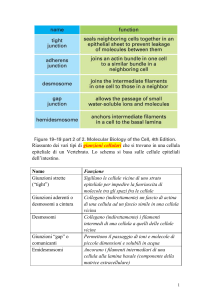
In base alle funzioni che svolgono, le giunzioni cellulari si possono classificare come segue:
Giunzioni occludenti (o tight junctions). La membrana plasmatica di una cellula è in
contatto con la membrana plasmatica della cellula adiacente. Nel punto di contatto delle membrane
delle due cellule, lo spazio extracellulare risulta praticamente assente. Il punto di contatto tra le
membrane è chiamato punto di fusione. Nel punto di fusione, specifiche proteine (es. claudine,
occludine), chiamate proteine di fusione, legano lo strato esterno della membrana plasmatica delle
due cellule adiacenti, creando una barriera impermeabile al passaggio di sostanze. Le proteine di
fusione prendono contatto con alcune proteine citoplasmatiche (proteine della zonula occludens),
che a loro volta si collegano ai microfilamenti di actina del citoscheletro. (FIG.9)
I punti di fusione tra due cellule possono essere numerosi. Le giunzioni occludenti possono
estendersi tutto intorno alla cellula come una cintura che viene chiamata zonula occludens. La
zonula occludens gioca un ruolo fondamentale nella regolazione del passaggio di sostanze da un
versante all’altro (esterno-interno) di un epitelio. La presenza di zonule occludens tra due cellule
adiacenti è particolarmente diffusa nell’epitelio cilindrico semplice che riveste la superficie interna
dell’intestino. Acqua e soluti non sono in grado di passare attraverso le zonule occludens e pertanto
il trasferimento deve avvenire grazie ad un meccanismo di trasporto attivo (→). La presenza delle
zonule occludens limita infatti e di fatto il passaggio per diffusione di sostanze lungo gli interstizi
tra due cellule contigue, creando quindi due ambienti distinti, quello apicale e quello baso-laterale.
Le molecole presenti nel lume intestinale, non potendo diffondersi tra le cellule, vengono assorbite
a livello dell’estremità apicale delle cellule dell’epitelio intestinale grazie alla presenza, nella
regione apicale della membrana plasmatica, di specifiche proteine di trasporto in grado di trasferire
attivamente le molecole dal lume intestinale al citoplasma della cellula e da qui verso la regione
baso-laterale della cellula, quindi per diffusione nel connettivo sottostante l’epitelio, dove vengono
assorbite dai capillari ed inviate nel circolo sanguigno e linfatico. La presenza di zonule occludens
è in grado di creare una differenza funzionale a livello delle cellule epiteliali. Si può verificare che
enzimi presenti sulla membrana cellulare siano di fatto segregati in zone diverse delle cellule
epiteliali per la presenza di zonule occludes tra cellule contigue. Ad esempio, mentre gli enzimi che
operano la digestione terminale di dipeptidi e disaccaridi sono localizzati a livello dei microvilli
della regione apicale delle cellule, l’enzima ATPasi, che trasporta acqua, sali, aminoacidi e
zuccheri, si trova nella porzione basolaterale della cellula, al di sotto della zonula occludens. (FIG.
10)
Giunzioni aderenti e ancoranti. Queste giunzioni sono formate da complessi proteici che
formano un collegamento strutturale tra due cellule adiacenti, estendendosi dal citoplasma di una
cellula al citoplasma della cellula contigua. In pratica, proteine transmembrana di due cellule
adiacenti si collegano tra loro all’esterno della cellula, mentre all’interno di ciascuna cellula si
connettono con proteine citoplasmatiche “di ancoraggio”, a cui a loro volta si legano filamenti del
citoscheletro. I filamenti del citoscheletro possono essere microfilamenti di actina o filamenti
intermedi di cheratina. Si possono definire giunzioni aderenti quelle in cui i filamenti del
citoscheletro sono costituiti da actina. Le proteine che formano le giunzioni aderenti appartengono a
due complessi, uno conosciuto da tempo e formato dalle proteine caderina-catenina, l’altro formato
dalle proteine nectina-afadina. L’organizzazione generale delle giunzioni aderenti, in questi due
casi, è molto simile: in entrambi i casi, infatti, le proteine trans membrana caderina e nectina si
collegano, all’interno della cellula, rispettivamente con proteine della placca di adesione della
famiglia delle catenine (alfa, beta e gamma catenine) e con l’afadina. Le caderine sono le principali
molecole di adesione intercellulare e richiedono la presenza di ioni calcio per potersi legare tra loro.
Fino ad oggi sono state identificate decine di caderine diverse. Le catenine sono proteine di
collegamento che prendono contatto direttamente, o con l’interpolazione di altre proteine di
connessione (es. alfa actinina), con i microfilamenti di actina del citoscheletro. Le giunzioni
aderenti possono estendersi tutto intorno alla cellula come una cintura che viene chiamata zonula
adherens. (Fig . 11)
Nelle giunzioni ancoranti, le proteine trans membrana della famiglia delle caderine (dermocolline e
desmogleine), si collegano all’interno della cellula con proteine della placca di adesione
(desmoplachine, placoglobine, placofiline), che a loro volta si connettono con i filamenti di
cheratina del citoscheletro. I filamenti di cheratina del citoscheletro convergono verso le placche di
adesione, ripiegandosi su se stesse nel punto di contatto con la placca di adesione ed acquisendo
così una forma a forcina. Le giunzioni ancoranti hanno spesso una forma rotondeggiante, da cui il
nome di macula adherens o desmosoma (FIG. 12)
Le giunzioni aderenti o ancoranti sono molto più resistenti alla trazione meccanica delle giunzioni
occludenti. Sono quindi largamente rappresentate, soprattutto come zonula adherens, nelle cellule
epiteliali che rivestono la superficie esterna del corpo e nelle cellule cardiache.
Le giunzioni aderenti e ancoranti sono altresì molto importanti per il mantenimento della coesione
tra l’epitelio ed il tessuto connettivo sottostante. In questa sede, le cellule epiteliali aderiscono alla
membrana basale attraverso un tipo di giunzioni aderenti chiamate adesioni focali ed attraverso un
tipo di giunzioni ancoranti chiamate emidesmosomi.
Le adesioni focali sono complessi proteici che collegano il citoscheletro di una cellula con la
matrice extracellulare (MEC). L’adesione tra la cellula e il MEC è resa possibile dalla presenza di
particolari proteine integrali di membrana, chiamate integrine, che svolgono una doppia funzione,
strutturale e funzionale. La funzione strutturale viene espressa dalla capacità di legare i filamenti di
actina del citoscheletro, attraverso l’interposizione di proteine di collegamento intracellulari come la
vincolina, talina, α-actinina, alle proteine della matrice extracellulare fibronectina, laminine,
collageni, etc. Da un punto di vista funzionale, le integrine sono in grado di trasmettere
informazioni tra l’esterno e l’interno della cellula. Le integrine non sono le uniche proteine cellulari
coinvolte nella formazione delle adesioni focali. Un ruolo analogo a quello delle integrine è svolto
dai sindecani (proteglicani transmembrana). Le adesioni focali sono strutture altamente dinamiche,
in costante rimaneggiamento. A differenza degli emidesmosomi, però, le adesioni focali mediano
l’attacco alla matrice extracellulare di cellule migranti non epiteliali: sono quindi giunzioni
transitorie. Complessivamente le proteine che le costituiscono sono circa un centinaio, suggerendo
quindi una grande varietà di funzioni svolte, appunto, dalle adesioni focali.
L’adesione delle cellule epiteliali alla membrana basale avviene grazie alla presenza di strutture
giunzionali, chiamate emidesmosomi perchè formati da metà desmosoma. Gli emidesmosomi sono
costituiti da una piastra di adesione all’interno della cellula epiteliale a cui si ancorano i filamenti
intermedi di cheratina. L’adesione con la membrana basale è mediata da integrine. (FIG. 13)
Giunzioni comunicanti (o gap junction). Sono giunzioni molto intime, in cui le
membrane di due cellule adiacenti sono in comunicazione grazie alla presenza di proteine canale.
Attraverso tali canali proteici, detti connessioni, si verifica il passaggio di ioni e molecole (delle
dimensioni non superiori ai 1000 dalton) tra due cellule vicine. Le proteine che formano il
connessone prendono il nome di connessine. In genere 6 connessine formano un connessone, che si
unisce ad un connessone presente sulla membrana della cellula adiacente per formare un canale. Le
connessine possono modificare il diametro del canale, che è in grado di aprirsi o chiudersi in
risposta a variazioni dell’ambiente intracellulare. Le giunzioni comunicanti sono presenti anche tra
cellule non epiteliali. Si riscontrano infatti tra le cellule muscolari lisce e cardiache, tra le cellule
nervose, tra i prolungamenti degli osteociti, tra le cellule del Sertoli nei tubuli seminiferi e tra le
cellule follicolari e l’ovocita. (FIG. 14)
Tra due cellule adiacenti possono essere presenti più tipi diversi di giunzioni, che formano i
cosiddetti complessi giunzionali (FIG. 15 complessi giunzionali).