi gli elettroni passano da un ione metallico ad un altro (ciascuno di questi ioni è strettmanete legato ad una molecola proteica che altera l’affinità verso gli elettroni del metallo)
Mentre gli elettroni si muovono lungo la catena respiratoria, l’energia viene immagazzinata sotto forma di gradiente protonico attraverso la membrana interna (1)
La fosforilazione ossidativa è possibile dato che i trasportatori di elettroni sono strettamente associati alle molecole proteiche.
Le proteine guidano gli elettroni lungo la catena respiratoria in modo che essi si muovano in sequenza da un complesso enzimatico al successivo.
Il trasferimento degli elettroni è accoppiato:
alla captazione e rilascio orientati di H+
a modificazioni allosteriche nelle pompe protoniche coinvolte nella conversione di energia.
Il risultato complessivo è il pompaggio di H+ attraverso la membrana interna – dalla matrice allo spazio intermembrane –
pilotato dal flusso energeticamente favorevole di elettroni .
La riduzione dell’ossigeno ad acqua
avviene in diversi passi (3)
La maggior parte delle proteine coinvolte sono raggruppate in tre grandi complessi enzimatici respiratori, ciascuno dei quali contiene proteine transmembrana che trattengono i complessi fermamamente nella membrana mitocondriale interna.
Ogni complesso della catena ha un’affinità verso gli elettroni superiore a quella del suo predecessore, e quindi gli elettroni passano in sequenza da un complesso all’altro finchè non sono trasferiti all’ossigeno, che ha l’affinità per gli elettroni superiore a tutti gli altri.
Mentre gli elettroni si muovono lungo la catena respiratoria, l’energia viene immagazzinata sotto forma di gradiente protonico attraverso la membrana interna (3)
Il movimento dell’H+ ha due conseguenze principali:
Genera un gradiente di pH attraverso la membrana interna, con pH superiore nella matrice rispetto allo spazio intermembrana (e al citosol, dato che la membrana esterna è permeabile ).
Genera un gradiente di voltaggio (potenziale di membrana) attraverso la membrana interna, in cui l’interno è negativo e l’esterno positivo (come risultato del flusso netto di ioni positivi).
11
06/12/2012
Mentre gli elettroni si muovono lungo la catena respiratoria, l’energia viene immagazzinata sotto forma di gradiente protonico attraverso la membrana interna (4)
Il gradiente di pH (ΔpH) pilota il ritorno degli H+ verso la matrice, così rinforzando l’effetto del potenziale di membrana (ΔV), che agisce per attrarre qualsiasi ione positivo verso la matrice e quindi per spingere gli ioni negativi fuori.
Complessivamente, il ΔpH e il ΔV costituiscono un gradiente elettrochimico protonico.
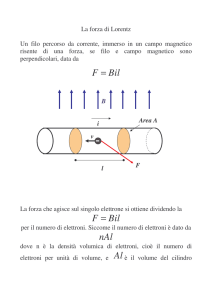
Il gradiente elettrochimico protonico esercita una forza proton‐motrice (misurata in millivolts, mV)
La catena respiratoria consiste di quattro complessi:
tre pompe protoniche e un collegamento fisico
con il ciclo dell’acido citrico (1)
Gli elettroni sono trasferiti dal NADH all’O2 mediante una catena di tre grandi complessi proteici chiamati NADH‐Q ossidoreduttasi, Q‐citocromo C ossido‐reduttasi e citocromo C ossidasi.
http://www.ncbi.nlm.nih.gov/books/NBK22505/
La catena respiratoria consiste di quattro complessi:
tre pompe protoniche e un collegamento fisico
con il ciclo dell’acido citrico (1)
Il fluire degli elettroni attraverso questi complessi transmembrana porta al
trasporto di protoni attraverso la membrana mitocondriale interna.
Gli elettroni sono portati dalla NADH-Q ossidoreduttasi alla Q-citocromo C
ossidoreduttasi, il secondo complesso della catena, dalla forma ridotta del
coenzima Q (Q) anche noto come ubichinone perchè è un chinone
ubiquitarii nei sistemi biologici.
http://www.columbiamitodiagnostics.org/tests/coen
zymeq10.html
http://www.ncbi.nlm.nih.gov/books/NBK22505/
Coenzima Q
Derivato chinonico con una lunga coda isoprenoide. Il numero di unità isopreniche di 5 atomi di carbono dipende dalla specie. I mammiferi più comuni contengono 10 unità isopreniche (coenzima Q10).
I chinoni possono esistere in tre stati di ossidazione. Nello stato totalmente ossidato
(Q) il coenzima Q ha due gruppi
chetonici. L’aggiunta di un elettrone e di un protone porta
alla forma semichinonica (QH∙).
Il semichinone è deprotonata in modo relativamente facile formando un anione radicalico
semichinonico (Q∙‐). L’aggiunta di un secondo elettrone e di un protone genera il diidroubichinone (QH2), la forma totalmente ridotta del coenzima Q che conserva I suoi protoni più strettamente.
Perciò, per i chinoni, le reazioni di trasferimento di energia sono accoppiate al legame e rilascio di un protone, una proprietà che è fondamentale per il transporto dei protoni
attraverso la membrana. La catena respiratoria consiste di quattro complessi:
tre pompe protoniche e un collegamento fisico
con il ciclo dell’acido citrico (2)
Struttura del coenzima Q (CoQ) o ubichinone, illustrante la sua capacità fi trasportare
due protoni e due elettroni. L’ubichinone è un chinone idrofobico che diffonde rapidamente all’interno della membrana mitocondriale interna.
L’ubichinone trasporta gli elettroni dal FADH2, generati dalla succinato deidrogenasi nel ciclo dell’acido citrico, alla Q‐
citocromo C ossidoreduttasi, generata mediante la succinato Q‐reduttasi.
Il citocromo C, una piccola molecola solubile, fa da navetta per gli elettroni dalla Q‐citocromo C ossidoreduttasi alla citocromo C ossidasi, la componente finale della catena e quella che catalizza la riduzione dell’O2 ad H2O.
12
06/12/2012
Struttura del citocromo c di cuore di cavallo
I citocromi sono emoproteine legate a membrane che contengono gruppi
eme e sono coinvolte nel trasporto di elettroni
La catena respiratoria consiste
di quattro complessi:
tre pompe protoniche e un collegamento fisico
con il ciclo dell’acido citrico (3)
La NADH‐Q ossidoreduttasi, la succinato‐Q reduttasi, la Q‐
citocromo c ossidoreduttasi e la citocromo c ossidasi sono anche noti come Complessi I, II, III e IV, rispettivamente.
La succinato‐Q reduttasi (complesso II), a differenza degli altri complessi, non pompa elettroni.
http://en.wikipedia.org/wiki/Cytochrome
http://www.ncbi.nlm.nih.gov/books/NBK26904/figure/A2535/?report=objectonly
Gli elettroni ad elevato potenziale del NADH entrano nella catena respiratoria a livello della NADH‐Q ossidoreduttasi
Stati di ossidazione delle Flavine
Gli elettroni del NADH entrano nella catena a livello della NADH‐Q ossidoreduttasi (NADH deidrogenasi), un enorme enzima /pompa protonica con almeno 34 catene polipeptidiche.
La reazione catalizzata da questo enzima sembra essere: Il passo iniziale è il legame di NADH e il trasferimento dei suoi due
elettroni ad elevata energia al gruppo prosptetico FMN (flavina
mononucleotide) di questo complesso per dare la forma ridotta FMNH2.
Come i chinoni, anche il FMN può accettare un elettrone invece che due
per formare un radicale semichinonico.
L’accettore di elettroni del FMN, l’anello isoalloxazinico, è identico a quello
del FAD.
Gli elettroni vengono allora trasferiti dal FMNH2 ad una serie di centri FeS, il secondo tipo di gruppi prostetici della NADH-Q ossidoreduttasi.
La riduzione della flavina mononucleotide (FMN) FMNH2 procede
mediante un intermediario semichinonico.
http://www.ncbi.nlm.nih.gov/books/NBK22505/figure/A2508/?report=objectonly
I centri Fe‐S nelle proteine ferro‐zolfo (dette anche proteine a ferro non‐emico) giocano un ruolo chiave in una gran diversità di reazioni biologiche di riduzione. Gli elettroni dei centri Fe‐S della NADH‐Q ossidoreduttasi sono convogliati al coenzima Q portando al pompaggio di quattro ioni di idrogeno dalla matrice del mitocondrio.
La NADH‐Q ossidoreduttasi contiene clusters di tipo 2Fe‐2S e di tipo 4Fe‐4S.
Gli atomi di ferro di questi complessi passano ciclicamente fra stati Fe2+ (ridotto) o Fe3+ (ossidati).
Al contrario dei chinoni e delle flavine, i centri Fe‐S di solito subiscono ossidazione‐riduzione senza rilasciare o legare protoni.
13
06/12/2012
Il diidroubichinone è il punto di ingresso degli elettroni del FADH2 delle flavoproteine
L’enzima del ciclo dell’acido citrico succinato deidrogenasi, che genera FADH2 con l’ossidazione del succinato in fumarato, fa parte del complesso succinato‐Q reduttasi (complesso III) una proteina integrale di membrana della membrana mitocondriale interna.
Il FADH2 non lascia il complesso [gruppo prostetico]; invece gli elettroni sono trasferiti a centri Fe‐S e successivamente al Q per dare entrata nella catena di trasporto degli elettroni.
Il complesso succinato‐Q reduttasi, e altri enzimi che trasferiscono gli elettroni del FADH2 a Q, al contrario della NADH‐Q ossidoreduttasi, non trasportano protoni.
Perciò, si forma meno ATP dall’ossidazione del FADH2 che non del NADH: Quinones
Each of these electron carriers in the respiratory chain picks up one H+ from the aqueous environment for every electron it accepts, and it can carry either one or two electrons as part of a hydrogen atom (yellow). When it donates its electrons to the next carrier in the chain, these protons are released. In mitochondria the quinone is ubiquinone (coenzyme Q), shown here; the long hydrophobic tail, which confines ubiquinone to the membrane, consists of 6 to 10 five‐carbon isoprene units, depending on the organism. The corresponding electron carrier in plants is plastoquinone, which is almost identical. For simplicity, both ubiquinone and plastoquinone will normally be referred to as quinone and abbreviated as Q.
http://www.ncbi.nlm.nih.gov/books/NBK28380/figure/A3520/
Gli elettroni fluiscono dall’ubichinolo al citocromo C tramite la Q‐citocromo C ossidoreduttasi
La seconda delle tre pompe protoniche della catena respiratoria è la Q‐
citocromo c ossidrduttasi (Complesso III o citocromo ossidasi).
Un citocromo è una proteina che trasferisce elettroni che contiene un gruppo prostetico eme.
Il ferro del gruppo prostetico alterna fra uno stadio ridotto ferroso (Fe2+) e uno stato ossidato ferrico (Fe3+) durante il trasporto degli elettroni.
La funzione della Q‐citcromo C ossidoreduttasi è quella di catalizzare la trasferta di elettroni dal QH2 al citocromo c ossidato (Cyt c), una proteina solubile in acqua, e concomitantemente pompare elettroni fuori dalla matrice mitocondriale.
http://www.ncbi.nlm.nih.gov/books/NBK22505/
http://www.ncbi.nlm.nih.gov/books/NBK26904/figure/A2540/?report=objectonly
Un gradiente protonico fornisce l’energia
per la sintesi dell’ ATP (1)
Il flusso di elettroni dal NADH all’O2 è un processo esergonico:
Questo processo è accoppiato alla sintesi di ATP, un processo endergonico:
Mitocondri
SINTESI DELL’ATP
La sintesi dell’ATP è svolta da un complesso molecolare presente nella membrana
mitocondriale interna:
Noto precedentemente come F 1 F 0 ATPase perchè è stato scoperto mediante
la sua catalisi del processo inverso, l’idrolisi dell’ATP.
ATP sintasi è il nome preferibile perchè enfatizza il suo ruolo reale nel
Viene anche chiamato Complesso V.
http://www.ncbi.nlm.nih.gov/books/NBK22388/
14
06/12/2012
Un gradiente protonico fornisce l’energia
per la sintesi dell’ ATP (2)
In 1961, Peter Mitchell propose che il
trasporto degli elettroni e la sintesi
dell’ATP fossero accoppiati mediante un gradiente protonico attraverso la membran mitocondriale interna, piuttosto che mediante un intermediario
covalente ad alta energia.
Nel suo modello, il trasferimento di elettroni lungo la catena respiratoria
portava al pompaggio di protoni dalla
matrice allo spazio fra la membrana
mitcondriale interna e quella esterna
La concentrazione di H+ diventa minore
nella matrice, e viene generato un campo elettrico in cui il lato della
matrice è negativo rispetto allo spazio
intermembrane. [n° protoni inferiore
rispetto allo spazio intermembrane]
http://www.ncbi.nlm.nih.gov/books/NBK22388/
http://www.nobelprize.org/nobel_prizes/chemistry/laureates/1978/
Structure of ATP Synthase
Figure 18.27. Structure of ATP
Synthase. A schematic structure is
shown along with detailed structures of
the components for which structures
have been determined to high
resolution. The P-loop NTPase domains
of the α and β subunits are indicated by
purple shading.
http://www.ncbi.nlm.nih.gov/books/NBK22388/figure/A2532/?report=object
only
15
06/12/2012
http://www.stanford.edu/group/hopes/treatmts/ebuffer/j1.html
Il gradiente protonico pilota la sintesi dell’ATP (1)
Il gradiente elettrochimico protonico attraverso la membrana mitocondriale interna è il processo fondamentale della fosforilazione ossidativa.
Questa è svolta mediante un enzima legato alla membrana interna, l’ATP sintasi.
L’ATP sintasi crea una via idrofilica attraverso la membrana interna che permette il flusso di protoni lungo il loro gradiente elettrochimico.
Mentre i protoni si incanalano lungo l’ATP sintasi, essi sono usati per pilotare la reazione energeticamente sfavorevole fra l’ADP e il Pi che permette il ripristino dell’ATP.
http://www.ncbi.nlm.nih.gov/books/NBK26904/figure/A2549/?report=objectonly
Natura elettrochimica del gradiente protonico
CHEMIOSMOSI (1)
Dato che i protoni sono carichi positivamente, il gradiente protonico stabilito attraverso la membrana mitocondriale interna ha componenti sia chimiche che elettriche. La componente chimica è la concentrazione di protoni, o gradiente protonico, che corrisponde ad una concentrazione circa dieci volte superiore di protoni nel versante citosolico della membrana mitcondriale interna (differenza di una unità di pH). Inoltre, vi è un potenziale elettrico attraverso la membrana, che deriva dall’aumento netto di cariche positive sul versante citosolico. http://www.ncbi.nlm.nih.gov/books/NBK9885/figure/A1647/?report=objectonly
16
06/12/2012
CHEMIOSMOSI (2)
Il NADH e il FADH2 trasportano protoni (H+) ed elettroni (e‐) alla catena di trasporto di elettroni localizzata sulla membrane mitocondriale interna. L’energia del trasferimento di elettroni lungo la catena trasporta dei protoni attraverso la membrana crea un gradiente elettrochimico. Man mano che i protoni che si accumulano seguono il gradiente elettrochimico di ritorno attraverso la membrane lungo un complesso ATP sintasi, il movimento dei protoni fornisce l’energia per sintetizzare ATP a partire dall’ADP e fosfato. Alla fine del sistema di trasporto di elettroni, due protoni, due elettroni e una metà di una molecola di ossigeno si combinano per formare acqua. Poichè l’ossigeno è l’accettore finale degli elettroni, il processo è detto respirazione aerobica. http://www.ncbi.nlm.nih.gov/books/NBK26894/figure/A2512/?report=objectonly
The ATP synthase is a reversible coupling device that can convert the
energy of the electrochemical proton gradient into chemical-bond energy, or
vice versa
http://www.ncbi.nlm.nih.gov/books/NBK26894/figure/A2525/?report=objectonly
Mitocondri
TRASPORTATORI E NAVETTE SULLE MEMBRANE MITOCONDRIALI
http://course1.winona.edu/sberg/ILLUST/PyruvateSymporter.jpg
17
06/12/2012
Trasportatori sulla membrana mitocondriale interna
Trasporto di metaboliti
attraverso la membrana mitocondriale interna (1)
http://www.ncbi.nlm.nih.gov/books/NBK21528/fig
ure/A4418/
http://www.ncbi.nlm.nih.gov/books/NBK9885/figure/A1650/
Trasporto di metaboliti
attraverso la membrana mitocondriale interna (2)
Il trasporto di piccolo molecule attraverso la membrane interna è mediato da proteine di trasporto che attraversano la membrana e guidato dal gradiente elettrochimico.
Ad esempio, l’ATP viene esportato dai mitocondri verso il citosol da un trasportatore che lo scambia con l’ADP. La componente di voltaggio del gradiente elettrochimico guida questo scambio: l’ATP porta una maggiore carica negativa (‐4) dell’ADP (‐3), e perciò l’ATP é esportato dalla matrice mitocondriale al citosol mentre l’ADP viene importato verso i mitocondri. Viceversa, il trasporto del fosfato (Pi) e del piruvato é accoppiato ad uno scambio con ioni idrossilici (OH‐); in questo caso, la componente di pH del gradiente elettrochimico guida l’esportazione degli ioni idrossilici, accoppiata al trasporto di Pi e piruvato verso i mitocondri.
Glycerol 3-Phosphate Shuttle
Electrons from NADH can enter the mitochondrial electron transport chain by
being used to reduce dihydroxyacetone phosphate to glycerol 3-phosphate.
Glycerol 3-phosphate is reoxidized by electron transfer to an FAD prosthetic
group in a membrane-bound glycerol 3-phosphate dehydrogenase.
Subsequent electron transfer to Q to form QH2 allows these electrons to enter
the electron-transport chain.
Mitocondri
NAVETTE PER IL NADH FORMATO NEL CITOSOL
Mitocondri
ULTRASTRUTTURA
http://www.ncbi.nlm.nih.gov/books/NBK22470/figure/A2548/
18
06/12/2012
Cellule parietali dello stomaco: secernono acido cloridrico
Tubuli renali
http://www.anatomyatlases.org/MicroscopicAnatomy/Section10/Plate1019
1.shtml
Tubuli distali del nefrone: riassorbono sodio dall’ultrafiltrato contro gradiente
http://lynlaukimdak.wikispaces.com/12.+Urology
JC‐1 Dye ‐ Mitchondrial Membrane Potential Probe
Mitocondri
FLUOROCROMI
http://www.invitrogen.com/site/us/en/home/Products-and-Services/Applications/Cell-Analysis/Flow-Cytometry/CellHealth-and-Viability-Assays-for-Flow-Cytometry/Apoptosis-Assays-for-Flow-Cytometry/JC-1-Dye-MitochondrialMembrane-Potential-Probe.html
19
06/12/2012
Membrane mitocondriali – JC‐1
Fig. 1. Cultured human bronchial smooth muscle cell stained with
mitochondrial membrane potential indicator JC-1. Red area correspond to
energized mitochondria with highly negative membrane potential. PlanApo
x63 1.4 oil immersion objective, CARV II preset filters/mirror combinations
- GREEN and RED.
http://www.pharmaceutical-int.com/article/flexible-imaging.html
Potential-dependent staining of mitochondria in CCL64 fibroblasts by JC-1
(T3168). The mitochondria were visualized by epifluorescence microscopy
using a 520 nm longpass optical filter. Regions of high mitochondrial
polarization are indicated by red fluorescence due to J-aggregate formation
by the concentrated dye. Depolarized regions are indicated by the green
fluorescence of the JC-1 monomers. The image was contributed by Lan Bo Chen, Dana Farber
Cancer Institute, Harvard Medical School.
http://www.invitrogen.com/site/us/en/home/support/Research-Tools/Image-Gallery/ImageDetail.alternateID.g000490.html
Βeta‐ossidazione degli acidi grassi
Mitocondri_seminario
Β‐ossidazione degli acidi grassi
Anche gli enzimi che degradano gli acidi grassi derivati
dai trigliceridi producono acetil CoA nei mitocondri. Ogni molecola di acido grasso, sotto forma attivata di acil CoA, viene degradata completamente mediante un ciclo di reazioni che scinde due carboni alla volta a partire dall’estremità carbossilica, generando una
molecola di acetil CoA ad ogni giro del ciclo. In questo processo sono prodotte una molecola di NADH e una molecola di FADH2. Gli acidi grassi sono ossidati sopratutto nei mitocondri.
[N.B. L’inizio della degradazione degli acidi grassi a
catena molto lunga si svolge nei perossisomi].
Gli acidi grassi debbono essere attivati prima di entrare
nella matrice mitondriale.
L’adenosina trifosfato (ATP) pilota la formazionedi un
legame tioester fra il gruppo carbossilico di un acido
grasso e il gruppo sulfidrilico del CoA.
Questo processo di attivazione si svolge nella membrana
mitocondriale esterna, dove è catalizzato dalla acil CoA
sintetasi.
http://www.ncbi.nlm.nih.gov/books/NBK26882/figure/A302/?report=objectonly
http://www.ncbi.nlm.nih.gov/books/NBK22581/
20
06/12/2012
La Carnitina trasporta gli acidi grassi a catena lunga attivati
verso la matrice mitocondriale
Gli acidi grassi sono attivati nella membrana mitocondriale esterna
mentre sono ossidati nella matrice mitocondriale. E’ necessario un meccanismo speciale per trasportare le molecole a lunga
catena di acil‐Coa attraverso la membrana mitocondriale interna.
Gli acidi grassi attivati sono trasportati attraverso la membrana mediante
cconiugazione con la carnitina, un alcool zwitterionica.
Il gruppo acilico è trasferito dall’atomo di zolfo del CoA al gruppo
idrossilico della canitina formando acil carnitina. Questa reazione è catalizzata dalla carnitine aciltrasferasiche è legata alla
membrana mitocondriale esterna. http://lpi.oregonstate.edu/infocenter/othernuts/carnitine/structure.html
http://www.ncbi.nlm.nih.gov/books/NBK22581/
Acil Carnitine Traslocasi
L’entrata della acilcarnitina nella matrice mitocondriale è mediata da una
traslocasi. La carnitina ritorna al lato citosolico della membrana mitocondriale
internamediante scambio con l’acil carnitina.
Trasporto dell’ Acil-CoA dal citosol alla matrice mitocondriale
http://en.wikipedia.org/wiki/Carnitine-acylcarnitine_translocase
http://www.ncbi.nlm.nih.gov/books/NBK22581/figure/A3055/?report=objectonly
Schema dell’ossidazione degli acidi grassi
Oxidation of fatty acids in mitochondria
Four enzyme-catalyzed reactions convert a
fatty acyl CoA molecule to acetyl CoA and a
fatty acyl CoA shortened by two carbon
atoms. Concomitantly, one NAD+ molecule is
reduced to NADH and one FAD molecule is
reduced to FADH2. The cycle is repeated on
the shortened acyl CoA until fatty acids with
an even number of carbon atoms are
completely converted to acetyl CoA. Fatty
acids with an odd number of C atoms are
rare; they are metabolized to one molecule of
propionyl CoA and multiple acetyl CoA
molecules.
http://www.ncbi.nlm.nih.gov/books/NBK21624/figure/A4361/?report=
objectonly
http://www.ncbi.nlm.nih.gov/books/NBK28177/figure/A2964/
21