Università degli studi di Roma
“La Sapienza”
Facoltà di Ingegneria delle Telecomunicazioni
“Studio e caratterizzazione del riflesso sudomotorio
mediante imaging termica”
Laureanda
Silvia Carducci
Relatore
Prof. Roberto Cusani
Correlatore
Dr. Arcangelo Merla
1
A tutti coloro che, credendo in me,
mi hanno aiutato a giungere fin qui.
2
Sentiti ringraziamenti al Prof. Roberto Cusani ed al Dr. Arcangelo Merla,
senza l’aiuto dei quali non avrei potuto svolgere questo lavoro al meglio.
3
INDICE
7
Introduzione
Capitolo I
Fisiologia della risposta cutanea galvanica e termica
11
1.1
11
Il sistema nervoso autonomo
1.1.1 Il sistema limbico.
12
1.1.2 Anatomia funzionale del sistema nervoso simpatico.
14
1.1.3 Caratteristiche fondamentali delle funzioni del
15
sistema nervoso simpatico.
1.1.4 Meccanismi di produzione, trasporto e rimozione
16
dei trasmettitori simpatici.
1.1.4.1 Sintesi dell’acetilcolina.
16
1.1.4.2 Sintesi della noradrenalina e dell’adrenalina.
17
1.1.5 La funzione della midollare del surrene.
18
1.2
Anatomia e fisiologia della pelle
23
1.3
Fisiologia del segnale GSR
26
1.3.1
Ghiandole sudoripare e meccanismi base nella
produzione del sudore
27
1.3.1.1
Ghiandole eccrine
27
1.3.1.2
Ghiandole apocrine
28
1.3.1.3
Meccanismi di secrezione del sudore
29
1.3.2
Innervazione delle ghiandole sudoripare
30
1.3.3
Aspetti neurologici della risposta elettrodermica
31
1.3.4
Meccanismi elettro fisiologici delle risposte elettrodermiche
33
1.3.5
Genesi della risposta elettrodermica.
34
1.4
Fisiologia del segnale termico TSR
35
1.4.1 Fisica della dispersione termica a livello cutaneo
37
4
Capitolo II
Materiali e metodi
42
2.1
Descrizione del paradigma sperimentale
42
2.2
Acquisizione del segnale galvanico
45
2.2.1 Strumentazione
2.2.2
46
2.2.1.1 ML116 GSR Amp
46
2.2.1.2
Elettrodi
47
2.2.1.3
unità di controllo Powerlab
48
Metodologia di acquisizione
49
2.2.3 Registrazione
49
2.2.4 Risposta ad uno stimolo emotigeno
50
2.3
50
Acquisizione del segnale termico
2.3.1 La radiazione infrarossa
52
2.3.2 Misure di temperatura
55
2.3.3 L’imaging funzionale infrarosso.
57
2.3.4 Strumentazione.
2.3.4.1 Sensori e Criocooler
58
2.3.4.2 Ottica
61
2.3.4.3 Termocamera ed unità di controllo
61
2.3.4.4 Visualizzazione dei dati infrarosso
62
2.3.5 Gestione software della termocamera: il programma Infra 1.4
64
2.3.6 Calibrazione del corpo nero
64
2.3.7 Standardizzazione delle misure IRFI
65
Capitolo III
Analisi dei dati
3.1
67
Wavelet
3.1.1 Analisi tempo frequenza
67
3.1.2 La Trasformata Wavelet Continua: CWT
72
3.1.3 Dalla CWT discreta alla DWT
78
3.1.4 Analisi multirisoluzione
80
5
3.1.5 Analisi e sintesi
80
3.1.6 Analisi multirisoluzione come implementazione di filtri
81
3.2
Analisi dei segnali
88
3.3
Analisi del GSR
89
3.3.1 Analisi wavelet del GSR
91
3.3.2 Implementazione di una wavelet madre adattata al GSR
95
3.4
Analisi del TSR
113
3.4.1 Utilizzo delle wavelet per l’estrazione della struttura del segnale
114
3.5
Elaborazione dei dati
127
3.6
Modellizzazione del GSR e del TSR
130
Capitolo IV
Risultati
132
Capitolo V
Discussione
139
Conclusioni
143
Bibliografia
144
Ringraziamenti
146
6
Introduzione
Tra i segnali biologici di interesse biomedico comunemente studiati ed utilizzati (ECG,
EEG, EMG, EOG, frequenza respiratoria, etc)(1) ve ne è uno di particolare interesse,
per le possibili e potenziali implicazioni psicometriche ad esso associate,
comunemente noto in letteratura come Galvanic Skin Response (GSR) o risposta
cutanea galvanica. Questo segnale, rilevabile con opportuni sensori posti sul palmo
delle mani o sulle dita, si origina dalla variazione di impedenza cutanea, legata alla
quantità di sudore prodotto dalle ghiandole sudoripare. Poiché, come vedremo, la
sudorazione cutanea rappresenta un indicatore attendibile di possibili stati di stress
mentale ed emozionale del soggetto in esame, la registrazione del GSR viene utilizzata
per monitorare eventuali risposte e reazioni psicologiche, indotte o spontanee, negli
individui. La relativa semplicità del rilevamento del segnale e la presenza nel suo
tracciato di caratteristiche qualitative immediatamente riconducibili agli stimoli hanno
reso il GSR un segnale ampiamente utilizzato in un’ampia varietà di studi e di
condizioni sperimentali.
Una delle applicazioni probabilmente più conosciute, ed al tempo stesso più
controversa, dell’analisi qualitativa del GSR è il suo utilizzo, in associazione con la
registrazione di altri parametri fisiologici, nella macchina della verità. Fra le altre
applicazioni, si ricorda l’utilizzo del GSR negli studi mirati a valutare l’impatto
emozionale di suoni o luci (si veda, ad esempio, “Investigation into the effects of
music and meditation on galvanic skin response”, 2000, N.N. Sudheesh, K.P. Joseph),
il livello di stress individuale (si veda, ad esempio, in relazione all’attività lavorativa,
“Stima dell’ individualità psico- fisiologica di operatori di call center durante l’attività
lavorativa”, 2001, R. Piacentini, P. Sirabella, A. Colosimo, P. Montanari, F. Davide.),
(1) ECG: elettrocardiogramma.
EEG: elettroencefalogramma.
EMG: elettromiografia.
EOG: elettro-oculografia.
7
oppure negli studi sul sonno (“ Skin sympathetic nerve function during sleep – a study
with effector responses”, 2002, R. Kobayashi, Y. Koike, M. Hirayama, H. Ito, G.
Sobue), o in presenza di condizioni patologiche (“ Sympathetic skin response in
patients with spinal cord injury”, 2004, T. Ogura, T. Kubo, K. Lee, Y. Catayama et al.;
“ Evaluating the response of mild Hipertensives to biofeedback- assisted relaxation
using a mental stress test”,1996, E. Paran, M. Amir, N. Yaniv).
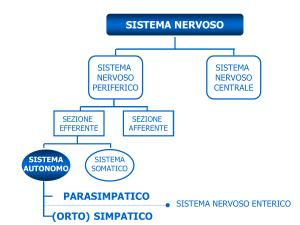
Da un punto di vista fisiologico, il GSR si trova sotto il diretto controllo del sistema
nervoso autonomo (SNA) ed, in particolare, della sua componente simpatica (SNS).
Questo implica che il GSR è un indice diretto dell’attivazione del SNS e che le
informazioni che si ricavano dall’analisi del GSR sono direttamente riferibili
all’attività del sistema nervoso simpatico.
Come verrà meglio specificato in seguito, il SNS è, insieme al Sistema Nervoso
Parasimpatico (SNP), una delle strutture cardine che regolano, a diversi livelli, molti
dei processi regolatori e dei meccanismi di controllo del corpo umano. Tra questi, vi
sono i processi termoregolatori cutanei preposti al controllo dello scambio termico tra
corpo ed ambiente, controllati attraverso un doppio meccanismo: le ghiandole
sudoripare, da una parte, ed il sistema vascolare cutaneo dall’altra. Sulla base delle
precedenti considerazioni, negli ultimi anni, è stato proposto di monitorare lo stato
emozionale individuale ricorrendo alla registrazione della risposta termica cutanea
(Sympathetic Thermal Response o Thermal Skin Response - STR o TSR), tramite
imaging termico(2) , utilizzando termocamere ad infrarossi.
(2)
Merla A, Di Donato L, Rossini PM, Romani GL.(2004)
“Emotion detection through Functional Infrared Imaging: preliminary results”
Biomedizinische Technik,
48(2): 284-286, 2004.
Merla A, Di Donato L, Romani GL, Rossini PM.(2003)
“Recording of the Sympathetic Thermal Response by means of Infrared Fut 2004dddd943( )-3.16695(I)-4.55617(n)-6.16
8
Questa tesi affronta proprio tale problema, sia da un punto di vista qualitativo,
considerando la natura biologica dei due segnali ed il loro legame funzionale, che
quantitativo, attraverso l’analisi dei due segnali, basata sulla decomposizione tramite le
funzioni Wavelets. Lo studio è stato condotto su dati sperimentali raccolti presso il
Laboratorio di Imaging Funzionale Infrarosso e Teletermografia Digitale dell’ITAB –
Istituto Tecnologie Avanzate Biomediche, della Fondazione “Università G.
D’Annunzio” di Chieti.
La scelta di ricorrere alle Wavelets si basa sulla natura non stazionaria dei segnali in
esame, cui un approccio “classico” quale, ad esempio, tramite Trasformata di Fourieur
può risultare insoddisfacente o addirittura inapplicabile. D’altro canto, le Wavelets si
stanno rivelando uno strumento estremamente potente nell’analisi dei segnali alle loro
diverse frequenze e nella ricostruzione nel dominio temporale delle componenti dello
stesso, legate esclusivamente alle frequenze d’interesse. Questa caratteristica consente,
di fatto, di mantenere sia l’ informazione in frequenza che temporale,
simultaneamente.
La tesi si sviluppa in cinque capitoli: il primo affronterà la fisiologia dei due segnali e
sarà orientato ad evidenziare le strutture di controllo del SNS ed i meccanismi
fisiologici ad esso sottoposti, che generano i due segnali in esame; il secondo, i metodi
di registrazione del GSR e del segnale termico ed il protocollo sperimentale utilizzato;
il terzo conterrà inizialmente una breve trattazione sulla teoria delle Wavelets, a cui
seguirà l’ analisi dei dati; il quarto i risultati sperimentali ottenuti; il quinto la
discussione e l’interpretazione dei dati; cui seguiranno le conclusioni.
10
Capitolo primo
Fisiologia della risposta cutanea galvanica e termica
1.1
Sistema nervoso autonomo
Il sistema nervoso autonomo rappresenta la parte del sistema nervoso deputata al
controllo delle attività viscerali dell’organismo tra cui ci sono la pressione arteriosa, la
motilità e l’attività secretoria dell’ apparato gastroenterico, la sudorazione, la
temperatura corporea.
Una caratteristica importante del sistema nervoso autonomo è la rapidità e l’intensità
con cui può modificare la funzionalità viscerale; ad esempio in soli 3-5 secondi può far
aumentare fino a due volte la frequenza cardiaca, in soli 10-15 secondi può far
raddoppiare il livello della pressione arteriosa o, all’ opposto, in 4-5 secondi può
determinarne una tale diminuzione fino a produrre una crisi vagale. Inoltre, in pochi
secondi può indurre sudorazione. Molte di queste modificazioni sono strettamente
correlate allo stato emozionale del soggetto.
Il sistema nervoso autonomo è attivato principalmente da centri situati nel midollo
spinale, nel tronco cerebrale e nell’ ipotalamo. Inoltre alcune porzioni della corteccia
cerebrale, specialmente quella limbica, possono trasmettere impulsi ai centri inferiori
ed in tal modo possono influenzare le caratteristiche del controllo autonomo.
Il sistema nervoso autonomo opera spesso mediante riflessi viscerali, per cui i segnali
afferenti in arrivo ai gangli autonomi, ai centri spinali, tronco encefalici o ipotalamici
trasmettono in via riflessa appropriati comandi al fine di regolarne l’attività. I segnali
afferenti provenienti dal sistema nervoso autonomo sono trasmessi all’organismo
attraverso due principali sezioni, il sistema nervoso simpatico e quello parasimpatico.
11
1.1.1
Il sistema limbico
Un ruolo centrale nel sistema nervoso centrale è giocato dal sistema limbico. Il termine
“limbico” significa “ai confini” e venne originariamente introdotto per designare
alcune strutture nervose localizzate ai bordi delle regioni basali del cervello. Il
significato di questa espressione si è però esteso fino ad indicare l’intero complesso
dei circuiti neuronali che controllano il comportamento emotivo e le pulsioni
motivazionali.
Una parte rilevante del sistema limbico è costituita dall’ipotalamo e dalle strutture ad
esso correlate. Oltre a svolgere un ruolo consistente nel controllo del comportamento,
queste aree regolano anche numerose condizioni interne all’organismo, come la
temperatura, i meccanismi della fame e della sete, il peso corporeo. Queste funzioni
interne vengono designate nel loro complesso come funzioni vegetative ed il loro
controllo è strettamente collegato al comportamento individuale.
Fig. 1.1 : Il sistema limbico (tratto da Guyton & Hall, EDIS, 2005).
La figura 1.1 illustra le componenti anatomiche del sistema limbico, costituito da un
complesso di strutture tra loro strettamente connesse, situate alla base del cervello. Nel
mezzo di queste strutture si trova l’ipotalamo, che, dal punto di vista fisiologico,
12
rappresenta uno degli elementi centrali dell’intero sistema. La figura rappresenta
quindi, schematicamente, la posizione chiave occupata dall’ipotalamo, ed evidenzia
anche le altre componenti sottocorticali del sistema limbico, poste intorno ad esso.
Intorno alle strutture limbiche sottocorticali si trova la corteccia limbica, che funziona
da via di comunicazione bidirezionale e da elemento di associazione tra la
neocorteccia e le strutture limbiche sottocorticali.
Molte delle funzioni comportamentali svolte dall’ipotalamo e da altre strutture
appartenenti al sistema limbico si attuano attraverso la mediazione dei nuclei reticolari
tronco-encefalici e di nuclei ad essi associati.
La stimolazione della porzione eccitatoria della formazione reticolare esalta il grado di
eccitabilità corticale e spinale, infatti, ogni volta che la corteccia cerebrale viene
attivata, sia per effetto di processi mentali che di attività motorie, segnali discendenti
vengono inviati alle aree eccitatorie tronco-encefaliche, le quali a loro volta rinforzano
di rimando il grado di eccitazione corticale. Ciò serve a mantenere o ad esaltare il
livello di eccitazione della corteccia cerebrale, secondo un meccanismo generale di
feedback positivo, per cui una qualunque attività cerebrale una volta iniziata promuove
un ulteriore aumento di attività e di vigilanza mentale.
La maggior parte dei segnali ipotalamici per il controllo del sistema nervoso autonomo
vengono trasmessi attraverso nuclei situati nel tronco encefalico.
13
1.1.2
Anatomia funzionale del sistema nervoso simpatico
Fig. 1.2 : Sistema nervoso simpatico (tratto da Guyton & Hall, EDIS, 2005).
La figura 1.2 mostra l’organizzazione generale della componente periferica del sistema
nervoso simpatico, e mostra una delle due catene di gangli paravertebrali simpatici
presenti ai lati della colonna vertebrale, due gangli (o plessi) prevertebrali (il celiaco e
l’ipogastrico) ed i nervi che collegano i gangli con i vari organi interni. Le fibre
passano dapprima nella catena simpatica e da qui raggiungono i tessuti e gli organi cui
sono destinate. La via simpatica è formata da due neuroni, un neurone pregangliare ed
un neurone postgangliare. Il corpo cellulare del neurone pregangliare si trova nel
midollo spinale ed il suo assone passa dal midollo al nervo spinale corrispondente e da
qui ad uno dei gangli della catena simpatica. I neuroni postgangliari con cui fanno
14
sinapsi i neuroni pregangliari possono trovarsi sia in un ganglio della catena simpatica
che in uno dei gangli prevertebrali.
Particolarmente importanti sono quelle fibre pregangliari simpatiche che, senza dar
luogo a sinapsi, passano dalle cellule del midollo spinale alle catene simpatiche ed ai
nervi splancnici, per raggiungere attraverso di essi la ghiandola midollare del surrene.
Qui le loro terminazioni entrano in contatto diretto con speciali cellule neuronali
modificate, le quali secernono adrenalina e noradrenalina nel sangue circolante. Queste
cellule derivano embriologicamente dal tessuto nervoso e sono assimilabili a neuroni
postgangliari, infatti esse presentano rudimentali prolungamenti nervosi, attraverso i
quali avviene la secrezione di tali ormoni.
1.1.3
Caratteristiche fondamentali delle funzioni del sistema nervoso
simpatico
I neuroni postgangliari simpatici secernono acetilcolina o noradrenalina. Quindi le
terminazioni simpatiche utilizzano come neurotrasmettitori l’acetilcolina o la
noradrenalina. Le fibre che secernono acetilcolina sono dette colinergiche, mentre
quelle che secernono noradrenalina sono dette adrenergiche. Tutti i neuroni
pregangliari sono colinergici, quindi applicando
acetilcolina o altra sostanza
acetilcolinosimile sui gangli del sistema nervoso autonomo tutti i neuroni postgangliari
simpatici vengono eccitati.
La maggior parte dei neuroni postgangliari simpatici sono adrenergici, ad eccezione
delle fibre simpatiche, destinate alle ghiandole sudoripare, ai muscoli piloerettori e ad
alcuni vasi sanguigni, che sono colinergiche.
Questi ormoni agiscono, a loro volta, sui vari organi effettori determinando i rispettivi
effetti simpatici. Spesso la noradrenalina viene definita come trasmettitore (o
mediatore) del sistema simpatico, a causa della predominanza di fibre postgangliari
adrenergiche. Come verrà meglio evidenziato in seguito, le fibre colinergiche dirette
alle ghiandole sudoripare rivestono un ruolo estremamente importante nella genesi del
riflesso sudomotorio.
15
1.1.4
Meccanismi di produzione, trasporto e rimozione dei trasmettitori
simpatici
In questa sezione vengono brevemente descritti i meccanismi di genesi e di rimozione
dei principali trasmettitori simpatici: acetilcolina, adrenalina e noradrenalina.
1.1.4.1
Sintesi dell’acetilcolina
L’ acetilcolina viene prodotta nei terminali colinergici (assoplasma) ed immagazzinata
in alta concentrazioni in vescicole fino a che non viene liberata. La reazione chimica di
base relativa alla sintesi è:
Acetil-CoA + Colina
Colina-acetil-transferasi
Acetilcolina
Fig. 1.3 : Struttura molecolare dell’acetilcolina (tratto da Guyton & Hall, EDIS, 2005).
L’acetilcolina, una volta secreta dalle terminazioni colinergiche, rimane attiva nel
tessuto per alcuni secondi, per essere poi in massima parte scissa in ione acetato e
colina, ad opera dell’enzima acetilcolinesterasi, presente nel tessuto connettivo locale.
La colina così formatasi viene poi trasportata di nuovo all’interno del terminale, dove
viene riutilizzata per sintetizzare nuova acetilcolina.
16
1.1.4.2
Sintesi della noradrenalina e dell’adrenalina
La sintesi della noradrenalina inizia nell’assoplasma della terminazione nervosa delle
fibre adrenergiche, ma si completa all’interno delle vescicole secretorie. I passaggi
fondamentali sono:
1. tirosina
idrossilazione
DOPA
decarbossilazione
2. DOPA
dopamina
3. trasporto della dopamina all’interno delle vescicole
4. dopamina
Idrossilazione
noradrenalina
Nella midollare del surrene questo processo prosegue ulteriormente attraverso la
trasformazione di circa l’80% della noradrenalina in adrenalina:
metilazione
5. noradrenalina
adrenalina
Dopo la liberazione dai terminali nervosi, la noradrenalina viene rimossa dal sito di
secrezione in tre modi diversi:
1. Riassorbimento all’interno della terminazione adrenergica stessa mediante un
sistema di meccanismo di trasporto attivo, responsabile della rimozione del 5080% del trasmettitore secreto.
2. allontanamento per diffusione nei liquidi circostanti e successivamente nel
sangue, modalità responsabile della rimozione della maggior parte della
noradrenalina restante.
3. distruzione enzimatica di una piccola parte.
Solitamente la noradrenalina, liberata direttamente in un tessuto dalle terminazioni
adrenergiche, rimane attiva per pochi secondi, il che sta ad indicare che la sua
rimozione, per riassorbimento e per diffusione, è un evento rapido. La noradrenalina e
17
l’adrenalina secrete nel sangue dalla midollare del surrene, invece, rimangono attive
finché non diffondono in tessuti dove vengono distrutte per via enzimatica, tramite
catecol-O-metiltransegerasi, e ciò avviene specialmente a livello epatico. Di
conseguenza, quando vengono secrete nel sangue, sia la noradrenalina che
l’adrenalina si mantengono molto attive per 10-30 secondi, dopo di che la loro attività
degrada in un tempo compreso tra 1 e pochi minuti.
L’acetilcolina attiva due differenti tipi di recettori, denominati rispettivamente
muscarinici e nicotinici. I recettori muscarinici si trovano in tutte le cellule effettrici
che vengono attivate da neuroni postgangliari colinergici del simpatico. I recettori
nicotinici si trovano, invece, nelle sinapsi tra i neuroni pre e postgangliari del
simpatico.
I principali recettori adrenergici sono i recettori alfa e beta. La noradrenalina eccita
principalmente i recettori alfa, sebbene possa eccitare in minor misura anche i recettori
beta. L’adrenalina, invece, eccita entrambi i tipi di recettori all’incirca nella stessa
misura, quindi gli effetti della noradrenalina e dell’ adrenalina sui diversi organi
effettori dipendono in larga misura dalla tipologia dei recettori presenti negli organi
stessi.
Inoltre, la stimolazione dei nervi simpatici su diverse funzioni viscerali dell’organismo
provoca effetti eccitatori in alcuni organi, ma inibitori in altri.
1.1.5
La funzione della midollare del surrene
La stimolazione delle fibre simpatiche dirette alla midollare del surrene provoca la
liberazione in circolo di grandi quantità di adrenalina e noradrenalina, che con la
circolazione vengono trasportate a tutti i tessuti dell’organismo. Mediamente la
secrezione della midollare del surrene è costituita per circa l’80% da adrenalina e per il
20% da noradrenalina, ma il rapporto tra questi due ormoni può anche variare a
seconda delle condizioni fisiologhe.
Le azioni sui diverse organi dell’ adrenalina e della noradrenalina circolanti sono
pressappoco uguali a quelle prodotte dalla stimolazione simpatica diretta, eccetto che
18
gli effetti si prolungano per un tempo 5-10 volte maggiore, poiché la rimozione di
questi ormoni dal sangue avviene piuttosto lentamente.
La noradrenalina circolante induce costrizione di quasi tutti i vasi
sanguigni
dell’organismo, aumento dell’attività cardiaca, inibizione della motilità e della
secrezione del tubo digerente e dilatazione della pupilla.
L’adrenalina ha effetti pressoché uguali a quelli della noradrenalina, differenziandosi
da essa per i seguenti aspetti. In primo luogo, per via della sua maggiore attività sui
recettori beta, l’ adrenalina esercita sul cuore un’azione più intensa rispetto a quella
della noradrenalina. In secondo luogo, a livello muscolare essa provoca soltanto una
vasocostrizione lieve, se confrontata a quella molto più intensa prodotta dalla
noradrenalina. Dal momento che i vasi muscolari rappresentano la porzione maggiore
dell’intero letto vascolare dell’organismo, questa differenza nell’effetto vasocostrittore
è particolarmente importante, in quanto, se da un lato la noradrenalina induce un forte
aumento globale delle resistenze periferiche e perciò fa innalzare di molto la pressione
arteriosa, dall’altro l’adrenalina ha un effetto ipertensivo minore, ma fa aumentare
assai di più la gittata cardiaca, a causa dei suoi effetti eccitatori sul cuore.
Una terza differenza tra le azioni dell’adrenalina e della noradrenalina riguarda gli
effetti sul metabolismo dei tessuti, che nel caso dell’adrenalina secreta dal surrene
provoca un aumento del metabolismo che può spesso raggiungere il 100%, esaltando
così l’attività e l’eccitabilità di tutto l’organismo.
Riassumendo, la stimolazione della midollare del surrene induce la liberazione di
ormoni che hanno sull’organismo effetti pressoché identici a quelli della stimolazione
simpatica diretta, salvo che essi si prolungano notevolmente (fino 1-2 minuti) dopo la
cessazione dello stimolo.
Le uniche differenze significative sono dovute agli effetti beta dell’adrenalina, la
quale determina aumento del metabolismo e della gittata cardiaca in misura maggiore
di quanto non faccia la stimolazione simpatica diretta, in cui viene rilasciata soltanto
noradrenalina.
La liberazione di adrenalina e noradrenalina da parte della midollare del surrene è
quasi sempre associata anche alla stimolazione diretta dei vari organi per attivazione
generalizzata
del
simpatico.
In
tal
modo,
gli
organi
vengono
stimolati
19
simultaneamente attraverso un doppio meccanismo, e cioè direttamente, ad opera delle
fibre simpatiche ed indirettamente, mediante gli ormoni della midollare surrenale. In
effetti, le due modalità di stimolazione si integrano a vicenda ed in genere l’una può
sostituire l’altra.
Un altro aspetto importante della funzione della midollare surrenale è la capacità di
stimolare, mediante adrenalina e noradrenalina, strutture dell’organismo che non sono
normalmente provviste di innervazione simpatica diretta. Questi ormoni ad esempio,
ed in particolar modo l’adrenalina, sono in grado di stimolare il metabolismo di tutte le
cellule corporee, anche se solo una piccola porzione di esse possiede un’innervazione
simpatica diretta.
Per quanto riguarda le frequenze di attivazione del sistema simpatico una frequenza di
attivazione di un impulso al secondo è in genere sufficiente a mantenere il normale
effetto simpatico ed una completa attivazione può aversi già con frequenze comprese
tra 10-20 impulsi al secondo.
Una caratteristica fondamentale da considerare del sistema simpatico è la sua costante
attivazione, che implica un livello basale continuamente presente noto come tono
simpatico. L’importanza del tono sta nel fatto che esso consente ai diversi settori del
sistema nervoso autonomo di aumentare o ridurre l’attività dell’organo interessato,
semplicemente mediante una modificazione del tono basale. Un esempio molto
importante riguardante l’azione del tono simpatico è il mantenimento di un grado di
vaso costrizione, corrispondente normalmente a circa la metà del loro diametro
massimo, in quasi tutti i vasi sanguigni dell’organismo. Aumentando il grado di
attivazione simpatica, i vasi possono essere ulteriormente costretti, mentre inibendo il
normale tono simpatico essi possono essere dilatati. Se non esistesse un tono simpatico
di base il sistema simpatico potrebbe provocare soltanto vaso costrizione e mai vaso
dilatazione.
Il tono simpatico è condizionato dall’attività secretoria della midollare del surrene.
L’attività secretoria basale della midollare del surrene è normalmente di circa 0.2
µg/minuto per l’adrenalina e di circa 0.05 µg/minuto per la noradrenalina. Tali
quantità sono considerevoli e risultano sufficienti a mantenere una pressione
20
sanguigna di poco inferiore a quella normale, anche dopo rimozione completa
dell’innervazione simpatica diretta di tutto l’apparato cardiovascolare. È evidente,
pertanto, che il tono globale del sistema nervoso simpatico è sostenuto in gran parte
dalla secrezione basale di adrenalina e noradrenalina, ed in grado minore
dall’attività diretta del simpatico.
In molti casi l’attivazione del simpatico causa una “scarica di massa” che può
coinvolgere l’intero sistema come un complesso unitario. Spesso ciò si verifica in
seguito ad attivazione ipotalamica indotta da paura, terrore o dolore intenso. La
risposta è una reazione generalizzata a tutto l’organismo che prende il nome di
reazione di stress o di allarme.
Altre volte l’attivazione interessa settori circoscritti del sistema simpatico e questo
avviene in particolare nel caso di riflessi che coinvolgono il midollo ma non i centri
soprassiali. Esempi molto importanti di questo comportamento sono:
1. il processo di termoregolazione, in cui il sistema simpatico regola la
sudorazione e il flusso ematico cutaneo, senza influenzare altri organi innervati
dal sistema stesso.
2. l’attività muscolare, in cui vengono attivate fibre simpatiche colinergiche
vasodilatatrici, dirette ai muscoli scheletrici.
3. i “riflessi locali”, mediati da fibre sensitive afferenti che, attraverso i nervi
simpatici, raggiungono i gangli ed il midollo spinale e operano su aree
circoscritte. Ad esempio, il riscaldamento di un’area cutanea limitata provoca
vasodilatazione ed intensa sudorazione localmente circoscritte, mentre il
raffreddamento induce localmente effetti opposti.
Nel caso di una “scarica di massa “ l’attivazione simultanea di gran parte delle
componenti del sistema nervoso simpatico potenzia la capacità del corpo di sostenere
un’ attività fisica intensa attraverso vari meccanismi. Questi sono:
1. aumento della pressione arteriosa.
2. aumento del flusso sanguigno a carico dei muscoli in attività e contemporanea
riduzione del flusso a carico di altri organi, quali il tubo digerente, i reni, la
cute, non impegnati nell’ attività fisica.
21
3. aumento del metabolismo cellulare basale dell’intero organismo.
4. aumento della glicemia.
5. aumento della glicolisi muscolare ed epatica.
6. aumento della forza muscolare.
7. aumento dell’attività mentale.
8. aumento della velocità di coagulazione del sangue.
L’interazione di tutti questi effetti permette al soggetto prestazione fisiche molto più
intense e prolungate di quelle che normalmente sarebbe in grado di sostenere. Poiché
di solito è uno stress mentale o fisico quello che eccita il sistema simpatico, si ritiene
che la finalità del simpatico sia quella di provvedere all’ attivazione di emergenza dell’
organismo in condizioni di stress (reazione da stress).
Il sistema simpatico viene attivato intensamente anche da diversi stati emozionali. Ad
esempio nello stato di rabbia o ira, evocato principalmente in seguito ad attivazione di
centri ipotalamici, la trasmissione di segnali attraverso la formazione reticolare troncoencefalica al midollo spinale produce una scarica di massa del sistema simpatico che
produce gli effetti sopra elencati. Come citato, questa reazione è detta reazione di
allarme del simpatico o anche reazione di attacco o fuga, poiché in questo stato il
soggetto decide immediatamente se restare e combattere o scappare. In entrambi i casi
le reazioni simpatiche permettono risposte estremamente rapide e vigorose.
22
1.2
Anatomia e fisiologia della pelle
Sia il GSR che il TSR sono segnali dovuti a variazioni di grandezze nel tempo, legate
alle strutture cutanee. Appare quindi importante richiamare brevemente la descrizione
di tali strutture, in quanto strettamente connesse alla genesi dei segnali considerati.
La pelle è l’organo più esteso del corpo umano e permette di interfacciarsi col mondo
esterno. La sua struttura è in grado di soddisfare a numerose funzioni, quali: la difesa
da agenti esterni ( chimici, meccanici, assalti termici, alcuni tipi di radiazioni ed
infezioni); autoriparazione del tessuto in seguito a ferite; termoregolazione;
regolazione della pressione sanguigna; traspirazione; perspirazione; percezione (tocco
con meccanorecettori, calore con termorecettori, pressione con barorecettori, dolore
con nocirecettori); mantenimento del bilancio complessivo dell’acqua corporea.
Riguardo al mantenimento della temperatura corporea il corpo umano deve poter far
fronte a due diverse situazioni: la prima è un abbassamento della temperatura
dell’ambiente esterno, la seconda un innalzamento della temperatura dell’ambiente
esterno. In entrambi i casi la regolazione dello scambio di calore con l’esterno
coinvolge due meccanismi: la quantità di flusso sanguigno presente nei vasi più
superficiali e la quantità di sudore presente sulla pelle. Infatti, in generale,ad una
maggiore quantità di sangue e di sudore corrisponde una maggiore dissipazione del
calore e viceversa.
Inoltre, la pelle si comporta come una barriera selettiva, che impedisce l’ ingresso di
sostanze esterne e facilita il passaggio di alcune sostanze dal sistema circolatorio
all’esterno.
Le proprietà meccaniche ed elastiche della pelle sono quasi completamente
determinate dalla quantità d’acqua che contiene, di cui una sorgente è il flusso
sanguigno mediante una fitta rete capillare che giunge fino alla superficie del derma.
Lo spessore della pelle varia rispetto alle zone considerate od ai traumi subiti,
comunque in media è dell’ordine di 0.07-0.15mm, fatta eccezione per i palmi delle
mani e le piante dei piedi che raggiungono e possono superare 1mm di spessore. La
pelle si compone di tre strati principali, avente ognuno caratteristiche peculiari:
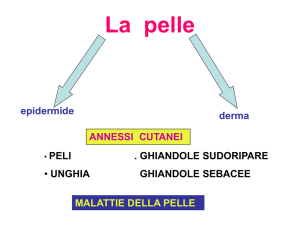
23
epidermide, derma, ipoderma o strato sottocutaneo. I tre strati hanno funzioni, spessore
e resistenza differenti.
Fig. 1.4 : Struttura degli strati cutanei
In particolare:
Epidermide si compone di cinque strati che sono, dall’esterno all’interno:
• Stratum Corneum: svolge un ruolo fondamentale nei fenomeni
elettrodermici quali il GSR ed è composto e continuamente rigenerato
dall’immissione di cheratinociti morti, ossia cellelule cutanee prive di
nucleo ormai essiccate. In genere questo strato è secco, ma diventa
umido in presenza di sudore.
• Stratum Lucidum: è presente solo in zone in cui la pelle è molto spessa.
• Stratum Granulosum: produce cheratina.
• Stratum Spinosum: è composto di cheratociti.
• Stratum Germinativum: è lo strato più profondo dell’epidermide.
24
Derma è lo strato intermedio della pelle. Contiene vasi sanguigni, vasi linfatici,
bulbi piliferi e ghiandole sudoripare. Questo strato è tenuto insieme da una
proteina chiamata collagene, che, inoltre, conferisce resistenza ed elasticità alla
pelle. Nel derma si trovano i recettori specifici per il tatto ed il dolore. Anche il
derma svolge un ruolo importante nei fenomeni legati al GSR.
Ipoderma (o sottocute) è lo strato più profondo della pelle, è formato da una
rete di collagene e cellule grasse. Aiuta nel mantenimento del calore corporeo e
nello stesso tempo costituisce una prima difesa da ferite e traumi, assorbendo
gli urti, per gli organi interni. Il suo ruolo nel GSR è meno importante rispetto
agli altri due strati.
La pelle viene irrorata dai sistemi linfatici e sanguigno; infatti nell’ipoderma si trovano
i vasi da cui si ramificano prima vasi più piccoli, che giungono fino alle ghiandole
sudoripare ed ai bulbi piliferi, e poi una rete di capillari sotto-papillari. La maggior
parte della linfa fluisce attraverso i dotti ipodermici. Il sangue, la linfa, i fluidi
interstiziali contribuiscono attraverso il loro contenuto ionico alla conducibilità
elettrica negli strati interni della pelle.
25
1.3
Fisiologia del segnale GSR
Il termine Galvanic Skin Response (GSR) si riferisce ai cambiamenti nel tempo delle
proprietà elettriche della pelle, che si verificano in risposta a diversi tipi di
stimolazione. Le
proprietà elettriche della pelle si misurano attraverso la sua:
conduttanza (skin conductance: SC), resistenza (skin resistance: SR), potenziale (skin
potential: SP), impedenza (skin impedance: SZ), ammettenza (skin admittance: SY).
Tali cambiamenti possono essere osservati su diverse parti del corpo, ma quelli di
maggiore interesse per fini biomedici si registrano sui palmi delle mani e sono dovuti
quasi esclusivamente alle variazioni del contenuto in ioni dei vari strati della pelle,
causati dalla sudorazione e quindi dipendenti dall’attività delle ghiandole sudoripare.
Poiché l’intero apparato secretorio delle ghiandole esocrine è completamente innervato
dal sistema nervoso autonomo simpatico, dall’analisi dei cambiamenti nel tempo delle
proprietà elettriche della pelle si possono ottenere informazioni dirette sull’attività di
tale sistema.
In letteratura il GSR viene diviso in 2 componenti: un segnale di fondo cha cambia
lentamente nel tempo, detto componente tonica o di livello L, un segnale che cambia
velocemente nel tempo, e che modula il precedente, detto componente fasica o di
risposta R. Entrambe queste componenti sono originate a livello periferico e sono
mediate direttamente dal sistema nervoso centrale (CNS). Esse portano differenti
informazioni. Infatti si suppone che la componente tonica sia legata ad un’attivazione
cerebrale ad ampio spettro, e sia quindi direttamente connessa all’ azione di
noradrenalina ed adrenalina prodotte dalla midollare del surrene, in pratica, essa è
legata a quello che in precedenza è stato indicato come “tono simpatico”. La
componente fasica è invece legata a risposte immediate agli stimoli delle ghiandole
sudoripare e dipende dall’ azione dell’ acetilcolina prodotta e smaltita in loco. Quindi
il sistema simpatico agisce tramite un doppio controllo sulle caratteristiche di questo
segnale. Si suppone inoltre che le due componenti siano legate insieme da alcune
interazioni non lineari a livello periferico. Infatti si ritiene che, anche se l’innervazione
delle ghiandole è colinergica, l’innervazione dei capillari che le irrorarono è
adrenergica, quindi una modificazione nel flusso ematico per vasocostrizione può
influenzare la produzione di sudore.
26
1.3.1
Ghiandole sudoripare e meccanismi base nella produzione del
sudore
Ogni persona possiede in media 2.6 milioni di cellule sudoripare distribuite sull’intero
corpo, fatta eccezione per le labbra, i canali dell’orecchio interno, i capezzoli e gli
organi genitali esterni. La densità media per cm2 è variabile, infatti mediamente in un
adulto si hanno: 233 ghiandole/cm2 sui palmi delle mani, 620 ghiandole/cm2 sulle
piante dei piedi, 360 ghiandole/cm2 sulla fronte, 120 ghiandole/cm2 sulle cosce. Il
numero totale di ghiandole sudoripare non cambia nel corso della vita, ma cambia la
loro densità superficiale man mano che l’individuo cresce.
Le ghiandole sudoripare si dividono in due grandi categorie: apocrine ed eccrine, le
quali differiscono nel modo di secernere il sudore.
1.3.1.1
Ghiandole eccrine
Queste ghiandole si trovano pressoché sull’intera superficie del corpo, in particolare
sui palmi, sulle piante e sulla fronte. Il sudore è prodotto attraverso meccanismi
osmotici che verranno analizzati in dettaglio nella sezione successiva. Dopo la
produzione di sudore il processo secretorio prevede il passaggio del fluido dalle cellule
al dotto, attraverso un membrana che li mette in comunicazione. La produzione di
fluido avviene in modo continuativo, anche se non sempre è associato ad una
espulsione di sudore.
In figura 1.5 è riportato lo schema di una ghiandola sudoripara eccrina.
Strutturalmente le ghiandole sono formate da un tubulo non ramificato, che prima si
avvolge su se stesso e poi prosegue come dotto fino alla superficie dell’epidermide.
Passando dal derma all’epidermide il dotto assume la forma di una bobina a spirale.
La parte secretoria si trova tra l’ipoderma ed il derma. Da questa sezione, il dotto
prosegue attraverso il derma con un percorso ondulato e termina a spirale nell’
epidermide.
27
Fig. 1.5: Ghiandola sudoripara eccrina
Sia la parte secretoria che il dotto sono formati da un doppio strato di cellule: “cellule
basali“(esterne) e “cellule luminali” (interne). Questi strati circondano la parte cava
del dotto che ha un diametro di 5-10 µm. Nella parte finale (epidermica) del dotto c’è
un solo strato. L’intero dotto ed una piccola parte tra il derma e la parte secretoria sono
ricoperti da uno strato di cellule miofiteliali, che ricordano le cellule di muscolo liscio,
e avvolgono il dotto a spirale. Lo strato miofiteliale è molto importante per i
meccanismi che favoriscono il passaggio del sudore nel dotto. È infatti la sua
contrazione a forzare le secrezioni ghiandolari nel dotto.
1.3.1.2
Ghiandole apocrine
Queste ghiandole si trovano in zone localizzate del corpo: sulle ascelle e nell’area anogenitale, quindi non svolgono un ruolo importante nell’attività legata al GSR.
Il processo secretorio prevede una interruzione della secrezione delle cellule ed una
liberazione del loro contenuto nel lumen della ghiandola
attraverso un processo
necrobiotico.
In genere hanno origine in prossimità dei bulbi piliferi.
Le ghiandole apocrine, al contrario di quelle eccrine, non funzionano nei neonati.
Le ghiandole di interesse rispetto alle attività legate al GSR sono quelle eccrine.
28
1.3.1.3
Meccanismi di secrezione del sudore
Quando le ghiandole sudoripare vengono stimolate dalle terminazioni nervose, tutte le
cellule secernono un fluido ottenuto filtrando il plasma, il quale è chiamato secrezione
primaria (o precursore del sudore). Esso è simile al plasma, ma non contiene proteine e
acidi grassi. Infatti contiene prevalentemente acqua e ioni (alta concentrazione di Na+ e
Cl- , bassa concentrazione di K+ ) e risulta ipertonica rispetto al sangue. Questo fluido
contiene approssimativamente: Na+,alla concentrazione di circa 147-151 mM; Cl- , a
circa 123-124 mM; K+, a circa 5 mM; bicarbonato, a 10-15 mM; ed anche anione
lattico a 15-20 mM, come pure piccole quantità di altri ioni, urea e vitamine.
Il precursore del sudore si muove dalla parte secretoria attraverso il dotto e verso la
superficie della pelle sotto l’azione combinata della pressione idrostatica e delle
contrazioni ritmiche, a frequenza di circa 12-21 Hz, dello strato miofiteliale che
circonda il dotto. Queste contrazioni sono indotte dall’azione delle fibre nervose
colinergiche del sistema simpatico.
Quando il fluido raggiunge la parte dermale del dotto, la sua composizione subisce
diverse modificazioni, soprattutto in relazione al tasso di sudorazione. Nel caso di
limitata produzione di sudore ( stato di riposo o temperatura esterna bassa), le cellule
appartenenti allo strato dritto del dotto hanno il tempo di riassorbire la maggior parte
del sodio e del cloro presenti nel fluido, mentre l’acqua è riassorbita per osmosi.
Pertanto questo sudore è significativamente differente da quello della secrezione
primaria. Ciò è confermato dalla concentrazione ionica presente nella secrezione: il
sodio passa da 147-151 mM a 10-140 mM e il cloro da 123-124 mM a 10-30 mM,
mentre la concentrazione del potassio aumenta. Nel caso di elevata produzione di
sudore (temperatura esterna alta, intensa attività fisica o intense emozioni), le cellule
del dotto non hanno il tempo di riassorbire gli ioni della secrezione primaria ed il
sudore raggiunge la superficie della pelle.
Questi meccanismi sono importanti per capire gli aspetti fisiologici delle risposte
elettrodermiche.
29
1.3.2
Innervazioni delle ghiandole sudoripare
Le fibre nervose presenti nella pelle, e che innervano le ghiandole sudoripare,
appartengono al sistema nervoso simpatico.
Le ghiandole sudoripare sono altamente vascolarizzate e densamente innervate da una
fitta rete di terminazioni nervose, che sono sia di tipo colinergico che adrenergico. La
parte secretoria delle ghiandole eccrine è innervata solo dal sistema simpatico
colinergico che raggiunge anche il derma. Ciò significa che le ghiandole eccrine hanno
sinapsi post-ganglioniche colinergiche, hanno cioè l’acetilcolina come trasmettitore
sinaptico. A tal proposito si ricorda che questo trasmettitore è prodotto ed utilizzato in
loco.
Come per l’intero sistema simpatico, anche per la sudorazione non si ipotizza
l’esistenza di una innervazione che ne inibisca la produzione, poiché in assenza di
stimoli per la produzione, il sudore evapora tanto velocemente che un’ ulteriore
inibizione non potrebbe incidere significativamente sulla quantità. Ciò è importante ai
fini della durata del segnale termico associato alla sudorazione.
Poiché la sudorazione si verifica in risposta ad esigenze e stimoli differenti , essa deve
assolvere a funzioni estremamente diverse tra loro, quali la termoregolazione, la
sudorazione “emozionale” ( o riflesso sudomotorio), o ad altro tipo di sudorazione
fisiologica, quale la perspiratio insensibilis necessario al bilancio idrico. Questo porta
ad una variabilità ed ad una specificità delle funzioni e dei comportamenti delle
ghiandole in relazione alla regione considerata. È da sottolineare il fatto che un diverso
comportamento non è legato a differenze strutturali e anatomiche delle ghiandole, ma
solo alla regione di appartenenza.
Le ghiandole eccrine si suddividono in tre gruppi in relazione ai diversi comportamenti
corrispondenti alle regioni anatomiche cui appartengono:
1. Le ghiandole eccrine dei palmi delle mani e delle piante dei piedi. Rispondono
sostanzialmente a stimoli emozionali, quali dolore, paura ed ansia, mediati
direttamente dalle strutture cerebrali coinvolte nel processamento delle
emozioni; oppure a particolari stimolazioni fisiologiche, quali movimenti e
30
respirazione profonda. Si attivano per termoregolazione solo a fronte di un
innalzamento della temperatura esterna superiore ai 30° C.
2. Le ghiandole eccrine del volto, delle ascelle e degli avambracci. Occupano una
posizione intermedia tra la termoregolazione e la sudorazione emozionale,
rispondendo in modo più moderato a stimoli psichici e termici.
3. Le ghiandole eccrine delle altre zone del corpo. Assolvono pressoché
esclusivamente alla funzione di termoregolazione.
Questa distinzione non è, ovviamente, assoluta, in quanto nei fenomeni biologici non
esistono soglie nette di attivazione di un determinato meccanismo.
È importante sottolineare che ai fini delle indagini psicometriche termografiche, il
volto rappresenta un sito di elezione, poiché essendo esposto all’imaging termico, è
registrabile molto più semplicemente delle mani.
1.3.3 Aspetti neurologici della risposta elettrodermica
Il sudore svolge, dunque, un ruolo prevalentemente di termoregolazione, a parte i
palmi, la piante ed il volto. Le ghiandole in queste ultime zone sono attivate in risposta
a stimoli emozionali quali, ad esempio, la paura, il piacere, l’agitazione; a stimoli
fisiologici quali brevi inspirazioni profonde, movimenti e tatto; a stress mentale. Le
ricerche condotte sulle ghiandole sudoripare e la loro innervazione indicano il
coinvolgimento di diversi centri nervosi, localizzati in diversi strati del sistema
nervoso centrale, in parte tra loro indipendenti. Tuttavia il percorso seguito da questi
meccanismi è ancora poco chiaro, infatti molte delle conoscenze acquisite derivano da
studi condotti sugli effetti delle lesioni e delle stimolazioni sugli animali.
Negli ultimi anni, grazie allo sviluppo di tecniche non invasive per l’imaging
cerebrale, si sono potuti studiare anche i centri nervosi umani coinvolti nel GSR.
L’ ipotalamo è l’ area del cervello che regola direttamente la secrezione del sudore;
infatti è il principale centro di termoregolazione, quindi si suppone giochi un ruolo
importante anche nella stimolazione del GSR. Le sue funzioni sono influenzate da
diverse strutture cerebrali, facenti parte del sistema limbico; inoltre, poiché l’ area
31
corticale interagisce con il sistema limbico, anch’essa può influenzare l’ipotalamo e, di
riflesso, il GSR. Questa affermazione è confermata dalla diminuzione o dalla
scomparsa della conduttanza della pelle, in risposta ad uno stimolo emozionale in
soggetti con lesioni laterali-frontali, rispetto a lesioni in altre zone corticali.
Quindi il riflesso sudomotorio sembra rimanere sotto il completo controllo del sistema
limbico e delle strutture cerebrali ad esso associate. Le strutture limbiche
maggiormente coinvolte nel controllo delle funzione ipotalamiche, legate al riflesso
sudomotorio, sono l’amigdala e l’ippocampo.
La figura 1.6 mostra uno schema delle strutture cerebrali coinvolte.
Fig.1.6 : Strutture limbiche coinvolte nel controllo delle funzioni
ipotalamiche relative al riflesso sudomotorio.
In particolare l’amigdala svolge una funzione eccitatoria, mentre l’ippocampo svolge
una funzione inibitoria.
È l’amigdala, in particolare, a giocare un ruolo chiave nella sudorazione in seguito ad
uno stimolo emozionale; questo perché è la struttura cerebrale coinvolta nelle
“memoria” legata ai modelli comportamentali (comportamenti legati alle emozioni,
32
comportamenti sociali, funzioni endocrine e autonome). Una conferma di tale
affermazione è data dagli studi condotti nel 1965 da Bagshaw sulle scimmie a cui era
stata praticata l’ amigdalectomia. L’attivazione del GSR non si esaurisce con l’attività
limbico-ipotalamica, ma si verifica anche in conseguenza di fenomeni quali respiri
profondi e movimenti. Questo suggerisce la presenza di almeno altre due sorgenti: i
gangli promotori basali e il sistema reticolare.
La prima è formata da aree corticali premotrici, le cui fibre per la trasmissione di
impulsi muscolo scheletrici si trovano in connessioni vicine alle fibre che controllano
la secrezione del sudore. Infatti quando queste aree corticali sono naturalmente o
elettricamente stimolate, o rimosse, si può osservare una intensa sudorazione.
La formazione reticolare è una rete di nuclei e fibre nella parte interna del cervello,
svolge un ruolo molto importante in diversi fenomeni, quali la stimolazione della
corteccia atta a mantenere lo stato d’ allerta, il processamento degli stimoli sensoriali,
la regolazione dei riflessi spinali. Questa può sia causare che influenzare e modulare il
GSR.
1.3.4
Meccanismi elettrofisiologici delle risposte elettrodermiche
Le proprietà elettriche sia dell’ intera pelle che di aree localizzate sono determinate da
diversi fattori. Quando si applica una differenza di potenziale alla pelle è possibile
osservare un flusso di corrente che la attraversa; questa è dovuta alla circolazione di
ioni liberi presenti nelle strutture cutanee. Infatti il sangue, il sudore nei dotti e il fluido
interstiziale che hanno una differente conduttività a causa della loro diversa
concentrazione ionica. Il derma e lo strato sottocutaneo, essendo irrorati dal flusso
sanguigno, dal sistema linfatico, dai fluidi interstiziali, si comportano anch’ essi come
buoni conduttori. D’altra parte lo strato corneo mostra una relativa impermeabilità
all’acqua e agisce, quindi, come una barriera per la diffusione degli ioni. Questa
barriera è però attraversata dai dotti delle ghiandole sudoripare, le quali permettono la
diffusione sia del sudore che degli ioni. I cambiamenti lenti dovuti a questo
meccanismo sono stati ipotizzati come i responsabili della “componente tonica” del
segnale elettrodermico. Oltre alle proprietà resistive, la pelle ha anche proprietà
33
capacitive. Esse derivano dall’attività delle membrane cellulari, che possono mostrare
caratteristiche simili ai condensatori. Infatti le membrane possono immagazzinare
energia elettrica e diventare generatori di potenziale. Le membrane che si comportano
come condensatori polarizzati sono presumibilmente localizzate principalmente nella
parte secretoria delle ghiandole sudoripare, nella zona di transizione tra il derma e
l’epidermide e nell’epidermide. In conclusione, la pelle può essere interpretata come
una rete elettrica formata da circuiti RC in serie e parallelo. È’ possibile supporre che
gli elementi capacitivi possano essere caricati molto velocemente, ma una volta
completata la carica non possa scorrere altra corrente e quindi non possa influenzare
ulteriormente la risposta elettrodermica.
1.3.5
Genesi della risposta elettrodermica.
La figura 1.6 mostra la sequenza di eventi che si verificano durante la risposta
elettrodermica nel dotto epidermico di una ghiandola sudoripara.
Fig.1.7 : Sequenza di eventi durante la risposta elettrodermica.
a) all’inizio della risposta, se lo strato superficiale dello strato corneo è ben
idratato, il poro sudoriparo e la parte distale del dotto sudoriparo sono chiusi
sotto l’effetto della pressione esercitata dallo strato corneo circostante (circa
20mmHg). Infatti la conduttanza non aumenta.
34
b) Se il sudore riempie il dotto, si verifica una maggiore conduttività del dotto
stesso e di conseguenza si misura un aumento di conduttanza nello strato corneo
(composto da cellule morte ormai essiccate).
c) Se il tasso di secrezione è abbastanza alto, può generare una pressione
intraduttale maggiore di quella dello strato corneo. In questo caso ci sarà una
apertura del poro del dotto, che viene
attraversato dal sudore. Quando la
porzione terminale del dotto si apre, la conduttanza aumenta nuovamente. Il
sudore sarà spinto fuori attraverso il poro ma continuerà anche a premere
lateralmente nello strato corneo. L’aumento di idratazione (lateralmente nello
strato corneo e assialmente attraverso il dotto) è il fattore più importante che
contribuisce ad un aumento veloce della conduttanza e si suppone essere
responsabile della “componente fasica” del segnale elettrodermico.
d) Quando il sudore lascia il dotto la pressione intraduttale diminuisce, a meno che
il tasso di secrezione sia tale da compensare il volume perduto. Se questa
diventa minore di quella esterna, il poro e la parte terminale del dotto si
chiudono nuovamente, causando una rapida diminuzione della conduttanza.
1.4
Fisiologia del segnale termico
A differenza del GSR, che viene studiato ed utilizzato da diversi anni, e per il quale
esiste anche una modellizzazione matematica della forma d’onda, non esiste un
analogo per la risposta termica simpatica. Ciò dipende probabilmente anche dalla
molteplicità di processi e sorgenti che lo generano e modulano. In questa sezione verrà
descritta
la fisiologia del segnale termico, ovvero le strutture fisiologiche che
determinano, in senso lato, la temperatura cutanea superficiale.
La temperatura corporea si distingue in: temperatura del “nucleo centrale” (o
temperatura interna) e temperatura superficiale della pelle ( o temperatura cutanea). In
quanto omeotermo, l’uomo mantiene la propria temperatura interna stabile entro limiti
relativamente stretti (37 +/- 0.2 °C), anche in presenza di significative variazioni della
temperatura
ambientale.
La
temperatura
cutanea
può
invece
variare
più
35
significativamente in relazione alla temperatura ambientale, all’attività fisica intensa o
in risposta a stimoli emozionali.
Lo scambio termico con l’esterno è un meccanismo che mira a mantenere costante la
temperatura interna, mantenendo in equilibrio il bilancio termico corporeo tra
produzione di calore e perdita di calore. Il calore è uno dei principali prodotti del
metabolismo corporeo ed è proporzionale ad esso. L’entità del metabolismo corporeo è
funzione: del grado di attività metabolica basale di tutte le cellule dell’organismo;
dell’incremento del metabolismo indotto dall’attività muscolare, dalla tiroxina (o altri
ormoni), dall’adrenalina, dalla noradrenalina e dall’attivazione del sistema simpatico.
La maggior parte del calore è prodotto negli organi profondi, in particolare nel fegato,
nel cervello, nel cuore e nei muscoli scheletrici durante l’attività fisica. Il calore viene
poi trasferito dal nucleo centrale alla pelle e da questa disperso nell’ambiente
circostante. L’entità della dispersione dipende da due fattori: la velocità di conduzione
del calore dalle sorgenti corporee fino alla pelle; la velocità di trasferimento del calore
dalla pelle all’ambiente circostante. La conduzione del calore dal nucleo centrale alla
pelle, attraverso uno strato isolante formato da tessuto adiposo sottocutaneo, si realizza
grazie ai vasi sanguigni che lo penetrano e si distribuiscono nella porzione di tessuto a
diretto contatto con la cute.
Fig. 1.8 : Circolazione cutanea (tratto da Guyton & Hall, EDIS, 2005)
Particolarmente importante è il plesso venoso cutaneo il cui flusso può presentare
variazioni estremamente ampie: da appena sopra lo zero fino al 30% della gittata
cardiaca. Infatti, quando il flusso è elevato, il trasferimento di calore si attua molto
36
facilmente; quando il flusso è ridotto la conduzione di calore risulta meno rilevante. È
importante sottolineare che la conduzione aumenta di circa 8 volte dallo stato di
vasocostrizione completa a quello di completa di vasodilatazione.
La conduzione di calore alla cute mediante il sangue è regolata dal grado di
vasocostrizione delle arteriole e delle anastomosi arterovenose, attraverso cui il sangue
affluisce al plesso venoso cutaneo. Il grado di vaso costrizione è controllato quasi
esclusivamente dal sistema nervoso simpatico.
1.4.1
Fisica della dispersione termica a livello cutaneo
Fig. 9 : Meccanismi di termodispersione (tratto da Guyton & Hall, EDIS, 2005)
I meccanismi di dispersione termica a livello cutaneo sono essenzialmente tre:
1. Irraggiamento
Una persona, esposta alla temperatura ambientale di circa 23 °C ed in
condizioni di moderata ventilazione, irraggia circa il 60% del calore
complessivamente eliminato. L’irraggiamento avviene attraverso emissione
infrarossa. La maggior parte dei raggi infrarossi emessi dal corpo umano ha
una lunghezza d’onda compresa tra 3 e 15 µm.
2. Conduzione
37
Di solito il corpo perde solo piccole quantità (circa il 3%) di calore per
conduzione diretta dalla sua superficie ad altri oggetti con cui si trova a
contatto. La perdita di calore per conduzione all’aria costituisce invece circa
il 15% di tali perdite.
3. Evaporazione
Quando evapora acqua dalla superficie corporea, da questa vengono sottratte
0,5 Calorie per ogni grammo di liquido evaporato. Anche in assenza di
sudorazione, il corpo umano cede acqua e liquidi attraverso la perspiratio
insensibilis (perspirazione) dalla superficie cutanea e dai polmoni, in
quantità compresa tra 450 e 600 ml al giorno. A ciò è associata una continua
perdita di calore pari a 12-16 Calorie per ora. La perspirazione non può
essere controllata
ai fini della termoregolazione,
poiché
avviene
indipendentemente dalla temperatura corporea. La perdita di calore per
evaporazione del sudore può, invece, essere controllata mediante la
regolazione della sudorazione da parte del sistema nervoso autonomo. I
meccanismi di controllo delle ghiandole sudoripare da parte del SNA,
effettuato attraverso il sistema nervoso simpatico e i meccanismi di
secrezione del sudore, sono stati ampiamente discussi nella sezione
riguardante il GSR. Le ghiandole sudoripare
possono essere stimolate
dall’adrenalina o dalla noradrenalina circolanti nel sangue, anche in assenza
di specifica innervazione adrenergica in quasi tutto l’organismo. Ciò è
importante durante l’attività fisica, quando questi ormoni vengono secreti
dalla midollare surrenale e l’organismo ha bisogno di eliminare il calore in
eccedenza prodotto dall’attività muscolare.
La temperatura del corpo è regolata quasi esclusivamente da meccanismi nervosi a
feedback, che operano quasi tutti attraverso centri termoregolatori situati
nell’ipotalamo, il quale si trova sotto il controllo del sistema nervoso autonomo.
Il segnale termico utilizzato in questa tesi proviene da alcune regioni del viso. Questa
scelta è dovuta innanzi tutto all’esposizione di tali zone all’imaging termico, alla
distribuzione sul volto delle ghiandole sudoripare ed alla verifica sperimentale della
38
presenza di risposta simpatica termica sul volto stesso. In particolare è stata
considerata la zona frontale. Tale scelta è stata effettuata in base a diverse
considerazioni. In primo luogo è stato osservato che durante la misurazione non si
verificavano fenomeni di saturazione del segnale. In secondo luogo è una zona in cui
artefatti dovuti al movimento sono meno rilevanti rispetto, ad esempio, alla zona orale,
dove si possono verificare eventi che “deformano” la zona in esame, come la chiusura
e l’apertura della bocca. In terzo luogo sono meno consistenti gli artefatti dovuti alla
respirazione, che, causando moti convettivi sul derma, possono alterare la misura.
Fig.1.10: Immagine termica in scala di grigio di uno dei soggetti partecipanti
all’esperimento. La colorbar riporta il range di temperatura usata per la
visualizzazione della distribuzione di temperatura sul volto del soggetto.
Nell’immagine si apprezza la presenza di due grandi vasi superficiali nella
regione frontale.
Nella zona frontale di interesse, il segnale termico è prodotto fondamentalmente dallo
strato cutaneo, dallo strato muscolare sottostante e dai vasi ematici superficiali.
39
Nello strato cutaneo sono presenti le ghiandole sudoripare da una parte e i piccoli vasi
e capillari sanguigni che irrorano le ghiandole, il derma e l’ipoderma dall’altra.
Ghiandole e capillari sono sotto il controllo del sistema simpatico, le prime per via
colinergica, gli altri per via adrenergica.
Le ghiandole sudoripare eccrine in questa zona assolvono a funzioni sia di
termoregolazione che di risposta sudomotoria. Volendo studiare solo l’ ultimo tipo di
comportamento, si dovrà tenere il soggetto in condizioni sperimentali controllate.
Le piccole vene e capillari ivi presenti rispondono anch’esse a stimolazioni
emozionali. L’attivazione del sistema simpatico, in questo caso, produce una
vasocostrizione dovuta alla necessità di preparare il corpo ad un’ intensa attività fisica,
e quindi di deviare un maggior afflusso sanguigno nel sistema muscolo-scheletrico.
La presenza simultanea di risposta sudomotoria e vasocostrizione periferica è alla base
della cosiddetta sudorazione a freddo.
Nello strato muscolare sono presenti muscoli mimici che permettono movimenti
espressivi, come ad esempio il corrugamento della fronte negli stati di attenzione e di
allerta. Quando ciò avviene, si verifica un aumento dell’attività muscolare, che causa
un aumento dell’attività metabolica cellulare, accompagnata da un maggior consumo
della molecola ATP (adenosintrifosfato). Questa molecola contiene legami fosforici ad
alto potere energetico, quando i muscoli si contraggono tali legami vengono scissi per
ricavarne energia. Ogni legame contiene circa 12 Kcal/mole di ATP in condizioni
fisiologiche, ma non più del 30% dell’energia prodotta viene effettivamente utilizzata,
il resto si trasforma in calore che deve essere disperso a livello cutaneo, aumentando la
temperatura locale.
40