Michele Rismondo
[email protected]
Insegnamento di
BIOLOGIA, ANATOMIA E
MORFOLOGIA VEGETALE
Lezione 04: ANATOMIA E MORFOLOGIA DEL FUSTO
Prima foglia
Gemma
STRUTTURA DI
UNA GIOVANE
PIANTA
(PLANTULA)
EPICOTILE
Cotiledone
IPOCOTILE
Radice
FUSTO O CAULE
E’ la parte del corpo della pianta che deriva dallo sviluppo
dell’apice del germoglio, presente nell’embrione.
Allungandosi dà origine a due tipi di appendici laterali:
RAMI (dai primordi dei rami o gemme laterali)
FOGLIE (dalle tracce fogliari).
FUNZIONI:
1-assicurare il collegamento e il trasporto tra la
radice e le foglie
2 - sostegno
3-da giovane: funzione clorofilliana
4-funzione di riserva (rizomi, bulbi, tuberi)
Il germoglio
Parte aerea della pianta che si origina
dall’embrione. E’ un sistema di organi
costituito da:
•Gemma apicale
•Fusto
•Foglie
•Rami
•Gemme ascellari
bozza fogliare
protoderma
procambio
apice
bozza
fogliare
meristema fondamentale
primordio di ramo
ZONA DI
DETERMINAZIONE
ZONA DI
DIFFERENZIAZIONE
Bozze
fogliari
Primordi
di
rami
APICE
Cordoni procambiali
Bozze
fogliari
Passaggio dalla
struttura embrionale
a quella
adulta primaria
del fusto
ZONA DI
DETERMINAZIONE
Epidermide
ZONA DI
DIFFERENZIAZIONE
Corteccia
Legno
Midollo Procambio
Libro
ZONA DI DETERMINAZIONE
Zona sub-apicale in cui le cellule si organizzano in tessuti
meristematici specifici che poi daranno luogo ai tessuti primari. La
determinazione del protoderma precede quella degli altri
meristemi. In questa fase iniziano a formarsi le appendici laterali del
fusto
PROTODERMA
EPIDERMIDE
PROCAMBIO
SISTEMA CONDUTTORE (conserva a lungo il carattere
meristematico. È il precursore dei fasci cribrovascolari)
MERISTEMA
FONDAMENTALE
PARENCHIMA CORTICALE
(dalla protocorteccia)
PARENCHIMA MIDOLLARE
(dal protomidollo)
ZONA DI DIFFERENZIAZIONE
E’ localizzata alla base della zona di determinazione.
E’ in questa che le cellule si “differenziano” assumendo la loro funzione
definitiva.
Fasci collaterali
aperti
Epidermide
Legno
Midollo
Libro
Procambio
residuale
Corteccia
Schema della STRUTTURA PRIMARIA
Epidermide: deriva dal
protoderma.
Funzione: tegumentale
F
X
Fascio conduttore
Parenchima midollare:
deriva dal meristema
fondamentale.
Funzione: riserva
Cilindro centrale: deriva in parte dal procambio
e in parte dal meristema fondamentale.
Funzioni: conduzione, riserva
Parenchima corticale: deriva dal meristema
fondamentale.
Funzioni: fotosintetica negli strati più
esterni e di riserva, in quelli più interni.
Talvolta anche funzione meccanica
(collenchima).
ZONA DI DIFFERENZIAZIONE
Sviluppo dei FASCI CONDUTTORI
da ciascun cordone procambiale si
formerà un fascio conduttore
formato da xilema e floema.
Per primi si formano protoxilema e
protofloema.
Protoxilema verso interno fusto
(endarco)
Protofloema verso esterno fusto
(esarco)
Gli altri elementi conduttori vengono
formati verso l’interno del fascio
(Metaxilema e Metafloema)
FASCI CRIBROVASCOLARI
F
COLLATERALE CHIUSO (monocotiledoni)
X
F
COLLATERALE APERTO (dicotiledoni e gimnosperme)
C
X
X
CONCENTRICO PERIFLOEMATICO
(rizoma di felce)
F
F
X
CONCENTRICO PERIXILEMATICO
(rizoma delle Monocotiledoni)
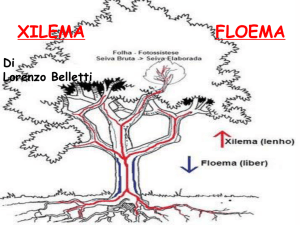
Linfa elaborata
Linfa grezza
TESSUTO VASCOLARE
(XILEMA)
TESSUTO
CRIBROSO
(FLOEMA)
LEGNO - LIBRO
XILEMA - e’ formato da diversi elementi:
tracheidi – Pteridofite e Gimnosperme
trachee - Angiosperme
fibre del legno - sostegno
cellule parenchimatiche - trasporto
•
•
Pteridofite e Gimnosperme: LEGNO OMOXILO (legno secondario formato da sole
tracheidi)
Angiosperme: LEGNO ETEROXILO (legno secondario formato da tracheidi, trachee,
fibre e parenchima del legno.
FLOEMA - e’ formato da diversi elementi:
cellule cribrose - Pteridofite e Gimnosperme
tubi cribrosi con cellule compagne - Angiosperme
fibre floematiche - sostegno
cellule parenchimatiche - trasporto
I fasci cribrosi e vascolari sono ravvicinati a formare un unico fascio
Fascio radiale
Fascio
collaterale
chiuso
Fascio
concentrico
Fascio
collaterale
aperto
Fascio
bicollaterale
FASCI CONDUTTORI
I fasci conduttori possono essere sparsi (Monocotiledoni), oppure
possono formare un anello (Dicotiledoni, Pteridofite, Gimnosperme)
Nelle Gimnosperme e nelle Dicotiledoni il cilindro centrale del fusto viene definito
EUSTELE (=ordinata distribuzione dei fasci cribro-vascolari che sono separati da
raggi midollari costituiti da parenchima interfascicolare.)
Nelle Monocotiledoni il cilindro centrale viene definito ATACTOSTELE (= disordinata
distribuzione dei fasci cribro-vascolari).
EUSTELE
Parenchima corticale
Fasci vascolari
Midollo
Collenchima
Fascio collaterale aperto
con xilema situato verso il centro del fusto e il floema
verso l'esterno, separati dalle cellule del procambio,
un tessuto indifferenziato che andrà a far parte del
cambio
cribrovascolare.
Nella porzione xilematica del fascio si distinguono: il
protoxilema, situato nella parte più interna
(endarco), costituito da piccole tracheidi anulate o
spiralate; e il metaxilema, a contatto con il
procambio, di cui fanno parte grosse trachee.
Nella porzione floematica il protofloema é situato
esternamente rispetto metaxilema. All'esterno del
floema si trova frequentemente un cappuccio di fibre
con
funzione
di
protezione.
Floema
Procambio
Metaxilema in
differenziamento
Metaxilema differenziato
Protoxilema
ATACTOSTELE (fusto di Monocotiledone). Fasci sono sparsi in tutto lo spessore della
stele e sono generalmente più grossi e radi al centro della stele, più piccoli e ravvicinati
alla periferia. In alcune Monocotiledoni i fasci arrivano a ridosso dell'epidermide e
quindi non é possibile distinguere una corteccia.
Parenchima corticale
In alcune Monocotiledoni i fasci arrivano a ridosso dell'epidermide e quindi non é
possibile distinguere una corteccia.
Anello di sclerenchima
Fasci vascolari
Fasci cribrovascolari collaterali chiusi
(non é presente il procambio tra xilema
e floema). Le Monocotiledoni non hanno
accrescimento secondario, e quindi non
si differenzia il cambio cribrovascolare.
Sclerenchima
Protofloema
Metafloema
Tubi cribrosi con
forma poliedrica e
piccole cellule
compagne
Metaxilema
Lacuna protoxilematica
Deriva dal protoxilema lacerato
STRUTTURA SECONDARIA DEL FUSTO
Le piante in base al loro ciclo stagionale di crescita possono essere definite:
•Annuali: l’intero ciclo vitale della pianta (seme - pianta vegetativa - pianta fiorita - seme) si
svolge in un anno
•Biennali: sono necessarie due stagioni per completare il ciclo dalla germinazione alla
formazione del nuovo seme. La prima stagione si conclude con la formazione della radice, di
un corto fusto e della rosetta basale. Nella seconda stagione si ha la crescita, fioritura,
fruttificazione, formazione dei semi e morte della pianta.
•Perenni: le strutture vegetative vivono per vari anni. Le erbacee passano i periodi
sfavorevoli come radici sotterranee dormienti, rizomi, bulbi o tuberi; le legnose
sopravvivono nella stagione avversa con la parte aerea.
Le piante erbacee annuali o biennali hanno un accrescimento secondario scarso o assente.
Le piante perenni, tra cui le Angiosperme dicotiledoni e le Gimnosperme hanno un
accrescimento secondario in spessore che inizia nel loro primo anno di vita. In alcune piante
questo accrescimento dura per molti anni.
CRESCITA SECONDARIA DEL FUSTO
Interessa le Dicotiledoni e le Gimnosperme, manca nelle piante che
vivono una sola stagione (annuali) e nelle Monocotiledoni.
Avviene ad opera di due MERISTEMI LATERALI
CAMBIO
CRIBRO-VASCOLARE
-CAMBIO INTRAFASCICOLARE
(si trova all’interno dei fasci conduttori)
-CAMBIO INTERFASCICOLARE
(si trova a livello dei raggi midollari)
CAMBIO
SUBERO-FELLODERMICO
Si forma nello strato esterno
della corteccia primaria
Il FELLOGENO è il meristema.
sughero
fellogeno
felloderma
PERIDERMA
Al termine dell’accrescimento
primario sono rimaste alcune cellule
di procambio all’interno dei fasci
conduttori.
Iniziano
a
dividersi
prima
le
cellule
parenchimatiche vicino ai fasci, poi quelle
adiacenti fino a completare il cerchio.
Il procambio residuale assume il nome di cambio
intrafasciale e le cellule tra un fascio e l’altro di
cambio interfasciale.
Quando entrambi i cambi sono in divisione
cellulare e si fondono in un unico cilindro
vengono chiamati: cambio cribro-legnoso.
Dal cambio cribro-legnoso si formano:
Xilema secondario (verso l’interno, endarco)
Floema secondario (verso l’esterno esarco)
FORMAZIONE DELL’ANELLO CAMBIALE NEL
FUSTO DELLE DICOTILEDONI
FLOEMA
MIDOLLO
XILEMA
CORTECCIA
CAMBIO
INTRAFASCIALE
CAMBIO
INTRAFASCIALE
CAMBIO
INTERFASCIALE
FLOEMA
FASCI COLLATERALI APERTI
Il cambio cribro-legnoso si
presenta come un mantello conico
interposto tra xilema e floema e si
estende a tutto il corpo
secondario della pianta.
In sezione trasversale il cambio
cribro-legnoso si presenta come un
anello continuo di cellule.
Il cambio cribro-legnoso è un meristema dello spessore di una o due cellule, ma prolifera in due
direzioni: verso l’nterno produce Xilema secondario e verso l’esterno Floema secondario.
In senso stretto si intende soltanto le iniziali cambiali, una per ogni fila radiale di cellule; in senso
più largo si intendono sia le iniziali che le immediatamente derivate perché difficile distinguere
le une dalle altre.
Divisione delle cellule del cambio cribro-legnoso durante una stagione vegetativa.
Dalle cellule iniziali si differenziano cellule del floema secondario e dello xilema
secondario. Vengono prodotte più cellule dello xilema.
Risultato: ispessimento del fusto ed incremento della circonferenza del cambio
cribro-legnoso che migra verso l’esterno.
Il cambio cribro legnoso comprende 2 tipi di cellule:
INIZIALI FUSIFORMI
Cellule allungate secondo l’asse del
fusto, appiattite tangenzialmente e con
estremità appuntite. In sezione
trasversale appaiono appiattite e a
forma di mattone.
Danno origine al sistema assiale o
longitudinale (es. trachee)
Trasporto acqua e minerali in senso
longitudinale
INIZIALI dei RAGGI
Cellule con forma più o meno cubica
isodiametrica orientate
orizzontalmente.
Danno origine al sistema radiale
costituito da cellule allungate
radialmente formanti i raggi midollari
(cellule parenchimatiche dei raggi +
tracheidi dei raggi)
Trasporto acqua e minerali in senso
radiale
Sezioni tangenziali
Cambio cribro-legnoso di melo
(lunghezza iniziali fusiformi circa
0,53mm)
Cambio cribro-legnoso di Robinia
pseudoacacia (stratificato: le iniziali
fusiformi sono disposte in file
orizzontali sulle superfici tangenziali)
CLIMI TROPICALI: in molte specie le cellule
del cambio si dividono in modo più o meno
continuo durante tutto l’anno e gli elementi
dello xilema e del floema si differenziano
gradualmente.
CLIMI TEMPERATI: il cambio rimane a
riposo in inverno e riprende la sua attività
in primavera.
Riattivazione: cellule cambiali assorbono
acqua, si espandono radialmente e si
dividono periclinalmente. Vengono aggiunti
nuovi strati di xilema e floema secondari.
La riattivazione del cambio è stimolata
dall’espansione delle gemme e dalla ripresa
della loro crescita.
tangenziale
radiale
trasversale
Vasi prodotti
in estate
Vasi prodotti
in primavera
Struttura secondaria del fusto di dicotiledone.
Raggi
parenchimatici
Fibrotracheidi
formate in
primavera
Canale
resinifero
Fibrotracheidi
formate in
estate
Sezione trasversale di un legno omoxilo.
La sezione trasversale mostra gli anelli di accrescimento annuale: anelli concentrici di xilema
secondario.
•Nei climi temperati si forma un anello per ogni stagione. Si può determinare l’età della pianta
contando gli anelli (1 anello = 1anno)
•Nei climi equatoriale l’accrescimento avviene durante tutto l’anno e gli anelli sono irregolari
Struttura secondaria del fusto di dicotiledone (Tilia).
Alburno: legno esterno, più chiaro, qui lo xilema è costituito da cellule ancora
attive ed è qui che avviene la maggior parte del trasporto di acqua e minerali in
soluzione.
Duramen: parte del legno più interna e scura; le sue cellule sono spesso piene di
resine che ne ostruiscono il lume; sono cellule per lo più inattive.
Duramen
LEGNO ETEROXILO: legno delle Angiosperme (piante a fiore); è un legno duro come quello
della quercia (Quercus), dell’olmo (Ulmus), del noce (Juglans).
E’ composto da molti tipi di cellule che possono conferire al legno disegni e trame di grande
interesse e valore economico
LEGNO OMOXILO: il legno delle Gimnosperme comprende legni dolci come il pino (Pinus) e
l’abete (Abies); ha una struttura più semplice perché è composto da pochi tipi di cellule per
lo più tracheidi o fibrotracheidi e da raggi midollari molto semplici.
TIPI DI FUSTO
BULBO
Pianta intera con fusto
fortemente raccorciato. La
gemma è circondata da foglie
trasformate in scaglie: interne
succose, esterne secche e
protettive
TUBERO
RIZOMA
Fusto modificato ricco di
riserve
che
cresce
orizzontalmente sottoterra.
Faccia
superiore
con
gemme, faccia inferiore con
radici
Organo sotterraneo con
funzioni di riserva
FUSTI
SOTTERRANEI
MODIFICAZIONI DEL FUSTO
RIZOMI: fusti sotterranei appena sotto la superficie (in alcuni casi fino a
40 cm di profondità).
Possiedono nodi e internodi. In corrispondenza di questi ci sono scaglie, piccole foglie
modificate non fotosintetiche. Le gemme all’ascella delle scaglie producono nuovi rami che
si allungano verso la superficie del terreno formando nuove piantine.
IRIS
RIZOMA
I rizomi sono dei fusti sotterranei che hanno
diverse funzioni: di riserva, di riproduzione
vegetativa e di sopravvivenza; questi organi,
nelle Monocotiledoni, hanno dei fasci
cribrovascolari particolari in cui lo xilema
avvolge il floema formando dei fasci
concentrici perixilematici.
Parenchima aerifero
Fasci perixilematici
Xilema
Floema
MODIFICAZIONI DEL FUSTO
TUBERI: porzioni terminali ingrossate di rizomi sotterranei.
La patata (Solanum tuberosum) ha tre tipi di fusto:
1. verticale con le foglie
2. rizomi sotterranei
3. tuberi, estremità rigonfie dei rizomi.
Gli occhi di un tubero sono le gemme ascellari formantesi all’ascella
di piccole foglie scagliose in corrispondenza di un nodo. Gli internodi
sono corti. Il corpo è pieno di amiloplasti
MODIFICAZIONI DEL FUSTO
BULBO: le sostanze di riserva sono immagazzinate in speciali foglie
carnose (catafilli)
La porzione di fusto è piccola e porta una gemma apicale (che produrrà il fusto erbaceo) e
gemma ascellare che genererà nuovi bulbi.
Le sostanze di riserva immagazzinate nel bulbo sono utilizzate in primavera (es. cipolla,
Allium cepa)
Allium cepa (cipolla)
Leopoldia comosa (lampacione)
Racemo cilindrico
con fiori fertili patenti
all’antesi e fiori sterili
formanti un ciuffo
apicale.
G bulb Euri-Medit.
Comune in
campi,incolti aridi,
margini fino a 1500 m
di quota.
MODIFICAZIONI DEL FUSTO
Il fusto delle Angiosperme (monocotiledoni e dicotiledoni),
può essere modificato per svolgere funzioni diverse da quelle
di sostegno, trasporto ed accrescimento.
Ad esempio può essere un mezzo difensivo o un organo di
adesione (rampicanti), può contribuire alla fotosintesi o
all’accumulo di sostanze di riserva.
FUSTI CON FUNZIONE DI SOSTEGNO (Viticci)
Modificazione del fusto per aggrapparsi ad un
sostegno. Es. Smilax aspera, Vitis vinifera
Smilax aspera
Vitis vinifera
FUSTI ERBACEI
MODIFICAZIONI DEL FUSTO
STOLONI: fusti orizzontali che si formano sopra la superficie del suolo (es.
Fragola, Fragaria vesca).
Ad ogni nodo dello stolone si forma una piccola foglia dalla cui ascella spunteranno una
nuova radice ed una gemma che daranno vita ad una nuova pianta. Gli stoloni facilitano la
diffusione della pianta.
Potentilla reptans
Fragaria
vesca
Cynodon dactilon
RAMI
RAMI MODIFICATI
RAMI MODIFICATI
CLADOFILLI (CLADODI): fusti fotosintetici appiattiti con aspetto e
funzione di foglia.
Non sono foglie perché si sviluppano all’ascella di piccole foglie scagliose. I cladofilli possono
portare fiori, frutti e piccole foglie.
Es. Asparago (Asparagus acutifolius) Pungitopo (Ruscus aculeatus e Ruscus hypoglossum)
Ruscus hypoglossum
Asparagus acutifolius (asparago selvatico)
Specie Stenomediterranea comune
nelle macchie, leccete,
boschi caducifogli, siepi
(0 – 1300 m)
Si distingue da Asparagus tenuifolius per i cladodi aghiformi, induriti e più o meno pungenti e
per i fusti legnosi.
Ruscus aculeatus (pungitopo)
Specie molto comune
presente nei boschi
termofili (0 - 600 m, al
Sud 0 - 1200 m)
I getti giovani sono
commestibili, anche se di
gusto amaro
Le foglie sono sostituite dai
cladodi (assi trasformati)
appiattiti, al centro dei quali
sono inseriti i fiori, piccoli e
dioici.
Ruscus hypoglossum (pungitopo a
foglie larghe)
Specie più mesofila delle precedente,
presente nei boschi freschi di
latifoglie, soprattutto faggete (0 –
1400 m).
RAMI MODIFICATI
SPINE: servono a proteggere la pianta dai predatori
Possono essere modificazioni di fusti (si originano da ascelle fogliari), oppure possono
derivare da trasformazioni fogliari o da emergenze dell’epidermide (es. aculei della
rosa)
Prunus spinosa L.
Aculei di Rosa gallica
Aculei di Rubus idaeus (lampone)
FUSTI CON FUNZIONE DI RISERVA:
Nelle piante adattate agli ambienti aridi sviluppano tessuti particolari detti succulenti in
grado di immagazzinare grandi quantità di acqua. Questi tessuti possono essere
localizzati nei fusti che diventano carnosi e ingrossati.
Fico d’India con rami appiattiti (Pale) con funzione fotosintetica e di accumulo idrico
Michele Rismondo
[email protected]
Insegnamento di
BIOLOGIA, ANATOMIA E
MORFOLOGIA VEGETALE
Approfondimento:
ANATOMIA E MORFOLOGIA DEL FUSTO DI VITIS
PERCHE’ STUDIARE L’ANATOMIA E LA
MORFOLOGIA DELLA VITE ?
E’ la base della
tecnica viticola
ARCHITETTURA DELLA VITE
SCHELETRO:
• FUSTO
• BRANCHE
• TRALCI
• GERMOGLI
ARCHITETTURA DELLA VITE
SCHELETRO
parte lignificata della pianta
FUSTO o CEPPO o TRONCO
l’asse principale della pianta
BRANCHE e BRANCHETTE
assi secondari che partono dal
fusto
SPERONI
rami corti di un anno
TRALCI
rami lunghi di un anno sui quali si sviluppano i GERMOGLI
CHIOMA
Germogli formati da foglie, germogli laterali, grappoli e viticci
ANATOMIA DELLA STRUTTURA PRIMARIA DEL FUSTO
DI VITE
Sezione trasversale
1 – epidermide
2 – parenchima corticale
3 – collenchima
4 – fibre pericicliche
5 – floema
6 – cambio
7 – xilema
8 – raggi midollari
9 - midollo
ANATOMIA DELLA STRUTTURA SECONDARIA DEL
FUSTO DI VITE
Sezione trasversale
1 – epidermide
2 – par. corticale
3 – collenchima
4 – fibre pericicliche
5 – floema duro (formato da fibre
liberiane) e molle (formato da tubi cribrosi
e parenchima ricco di amido e tannino)
6 – cambio
7 – raggio midollare
8 – midollo
9 – xilema
ANATOMIA DI UN TRALCIO MATURO ALLA FINE DEL
PRIMO ANNO
ANATOMIA DELLA STRUTTURA SECONDARIA DEL
FUSTO DI VITE
Sezione
trasversale
CLASSIFICAZIONE E IDENTIFICAZIONE DEI
LEGNI
Caratteristiche per l’identificazione dei legni:
distribuzione dei vasi (visibile in sezione trasversale)
1 - Legni a porosità diffusa
2 - Legni a porosità anulare
Il cambio durante la stagione vegetativa (primavera-estate) produce nuovo legno
che si sovrappone a quello degli anni precedenti.
1 – nei legni a porosità diffusa, il lume dei vasi è pressoché omogeneamente
distribuito su
tutto lo spessore dello strato legnoso annuale. Acero, betulla, ontano, carpino,
faggio, noce,
platano, pioppo, pero, salice, susino, tiglio.
2 – nei legni a porosità anulare, il lume dei vasi è di diversa ampiezza: quelli più
ampi nel legno primaverile o iniziale, quelli più piccoli nel legno estivo o finale.
Castagno, olmo, alcune querce, gelso, bagolaro, robinia, frassino, catalpa, ebano.
CLASSIFICAZIONE E IDENTIFICAZIONE DEI
LEGNI
Dal punto di vista filogenetico, i legni a porosità anulare (propria delle specie di
zone temperate settentrionali) sono più evoluti.
La distribuzione dei vasi è anche influenzata dalle condizioni ambientali e dall’età
della pianta.
La porosità influenza la funzionalità del legno. I legni a porosità anulare hanno
vasi più lunghi e conducono l’acqua con una velocità 10 volte maggiore dei legni
a porosità diffusa. Il legno primaverile è prodotto più rapidamente.
La porosità anulare e diffusa dipende anche dalla velocità di accrescimento delle
nuove foglie e dell’allungamento dei rami: l’accrescimento avviene rapidamente
in quelli a porosità anulare, dura anche d’estate nei legni a porosità diffusa.
CARATTERISTICHE DELLA SEZIONE TRASVERSALE
Cerchi porosi, raramente semi-porosi. Anelli del
legno iniziale discontinui e pori solitari (legno
a porosità anulare). Nel legno finale i pori sono
disposti in file radiali e in piccoli gruppi. Il limite
degli anelli di accrescimento è ondulato, segnato
da una/tre serie di bande di pori appiattiti
radialmente.
Le TILLE sono frequenti nei pori del
legno iniziale a partire dal centro
del
legno.
Parenchima
paratracheale. I raggi midollari
occupano buona parte della sezione
trasversale.
Sezione
trasversale di
un tralcio di vite
CARATTERISTICHE DELLA SEZIONE LONGITUDINALE
TANGENZIALE
Raggi multiseriati, 5-20 serie di raggi. In media i raggi sono alti
più di 2, spesso fino a 5 mm. Frequentemente i raggi hanno
cellule indistinte.
CARATTERISTICHE DELLA SEZIONE LONGITUDINALE RADIALE
Perforazioni semplici piatte nei vasi larghi,
raramente scalariformi, piatte nei vasi del
legno finale.
Raggi omogenei o eterogenei, spesso con
una o due file in quadrati fino alle cellule
marginali diritte. Le fibre liberiane,
occasionalmente settate, sono presenti.
Nel legno finale sono presenti tracheidi,
principalmente
con
punteggiature
scalariformi, ma con numerose forme di
transizione. Nelle cellule radiali sono
presenti i rafidi.