Microrganismi
Procarioti
Eubatteri-Archibatteri
Eucarioti
Funghi-Protozoi (alghe)
Dimensioni
~ 1-10 µm
~ 10-100 µm
genoma
Nucleoide diffuso
Nucleo con membrana
Crosoma
In genere circolare
lineari
Trascrizione
Traduzione
citoplasma
Nucleo
citoplasma
Ribosomi
70S (50+30)
80S (60+40)
Strutture interne
rare
Organelli delimitati da
membrane e citoscheletro
Mobilità
Flagelli di flagellina
Flagelli e ciglia di tubulina
Mitocondri
Assenti
Presenti (con rarissime
eccezioni)
Cloroplasti
Assenti
(alghe)
Parete cellulare
Presente
(funghi-chitina)
Divisione
Principalmente schizogonica
Mitosi/meiosi
(fissione/gemmazione)
Steroli in membrana
Generalmente assenti
colesterolo
la forma dei batteri viene spesso riferita a tre modelli principali
SFERE
SPIRALI
BASTONCELLI
Ma le variazioni sulle forme essenziali sono moltissime
E ci possono essere disposizioni diverse
Tetrade
Grappolo
irregolare
Sarcina
Diplo-
Strepto-
I batteri sono molto più piccoli delle
cellule eucariotiche
le loro dimensioni sono dell’ordine dei µm
(in media 1-10)
Circa 0,3 µm i più piccoli (micoplasmi)
Nanobatteri???!
Proteine complessate con sali
precipitati!
I più grandi..
Epulopiscium fishelsonii
(30-520 µm)
Thiomargarita namibiensis
(≈ 700 µm)
Dimensioni: Le dimensioni ridotte dei batteri sono
convenienti per lo scambio con l’ambiente esterno
r = 1μm
r = 2μm
superficie (4πr2) = 12,6 μm2
Volume 4/3πr3 = 4,2 μm3
Rapporto area/volume = 3
superficie (4πr2) = 50,3 μm2
Volume 4/3πr3 = 33,5 μm3
Rapporto area/volume = 1,5
C’è il trucco….
Struttura
(procariotica)
ribosomi
(plasmide)
pili
parete
Matrice
citoplasmatica
(capsula)
nucleoide
Membrana
citoplasmatica
(flagello)
Diversità strutturale: i procarioti non sono meno
organizzati, sono organizzati diversamente
PROCARIOTI
Archibatteri
(Archea)
Eubatteri
F-met
INIZIO DELLA
TRADUZIONE
Met
assenza di istoni
DNA
istoni
peptidoglicano
PARETE
assenza di
peptidoglicano
Legami estere
LIPIDI DI
MEMBRANA
Legami etere
CITOPLASMA
MATRICE
70% = H2O
Lipidi, Proteine, Sali
CITOSCHELETRO
BATTERICO
Scoperto relativamente di recente
proteine che si assemblano in strutture filamentose in
vitro e formano strutture ordinate intracellulari in vivo
divisione cellulare
determinazione della polarità della cellula,
regolazione della forma cellulare
ripartizione dei plasmidi…
OMOLOGHI DELLA TUBULINA
FtsZ :formazione del setto
mutanti che non producono FtsZ non formano setti
di divisione e danno forme filamentose
OMOLOGHI DELL’ACTINA
MreB
mantenimento della forma non
sferica (bastoncelli e spirilli)
mutanti di E. coli che non producono MreB
diventano di forma sferica
ParM : segregazione dei plasmidi
Omologo funzionale dei
filamenti intermedi
CreS: Crescentina
(Caulobacter crescentus)
Ha domini strutturali simili a quelli delle proteine
che costituiscono i filamenti intermedi
I mutanti che non la producono perdono
la tipica forma ricurva
Componenti del citoscheletro
esclusivamente procariotici
Gruppo “MinD-ParA
Min(DCE) posizionamento del setto
Formano polimeri elicoidali
ParA (ATPasi): posizionamento e
segregazione dei plasmidi nelle cellule figlie
FtsZ si trova nella quasi totalità dei
batteri
MreB è presente in tutti i
batteri non sferici
Crescentin è presente in almeno 1
batterio con morfologia “curva”
NUCLEOIDE
La regione in cui si
addensa è il “nucleoide”
o “regione nucleare”
A differenza di quello eucariotico, il DNA
procariotico è disperso nel citoplasma
Nella maggior parte dei casi i batteri hanno un solo cromosoma
circolare, ma questo non è sempre vero
Vibrio cholerae
ha due cromosomi..
Burkholderia cepacia
Ne ha tre..
il cromosoma e i plasmidi di
Borrelia burgodorferi sono lineari
Le dimensioni del cromosoma
batterico possono variare
~4 mgb
la maggior parte dei
batteri (es. E. coli )
S. coelicolor
0,65 mgb
Endosimbionti come
Buchnera apidicola
0,58 mgb
Il più piccolo :
Micoplasma genitalium
10 mgb
In ogni caso, la lunghezza del
cromosoma “rilassato”
(circa 1 mm)
È di 500-1000 volte superiore a
quella della cellula batterica
Nel citoplasma il cromosoma è
attorcigliato in “superavvolgimenti”
In natura il DNA è in genere
superavvolto negativamente
Questo significa che è avvolto intorno al
suo asse, in direzione CONTRARIA a
quella della doppia elica destrorsa
L’introduzione o l’eliminazione dei giri di
superavvolgimento è opera delle
“topoisomerasi”
Topoisomerasi I
rimuovono il superavvolgimento
Topoisomerasi II (DNA girasi)
Introducono un giro di avvolgimento
In natura il DNA è in genere
superavvolto negativamente
Questo significa che è avvolto intorno
al suo asse, in direzione CONTRARIA
a quella della doppia elica destrorsa
L’introduzione o l’eliminazione dei giri
di superavvolgimento è opera delle
“topoisomerasi”
Topoisomerasi I
rimuovono il superavvolgimento
Tagliando UN SOLO FILAMENTO
E RUOTANDOLO INTORNO
ALL’ALTRO
Topoisomerasi II (DNA girasi)
Introducono un giro di avvolgimento
Topoisomerasi II: INTRODUCE un giro di avvolgimento
si lega al DNA
Lo taglia..
(rottura della DOPPIA
ELICA)
E lo rilega
incrociandolo
Il progressivo aumento del grado di superavvolgimento porta al
formarsi di una struttura sempre più compatta
superavvolto
rilassato
Il DNA lineare
(es. cromosomi eucariotici)
Mantiene il superavvolgimento anche in
presenza di una rottura grazie alle proteine
che vi sono legate
Il cromosoma procariotico circolare potrebbe
passare allo stato rilassato ad ogni rottura
questa eventualità è prevenuta da un tipo di organizzazione
In domini di superavvolgimento isolati
Una rottura in un filamento non
pregiudica lo stato degli altri
quando una cellula procariotica
si replica rapidamente
Vi si possono trovare anche
diverse copie del cromosoma e
copie incomplete
Questo dipende dal particolare modo di
replicarsi del cromosoma stesso
Oltre al nucleoide, nei batteri
possono essere presenti
PLASMIDI
Circolari, con poche eccezioni
Alcuni (CONIUGATIVI) possono promuovere il
proprio trasferimento da una cellula all’altra
Molecole di DNA a doppio
filamento, in grado di replicarsi
autonomamente
I geni presenti sui plasmidi conferiscono nuove
proprietà ai loro ospiti
BATTERIOCINE
METABOLICHE
DI
RESISTENZA
DI
VIRULENZA
il citoplasma è
molto ricco di RNA
Sito di legame
con l’aa
3’
5’
rRNA
mRNA,
tRNA
le specie di RNA sono le stesse
che si trovano negli eucarioti
Legami
idrogeno
C
i ribosomi batterici misurano circa 15x20 nm
e sono formati da due subunita’
G
A
anticodon
50S = 34 proteine
due molecole di RNA 23S e 5S
30
50
Ansa
anticodon
30S = 21 proteine
una molecola di RNA 16S
S= Svedberg (costante di sedimentazione)
70s
il numero di ribosomi è particolarmente elevato: in una cellula in crescita
attiva l’RNA ribosomale può raggiungere il 15% della massa cellulare
Sintesi delle proteine
citoplasmatiche
Sintesi delle proteine
destinate alla secrezione
si trovano dispersi nella matrice
citoplasmatica
o adesi alla membrana
mRNA: nei procarioti contiene le “istruzioni” per la sintesi
di uno ma anche diversi polipeptidi
MONOCISTRONICO
POLICISTRONICO
Oltre alle regioni non tradotte al 5’ e al 3’ nei messaggeri
policistronici possono esserci intervalli tra le ORF
nel citoplasma di alcuni procarioti possono
essere presenti anche altre strutture
INCLUSIONI DI
MATERIALI DI RISERVA
VESCICOLE
GASSOSE
ENDOSPORE
MAGNETOSOMI
CARBOSSISOMI
CO2
Le vescicole gassose
Permettono ai batteri che le possiedono di galleggiare :
sono impermeabili all’acqua e permeabili ai gas
Due bottiglie completamente
riempite di una sospensione di
cianobatteri, che galleggiano
Colpendo il tappo con un
martello, le vescicole
collassano e rilasciano il gas
le cellule batteriche affondano
La membrana delle vescicole è rigida e formata da due
proteine: GvpA e GvpC
GvpA è un foglietto beta rigido che forma l’ossatura;
GvpC (a-elica) forma nastri che legano l’ossatura
La presenza delle vescicole diminuisce la densità della
cellula e ne favorisce il galleggiamento
Il rapporto percentuale tra le vescicole e
il contenuto citoplasmatico determina il
grado di galleggiabilità e la posizione
delle cellule batteriche
Alcuni batteri (es. Cianobatteri) possono modificare la propria posizione
collassando le vescicole o producendo una maggior quantità di polimeri densi
SOSTANZE DI RISERVA
! *#
!
nell’esistenza dei microrganismi
abbondanza e carestia si alternano continuamente
l’accumulo di sostanze di riserva è un fenomeno molto diffuso
Alcune sono molto
comuni
PHB Gly
Glicogeno
amido
S PO4
poli-βidrossibutirrato
(PHB)
PHB
NO3
S
polifosfati
Gly
PO4
Altre presenti solo in alcune specie e
correlate con metabolismi particolari
(zolfo, nitrati)
amido
PHB
glicogeno
sono riserve di carbonio
polimerico
-Chiaramente visibile al
microscopio ottico
-Presente solo nei procarioti
-Invisibili al microscopio ottico
-Presenti anche negli eucarioti
R
il poli-β-idrossibutirrato
C
H C
H2
O
O
E’ un composto lipidico della classe
dei poli-β-idrossi-alcanoati (PHA)
Le sue inclusioni sono circondate da un
monostrato di fosfolipidi
CIANOFICINA
Unico accumulo di azoto
biologico che si conosca
Polimero di acido
aspartico
In cui a ogni residuo ASP è
legato un residuo di arginina
ASP
ASP
ASP
ARG
ARG
ARG
Molecole lineari di polimeri
di unità fosfato
Formano la VOLUTINA
(granuli metacromatici)
volutina
I batteri che ossidano lo
zolfo lo immagazzinano
spesso nel citoplasma
Alcuni batteri (Thioploca, Thiomargarita)
ossidano lo zolfo usando NO3 al posto dell’ossigeno
Ma H2S e NO3 non coesistono nell’ambiente
NO3NO3
H2S
Immagazzinano NO3 in un vacuolo
utilizzando lo zolfo di riserva
Immagazzinano S0 nel citoplasma
utilizzando il nitrato di riserva
•Thiomargarita: cellule in divisione
microscopio confocale: il citoplasma (verde) è marginale e contiene granuli di
zolfo (bianco). La zona nera al centro è il vacuolo di nitrato
I magnetosomi
Sono particelle che permettono ai magnetobatteri di
orientarsi lungo il campo magnetico terrestre
Il loro numero e la loro disposizione
variano da specie a specie
La disposizione è mantenuta da proteine
del citoscheletro simili all’actina
formazione del magnetosoma
Membrana del magnetosoma (MM)
Fe3+
Fe2+
Fe3O4
CARBOSSISOMI
inclusioni proteiche
presenti in alcuni batteri che
fissano CO2
Aggregati cristallini dell’enzima RuBiSco
(ribulosio 1,5-difosfato carbossilasi)
ENTEROSOMI
analoghi dei carbossisomi
presenti solo in batteri che
NON fissano la CO2
aggregati cristallini di enzimi per l’utilizzo di
etanolamina e propanediolo
più piccoli e irregolari dei carbossisomi
la loro presenza è legata a quella del
substrato
ENDOSPORE
(spore dentro le cellule)
Le endospore sono frutto di una divisione cellulare ineguale
permettono ad alcune specie di sopravvivere anche molto a
lungo in situazioni critiche
Non hanno una funzione replicativa ma di sola
resistenza nei confronti di:
CALORE
ESSICCAMENTO
INVECCHIAMENTO
RAGGI UV
DISINFEZIONE
Le endospore hanno una struttura
caratteristica degli involucri esterni
MEMBRANA
PLASMATICA
PARETE DEL
CORE
TUNICA
PROTEICA
CORE
ESOSPORIO
CORTEX
Le endospore sono molto rifrangenti e chiaramente
visibili al microscopio ottico
La rifrangenza è dovuta al basso
contenuto di acqua (10-30% di quello di
una cellula vegetativa)
Oltre alla maggiore densità,
il core sporale
Ha un pH minore
(1 unità) del normale
Contiene acido dipicolinico
chelato con ioni Ca2+
(termoresistenza)
Contiene SASP
(Small Acid Soluble, spore specific Proteins)
Che legano e proteggono il DNA
C
C
O
O
Ca
C
C
O
O
Il processo di sporulazione
Si innesca in condizioni di carenza di nutrienti o altri
tipi di stress ambientali o in colture molto affollate
VII- rilascio
II-Divisione asimmetrica
VI- maturazione
V- tuniche
III- Protoplasto prespora
IV- Parete e cortex
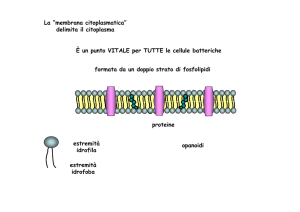
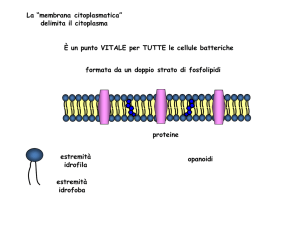
La “membrana citoplasmatica”
delimita il citoplasma
È un punto VITALE per TUTTE le cellule batteriche
formata da un doppio strato di fosfolipidi
proteine
estremità
idrofila
estremità
idrofoba
opanoidi
OH
Gli opanoidi hanno una struttura che
ricorda quella degli steroli
OH
R1
CH3
H3C
C
CH3
H2C
CH
H2C
Opanoide
(diploptene)
CH3
H
OH
CH3
sterolo
(colesterolo)
a differenza degli eucarioti
i batteri NON hanno steroli nella membrana
R2
Il principale dei fosfolipidi
FOSFATIDIL-SERINA
glicerolo
acidi grassi
O
H2
C
O
C
CH2
CH3
CH2
CH3
O
H
C
O
C
O-
H2
Legami estere
C
O
P
O
O
CH2
CH
NH2
O
Fosfoserina
C
OH
Negli archibatteri
R: Idrocarburi a catena
ramificata
CH3
H2C
C
C
CH2
H
H2
C
O
C
R
H
C
O
C
R
O-H2
C
O
P
O
O
I legami con il glicerolo sono
legami ETERE
due gruppi
(di-eteri)
possono unirsi a
formare tetra-eteri
La lunghezza dei tetraeteri può essere variata
tramite la formazione di anelli pentaciclici
Nelle membrane possono coesistere di- e tetra- eteri
Doppio strato,
flessibile
Monostrato, rigido
La membrana ha diversi ruoli
funzionali
è una barriera di
permeabilità
Verso l’esterno
(enzimi..)
È il luogo dei sistemi
di trasporto
supporta la disposizione
spaziale delle catene
respiratorie
Verso l’interno
(nutrienti)
Conserva l’energia
prodotta dalla cellula
NDH
ATP
FP
FeS
QH2
CITs
CO
Vi si trovano enzimi e trasportatori di biosintesi e sensori per la
percezione dell’ambiente
I batteri fotosintetici hanno sistemi complessi di membrane interne
Nei batteri
fotosintetici verdi
Batterioclorofille e
pigmenti accessori
Si trovano nei clorosomi: strutture allungate connesse
alla membrana da una placca basale
Nei batteri fotosintetici rossi
SULFUREI
vescicolari
esiste un complesso sistema di
membrane interne connesse alla
membrana citoplasmatica
tubulari
Che formano i “cromatofori
Nei batteri fotosintetici rossi NON SULFUREI
Le membrane interne sono
sintetizzate solo in anaerobiosi
Sono lamellari e connesse alla membrana citoplasmatica
CIANOBATTERI
Membrane tilacoidi
Contengono clorofilla e proteine
specifiche per la fotosintesi
Altre membrane interne ad assetto lamellare
si trovano nei batteri nitrificanti
Contengono sistemi di citocromi che agiscono come
sito di ossidazione dell’ammoniaca
Anche i batteri che ossidano il metano hanno membrane interne
simili a quelle dei batteri nitrificanti
perpendicolari all’asse della cellula
(tipo I)
O parallele all’asse della cellula
(tipo II)
LA MEMBRANA CITOPLASMATICA E’ IL LIMITE TRA IL CITOPLASMA E
GLI INVOLUCRI ESTERNI (ENVELOPE)
Nella quasi totalità dei casi, la cellula batterica
è delimitata da una parete caratteristica
delle cellule procariotiche
Che ne determina la forma
e la protegge dalla
lisi osmotica
monodermi
Parete
(peptidoglicano)
periplasma
Membrana cellulare
citoplasma
didermi
PERIPLASMA
Membrana esterna
PARETE
peptidoglicano
Membrana cellulare
citoplasma
Il principale componente della parete è il peptidoglicano (mureina)
formato da due aminozuccheri
Legame β (1-4)
glucosidico
NAM
acido acetilmuramico
(lactil etere di NAG)
NAG
N-acetilglucosamina
D-Glu
DAP(LYS)
D-A
e da pentapeptidi legati a NAM
D-Ala D-Glu e DAP sono aa inusuali
D-A
NAM e NAG formano catene molto lunghe
mediante reazioni di transglicosilazione
FORMANDO UN POLIMERO
PBP: enzimi bifunzionali di membrana provvedono alla transglicosilazione
rimuovono l’ultimo residuo D-Ala
e operano la transpeptidazione
NAG
NAM
NAG
NAM
NAG
NAM
DAP
DAP
DAP
D-A
D-A
D-A
PBP
D-A
D-A
che rafforza il polimero con legami crociati
tra le catene peptidiche
tra la D-ala di una catena e l’acido diaminopimelico (o lisina) di un’altra
I legami crociati sono possibili grazie alla presenza di
COOH
H2N – CH
COOH
H2N – CH
CH2
CH2
CH2
CH2
CH2
CH2
H2N – CH
COOH
LISINA
H2N – CH
H
AC. DIAMINOPIMELICO
questi aminoacidi possiedono due gruppi aminici e possono
formare due legami peptidici
La struttura del peptidoglicano è leggermente diversa
I legami crociati sono formati da ponti peptidici
Monodermi
In genere ponti di penta-glicina
Didermi
I legami crociati sono diretti,
tra un monomero e l’altro
Nei batteri monodermi il peptidoglicano è il
40-80% del peso secco della parete
Altri componenti caratteristici sono gli acidi
teicoici e teicuronici
acidi teicoici : polimeri di glicero o ribitol-fosfato
O
-O
P
O
Legame β 1,6
O C
CH2
COOH
D-Glucosio
D-Alanina
O
O C
O
H
OH HNCOCH3
O C
H
D-Alanina
O C
OH 2C O P O
H
H
H
O
OH
H
O
C
OH
H
H
OH
O
4-N-acetil-D-mannosaminurosil- β-(1- 6)-glucosio
O
Ribitolo
H
acidi teicuronici : polimeri di
subunità ripetitive, di cui una
formata da un acido uronico
MONODERMI
Acidi teicoici
Acidi lipoteicoici
peptidoglicano
I.M.
Porine
(di solito assenti nei monodermi)
VARIAZIONE SUL TEMA
“MONODERMI”: MICOBATTERI
Acil-lipidi
Acidi micolici
arabinogalattano
lipoarabinomannano
peptidoglicano
I.M.
Gli Archibatteri hanno struttura monodermica
e diversi tipi di parete cellulare
Pseudopeptidoglicano
Strato spesso di
Polisaccaridi/polisaccaridi solfati
glicoproteine
proteine
(strato S)
la mancanza di peptidoglicano comporta una mancata tintorialità
con la tecnica di Gram (Gram-negativi)
Ma la presenza di una sola membrana ne fa dei “Monodermi”
come i batteri Gram-positivi
pseudopeptidoglicano
(pseudomureina)
ac. N-acetiltalosaminuronico
NAT
Glu
Legame β (1-3)
glucosidico
N-acetilglucosamina
Stesse struttura e funzione
del peptidoglicano
LYS
Diversa composizione chimica
Glu
Gli aminoacidi che formano i
legami crociati sono “L”
LYS
Glu
NAT
il lisozima- che agisce sui legami
β-1,4 -è inefficace sulla pseudomureina
PRINCIPALI TIPI DI PARETI NEGLI ARCHEOBATTERI
Solo strato S
Strato S e involucro esterno
Solo pseudomureina
Metanocondroitina + strato S basale
Pseudomureina e strato S superficiale
DEINOCOCCO: TRANSIZIONE TRA STRUTTURA
MONO- E DI-DERMICA
(Carboidrati)
Strato S
O.M. non c’è LPS
Strato chiaro
Strato compartimentalizzato
Peptidoglicano
(strato “bucherellato”)
I.M.
la “membrana esterna” si trova solo
nei batteri didermi
fosfolipidi
proteine
Lipoproteine
(OMP)
Lipopolisaccaridi
(LPS)
è più permissiva della
membrana citoplasmatica
Il principale costituente è il lipolisaccaride (LPS)
Catene laterali
specifiche
“Nucleo” polisaccaridico (core)
P
Gruppi fosfato
P
disaccaride
Acidi grassi
Lipide A
La membrana esterna è legata covalentemente al
peptidoglicano dalla “lipoproteina di Braun”
m.e.
peptidoglicano
vi si trovano anche le PORINE, piccole proteine a tre subunità, che formano
canali per il passaggio di soluti idrofili di basso peso molecolare
La membrana esterna e quella interna, nei batteri didermi, delimitano
LO SPAZIO PERIPLASMICO
m.e.
periplasma
m.i.
Dove si trovano enzimi e proteine che non
possono diffondere attraverso le porine
Lo spazio periplasmico è gia “esterno” rispetto
alla cellula didermica
Vi si trovano proteine che svolgono
molte funzioni importanti
Enzimi
idrolitici
Proteine che legano i
soluti da trasportare
nella cellula
Proteine coinvolte nella
chemiotassi
(chemiocettori)
Gli Archibatteri hanno diversi
tipi di parete cellulare
Pseudopeptidoglicano
Strato spesso di
Polisaccaridi/polisaccaridi
solfati
glicoprotein
e
proteine
(strato S)
la mancanza di peptidoglicano comporta una mancata
tintorialità con la tecnica di Gram (Gram-negativi)
Ma la presenza di una sola membrana ne fa dei
“Monodermi” come i batteri Gram-positivi
pseudopeptidoglicano
(pseudomureina)
Legame β (1-3)
glucosidico
ac. N-acetiltalosaminuronico
NAT
Glu
LYS
LYS
Glu
NAT
Stesse struttura e
funzione del
peptidoglicano
Glu
N-acetilglucosamina
Diversa composizione
chimica
Gli aminoacidi che
formano i legami crociati
sono “L”
il lisozima- che agisce sui legami
β-1,4 -è inefficace sulla
pseudomureina







![5\) batteri [modalità compatibilità]](http://s1.studylibit.com/store/data/000928299_1-28a224c15810efb654d8d9e4b207aa31-300x300.png)