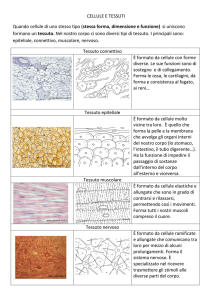
TESSUTI CONNETTIVALI
I tessuti connettivali sono costituiti da cellule polimorfe (autoctone e non
autoctone) e da sostanza intercellulare (componente amorfa o fondamentale
e componente fibrillare).
I tessuti connettivali originano dal mesenchima e si dividono in:
-tessuto connettivo propriamente detto,
-tessuto cartilagineo,
-tessuto osseo,
-sangue e linfa.
I tessuti connettivali svolgono molteplici funzioni:
-funzione trofica (sangue),
-funzione di sostegno (t. osseo, t. cartilagineo, t. connettivo
propriamente detto),
-funzione di difesa (sangue, t. connettivo propriamente detto),
-funzione di protezione (t. connettivo propriamente detto, t.
osseo),
-funzione di connessione (t. connettivo propriamente detto),
-funzione di riserva (t. adiposo, t. osseo).
La
matrice
amorfa
o
sostanza
fondamentale è un sistema colloidale
multifasico formato da una fase
disperdente acquosa nella quale sono
disciolti vari elettroliti, e da una fase
dispersa
costituita
da
enzimi,
glicoproteine
non
strutturali
e
proteoglicani.
La componente fibrillare si divide in:
componente microfibrillare, rappresentata da fibronectina, laminina,
condronectina, con funzione di raccordo ed adesione tra matrice
extracellulare e popolazioni cellulari confinanti,
componente macrofibrillare con funzione strutturale, rappresentata da
fibre collagene, fibre reticolari e fibre elastiche.
Le fibre collagene sono denominate fibre
bianche per il loro aspetto a fresco, possono
presentarsi in forma e dimensioni diverse, sono
molto resistenti alla trazione e ciclicamente
vanno incontro demolizione e rinnovamento.
Le fibre collagene sono costituite da
glicoproteine
fibrose
denominate
tropocollagene o collagene (rappresentano circa
1/3 delle proteine totali di un organismo) lunghe
280-300
nm,
formate
da
3
catene
polipeptidiche , costituite da circa 1000
aminoacidi, avvolte ad elica e stabilizzate da
legami idrogeno, ricche di glicina, prolina,
idrossiprolina, idrossilisina, a cui sono legate
unità saccaridiche costituite da galattosio o
galattosio-glucosio.
Le 3 catene polipeptidiche possono essere uguali o diverse e questo fa sì che esistano
più tipi di collagene (se ne conoscono almeno 14 tipi). Dalla polimerizzazione testa-coda
e latero-laterale di tali catene si formano microfibrille di 20-100 nm che presentano
una periodicità assile di 64-70 nm legata essenzialmente al fatto che la sovrapposizione
laterale è sfalsata di circa ¼ della lunghezza della molecola di tropocollagene. Le
microfibrille si aggregano in fibrille di 0.2-0.3 m dal cui assemblaggio derivano le fibre
collagene definitive.
Le fibre collagene sono prodotte generalmente da
fibroblasti, condroblasti, osteoblasti, attraverso le
seguenti tappe:
-trascrizione e traduzione di un polipeptide precursore
detto protocollagene fornito di peptidi di registro
amino e carbossi terminali uniti da ponti disolfuro.
Questi peptidi di registro hanno la funzione di
impedire la polimerizzazione del protocollagene
all’interno della cellula e di guidare l’allineamento delle
molecole di tropocollagene favorendone l’assemblaggio
in sede extracellulare, dopo la rimozione dei peptidi di
registro,
-idrossilazione dei residui di prolina e di lisina,
-glicosilazione di alcuni residui di idrossilisina e dei
peptidi di registro,
-secrezione del protocollagene all’esterno e sua
trasformazione in tropocollagene per distacco dei
peptidi di registro e successiva polimerizzazione,
-aggregazione
specifica
delle
molecole
tropocollagene per autoassemblaggio spontaneo,
di
-accrescimento delle fibrille di collagene influenzata
probabilmente dalle superficie cellulare della cellula
secernente.
Le fibre reticolari sono costituite da molecole
di tropocollagene che hanno una quota maggiore
di catene glucidiche laterali rispetto a quelle
delle fibre collagene, e che polimerizzano testacoda e latero-lateralmente determinando la
tipica bandeggiatura di 64-70 nm. Le fibrille
che scaturiscono dalla polimerizzazione non
hanno mai dimensioni superiori a 0.2 m, e si
dispongono a formare un reticolo; sono molto
abbondanti nel periodo embrionale, nell’adulto
permangono fibre reticolari a livello delle
membrane basali e dello stroma degli organi
linfoidi ed emopoietici (linfonodi, milza, midollo
osseo rosso) dove sono associate a cellule
reticolari.
Le fibre elastiche sono denominate fibre gialle
per il loro aspetto a fresco quando sono riunite
in grossi fasci in cui hanno andamento parallelo,
possono
presentarsi
anche
isolate
o
anastomizzate a formare reti o lamine. Hanno
dimensioni di 0.2-1 m e sono fortemente
estensibili. Formano alcuni legamenti, entrano
nella costituzione della parete di alcune arterie,
in quella delle cartilagini elastiche e
contribuiscono all’elasticità del polmone.
Le fibre elastiche sono costituite da una
componente amorfa che si forma come
tropoelastina (ricca in lisina) che diviene
elastina quando residui di lisina e di allisina
(aldeide della lisina) reagiscono originando
composti eterociclici chiamati desmosina ed
isodesmosina, e da una componente fibrillare
rappresentata da una glicoproteina, la
fibrillina che si organizza in microfibrille di
10-11 nm coinvolte anche nell’orientamento
della tropoelastina durante la formazione del
polimero di elastina.
Le cellule connettivali sono rappresentate da:
-cellule mesenchimali, cellule reticolari, fibroblasti, macrofagi, periciti, cellule
adipose, mastociti (cellule autoctone del connettivo propriamente detto);
- plasmacellule, cellule pigmentate, leucociti (cellule non autoctone del connettivo
propriamente detto);
-cellule della cartilagine;
-cellule del tessuto osseo;
-cellule del sangue.
Cellule del Tessuto Connettivo propriamente detto
La cellula mesenchimale è un elemento multipotente da cui originano i precursori di
varie cellule del tessuto connettivo. Si presenta stellata o fusiforme, con un grosso
nucleo ricco in eucromatina e numerosi poliribosomi. Non è facilmente identificabile
anche se spesso è localizzata nelle zone perivascolari.
La cellula reticolare è presente nel tessuto mieloide e negli organi linfatici. Ha forma
stellata o fusiforme, nucleo ovale o tondeggiante e in associazione con le fibre
reticolari tende a formare un reticolo cellulare. Produce lo stroma reticolare ed in
alcuni casi può avere attività fagocitaria. Alcuni autori ritengono che cellule reticolari
primitive possano essere elementi staminali dai quali originano elementi del midollo
osseo e macrofagi, tale ipotesi non risulta ancora provata.
I fibroblasti sono le cellule più numerose nei tessuti connettivi propriamente detti. I
fibroblasti giovani hanno forma stellata o fusiforme, con lunghi prolungamenti
citoplasmatici, hanno un nucleo ovale o tondeggiante con un evidente nucleolo,
l’abbondante citoplasma mostra un esteso REG ed un apparato del Golgi ben sviluppato.
I fibroblasti sono responsabili della secrezione del protocollagene e probabilmente
regolano il processo di polimerizzazione e l’orientamento spaziale delle fibre.
I fibrociti rappresentano la forma inattiva dei fibroblasti, il loro nucleo contiene più
eterocromatina, il citoplasma è più scarso ed hanno forma fusata.
I macrofagi o istiociti sono cellule fagocitarie del tessuto connettivo e, dopo i
fibroblasti sono le più rappresentate. Si dividono in macrofagi fissi e migranti. Quelli
fissi sono più numerosi negli spazi pericapillari e lungo le fibre collagene, hanno forma
fusata, nucleo reniforme, un ben evidente apparato del Golgi, abbondante REG e
lisosomi. Per azione di vari stimoli possono diventare migranti ed in tal caso appaiono
tondeggianti e dotati di prolungamenti cellulari.
I macrofagi derivano dai monociti e costituiscono un fattore importante nella difesa
dell’organismo dall’invasione di materiale particolato e nella fagocitosi di detriti
cellulari. Possono anche fondersi, al fine di aumentare la superficie di fagocitosi, in
cellule giganti da corpo estraneo, coinvolte ad esempio nei granulomi.
Anche se qualsiasi cellula che contorna i capillari può essere definita pericito, con
questo termine ci si riferisce più specificamente ad un tipo cellulare in intimo contatto
con l’endotelio dei piccoli vasi tanto che nei punti di contatto è assente la membrana
basale. Alcuni periciti sono anche dotati di attività contrattile attraverso la quale
regolano l’ampiezza del lume vasale.
I mastociti o cellule granulose basofile si trovano per lo più lungo il percorso dei piccoli
vasi riuniti in gruppetti. Volume e forma sono legati alla specie anche se spesso appaiono
ovoidali con un nucleo piccolo, tutti comunque mostrano a livello citoplasmatico la
presenza di granuli basofili contenenti eparina (anticoagulante e chiarificante del sangue)
ed istamina (vasodilatante), in alcune specie è presente anche serotonina. L’origine dei
mastociti non è ancora stata chiarita, alcuni autori li fanno derivare dalle cellule
mesenchimali.
Le cellule adipose o adipociti possono presentarsi isolate, riunite in gruppi o in spessi
strati associati in genere al tessuto connettivo lasso. La loro particolarità porta alla
descrizione di una varietà di tessuto connettivo: il tessuto adiposo.
Le plasmacellule svolgono un ruolo importante nella protezione dell’organismo dagli
antigeni in modo immediato e prolungato sintetizzando e secernendo immunoglobuline.
Derivano dai linfociti.
Le cellule pigmentate possono produrre pigmento (melanociti) o acquisirlo passivamente
(melanofori). Le cellule pigmentate connettivali non sono melanociti, il pigmento in esse
contenuto è stato acquisito passivamente o è stato fagocitato, per cui esse sono
melanofori o cromatofori.
Tessuti Connettivi Embrionali
I tessuti connettivi embrionali sono rappresentati da:
-Mesenchima, è un tessuto mesodermico ricco in cellule
mesenchimali che attraverso i loro prolungamenti costituiscono un
reticolo nel quale si dispone una sostanza fondamentale formata da
glicosaminoglicani e delicate fibre collagene e reticolari. E’ la
matrice embrionale di tutti i tessuti connettivi, il suo
differenziamento porta le cellule mesenchimali a divenire
fibroblasti dai quali deriveranno anche le cellule muscolari lisce.
Nell’adulto in seno ai connettivi si ritrovano piccoli gruppi di cellule
mesenchimali.
-Tessuto connettivo mucoso, E’ uno stadio avanzato del mesenchima,
costituito da cellule mesenchimali, fibroblasti, fibre collagene e
sostanza mucosa. Nell’embrione è presente nel funicolo ombelicale
(gelatina di Wharton) e nelle regioni subepidermiche. Nell’adulto
rappresenta un tessuto transitorio nella riparazione dei tessuti di
sostegno. E’ presente inoltre nella cresta e nei bargigli dei gallinacei
ed in alcune porzioni della lamina propria della mucosa omasale.
Tessuti Connettivi Propriamente Detti
Tessuto connettivo lasso o tessuto connettivo fibrillare lasso:
-è il connettivo più ubiquitario, è composto da un lasso ordinamento di
fibre collagene, elastiche e reticolari orientate a caso. L’elemento
cellulare predominante è il fibroblasto, sono assenti reticolociti. Nella
sostanza fondamentale l’acido ialuronico prevale sui condroitinsolfato e
sul dermatansolfato,
-è localizzato nel tessuto connettivo sottocutaneo, nelle lamine proprie
delle tonache mucose, nelle sottomucose, nel sottomesotelio,
nell’endostio, intorno alle fibre muscolari e a quelle nervose, forma la
pia madre e l’aracnoide, costituisce l’impalcatura connettivale interna
degli organi parenchimatosi.
Tessuto connettivo
irregolare:
denso
o
compatto
-è strettamente correlato al tessuto connettivo
lasso in quanto fibroblasti e fibre collagene ne
costituiscono la principale caratteristica. Le
fibre collagene formano una rete di fasci spessi
ed ondulati, nelle cui maglie sono alloggiate
poche cellule,
-è localizzato nel derma, nelle capsule fibrose
degli organi pieni,
forma periostio e
pericondrio, e la dura madre.
Tessuto connettivo
regolare:
denso
o
compatto
-è caratterizzato dall’orientamento ordinato e
parallelo delle fibre collagene, i fibrociti
sembrano delineare l’estensione dei singoli fasci,
-forma tendini, aponeurosi e legamenti bianchi.
Tessuto elastico:
-sebbene
presenti
come
tipo
cellulare
predominante
i
fibroblasti,
le
fibre
maggiormente
rappresentate
sono
quelle
elastiche più spesse di quelle collagene,
ramificate ed anastomizzate tra loro,
-costituisce il legamento nucale ed altri
legamenti gialli, entra nella costituzione della
parete delle arterie elastiche.
Tessuto connettivo reticolare:
-predomina la presenza di cellule e fibre reticolari,
-costituisce lo stroma di organi linfatici ed
emopoietici ed il connettivo interstiziale di vari
organi.
Tessuto connettivo pigmentato:
-in cui sono presenti cellule contenenti pigmento,
-è presente nell’iride e nella membrana coroidea
e nel connettivo associato a pelle pigmentata.
Tessuto adiposo
è composto da una popolazione omogenea di cellule adipose o adipociti e
rappresenta una fonte di energia prontamente disponibile in quanto accumula grandi
quantità di grassi. Svolge anche un ruolo di protezione da insulti meccanici e di
assorbimento di forze. Morfologicamente si distinguono:
-tessuto
adiposo
bianco
o uniloculare, le cui grandi
cellule
contengono
un’unica
gocciola lipidica,
-tessuto adiposo bruno o multiloculare, le cui
cellule, più piccole delle precedenti, contengono
più gocciole lipidiche, è tipico degli animali
ibernanti che ne traggono calore.
Tessuto Cartilagineo
La cartilagine, in alcuni vertebrati inferiori (ciclostomi, selaci) costituisce lo
scheletro definitivo dell’animale, nei mammiferi, invece, forma il primitivo
abbozzo fetale della maggior parte dello scheletro e, nel corso dello sviluppo pree post-natale, viene sostituita in larga misura dal tessuto osseo. Essa tuttavia
permane, nelle sue varietà, nell’individuo adulto dove trova localizzazione a livello
di:
Superfici Articolari delle ossa.
Dischi intervertebrali.
Coste o parti di esse.
Dischi e menischi intra-articolari.
Sostegno di alcuni organi dell’apparato respiratorio.
Sostegno del padiglione auricolare.
Nel tessuto cartilagineo troviamo elementi cellulari (condroblasti e condrociti) e
sostanza intercellulare (fibrillare ed amorfa)
Fatta eccezione per le superfici articolari e per la fibrocartilagine, la cartilagine
e’ rivestita da un involucro di tessuto connettivo fibroso compatto denominato
pericondrio.
A differenza degli altri tessuti connettivi, la cartilagine e’ sprovvista di nervi e di
vasi sanguigni ed e’ quindi nutrita per diffusione attraverso la sua matrice
gelificata.
Sulla base dell’abbondanza relativa della sostanza amorfa delle fibre che vi sono
incluse e della natura di queste ultime, si distinguono tre tipi di cartilagini: ialina,
elastica e fibrosa o fibrocartilagine.
CARTILAGINE IALINA
Presenta un aspetto translucido ed e’ molto diffusa nel corso dello sviluppo pre- e
post-natale, mentre nell’adulto persiste in alcune parti dello scheletro (coste,
setto nasale, cartilagini articolari, nasali ed alcune laringee, tracheali e
bronchiali).
I condrociti contengono un diplosoma, numerosi mitocondri, REG e ribosomi liberi, esteso
apparato del Golgi costituito di sacculi appiattiti, vescicole e vacuoli. Sono presenti quantità
variabili di particelle di glicogeno e fasci di filamenti intermedi di 10 m. L’estensione del
complesso di Golgi e del REG e la basofilia citoplasmatica aumentano nella cartilagine in
accrescimento o in rigenerazione.
Essi sono inglobati in una abbondante sostanza intercellulare che forma la capsula ed hanno
morfologia e distribuzione differente in seno al tessuto cartilagineo.
Nella parte profonda della cartilagine le cellule sono sferiche e tendenzialmente raggruppate
in numero di 3-5 (gruppi o nidi isogeni) I gruppi isogeni sono formati da elementi che derivano
per mitosi da un’unica cellula.
Nella parte superficiale della cartilagine i condrociti, di forma sferoidale e con asse maggiore
parallelo alla superficie, solo raramente sono raggruppati in nidi, risultano progressivamente
piu’ appiattiti e nella zona immediatamente sub-pericondrale essi hanno un aspetto fibroblastosimile con capsula scarsamente evidente.
La matrice cartilaginea è divisa tra aree territoriali ed aree interterritoriali.
L’area (matrice) territoriale è la regione che circonda ciascun gruppo isogeno e
che contiene concentrazioni più elevate di mucopolisaccaridi e relativamente meno
fibrille collagene (aspetto di alone più intensamente basofilo e metacromatico).
Nell’ambito della matrice territoriale i mucopolisaccaridi sono ancora più
concentrati nella sottile zona di matrice che circonda immediatamente ciascuna
lacuna
cartilaginea
formando
un
anello
intensamente
basofilo
e
metacromatico(capsula).
L’area
(matrice)
interterritoriale
o
interstiziale
è
la
regione interposta tra
i gruppi isogeni o
territori cellulari che
contiene
concentrazioni minori
di
condroitinsolfato
ed
un
numero
maggiore di fibrille
collagene(aspetto
meno
basofilo
o
leggermente
acidofilo).
La matrice o sostanza intercellulare è costituita da:
fibre collagene che non presentano il tipico periodo, sono molto sottili (10-40m)
e delimitano ampi spazi nei quali è compresa la componente amorfa. Il collagene
della cartilagine differisce anche a livello molecolare da quello della maggior parte
dei connettivi in quanto è costituito da tropocollagene formato da 3 catene
identiche 1(II) ed è quindi codificato da un solo gene. Le catene 1(II)
presentano un contenuto relativamente elevato in idrossilina direttamente correlato
con un’altra percentuale di polisaccaridi,
una componente amorfa in cui prevalgono proteoglicani e glicoproteine e nella quale
sono presenti, in quantità minori, lipidi e lipoproteine. Il grado di idratazione,
sempre elevato, varia sensibilmente con l’età.
I glicosaminoglicani presenti nella cartilagine sono:
condroitin-6-solfato (condroitinsolfato C)
condroitin-4-solfato (condroitinsolfato A)
cheratosolfato
acido ialuronico.
Aumenta in % con l’età
Diminuiscono in % con l’età
Parlando di cartilagine ialina va posta l’attenzione su due varietà particolari
rappresentate da cartilagine articolare e cartilagine metafisaria.
Cartilagine Articolare: è priva del pericondrio, ha forma di lamina ed è molto
levigata, per cui consente lo scorrimento delle superfici scheletriche impegnate
nelle diartrosi (articolazioni mobili). I condrociti hanno forma e disposizione
particolari, condizionate dalle sollecitazioni meccaniche. Si possono individuare i
seguenti strati a partire dalla superficie:
Strato tangenziale: condrociti con carenza di organuli,
Strato intermedio: condrociti di forma globosa,
Strato radiale: condrociti disposti linearmente secondo l’asse longitudinale
Strato calcificato: particolarità dello strato radiale nell’adulto.
Nella cartilagine articolare le fibre formano
un intreccio tridimensionale e fungono da
supporto ai proteoglicani ai quali sarebbe
prevalentemente devoluta la capacità di
opporsi alle sollecitazioni meccaniche.
I proteoglicani hanno la capacità di legare
acqua divenendo molecole molto espanse ed
idratate che determinano la reversibilità
delle deformazioni della cartilagine articolare
nei giovani.
dell’osso,
Cartilagine Metafisaria: viene detta anche cartilagine di coniugazione o di
accrescimento, si trova nelle ossa lunghe per tutta la durata
dell’accrescimento in lunghezza ed ha la forma di un disco interposto tra
epifisi e diafisi dell’ osso.
Sulla base della disposizione dei condrociti si possono distinguere le seguenti
zone procedendo dalla epifisi verso la diafisi:
zona delle cellule a riposo,
zona di proliferazione (cartilagine seriata),
zona delle cellule ipertrofiche,
zona della cartilagine in degenerazione.
CARTILAGINE ELASTICA
Nei mammiferi appartengono alla varietà elastica le
cartilagini del padiglione auricolare e del condotto uditivo
esterno, le cartilagini della tuba di Eustachio, alcune
cartilagini laringee.
La cartilagine elastica differisce dalla cartilagine ialina
per:
Elevato numero di fibre elastiche presenti nella matrice
(20-53%);
Basso contenuto in mucopolisaccaridi;
Minore omogeneità della matrice.
CARTILAGINE FIBROSA
La cartilagine fibrosa o fibrocartilagine si riscontra nei dischi
intervertebrali,nei labbri glenoidei, nei menischi, nella sinfisi
pubica e nella zona di inserzione sull’osso di alcuni tendini. La
cartilagine fibrosa è una forma di transizione tra il tessuto
connettivo denso o compatto e la cartilagine.
Le fibre collagene formano fasci grossolani; la componente
amorfa è piuttosto scarsa; i condrociti, poco numerosi, sono
circondati da una capsula basofila e metacromatica.
A differenza degli altri tipi di cartilagine,la fibrosa è priva di
un vero involucro connettivale o pericondrio e può presentare
vasi sanguigni.
CARTILAGINE CELLULARE, TESSUTO CORDOIDE, TESSUTO CONDROIDE
Questi tessuti,per vari aspetti (mancanza di vasi sanguigni,attitudine a
svolgere funzioni meccaniche) sono assimilabili alla cartilagine, ma da essa si
differenziano perché la sostanza intercellulare è molto scarsa.
Tessuto cordoide: (vescicoloso) rappresenta un residuo della notocorda
localizzato nei dischi intervertebrali. In essi si può distinguere una zona
periferica di fibrocartilagine e un disco centrale denominato nucleo polposo
formato da questa varietà di tessuto cartilagineo. La rottura ed erniazione del
nucleo polposo nel canale vertebrale costituisce la cosiddetta ernia del disco.
Cartilagine
cellulare:
varietà
di
cartilagine,generalmente
transitoria,
caratterizzata da scarsa matrice e numerose cellule addensate e
sferiche.Questa cartilagine è definitiva solo in alcune sedi (es. padiglione
auricolare di piccoli roditori).
Tessuto condroide: (pseudocartilagine) ha caratteristiche simili al tessuto
cordoide. E’ poco diffusa e si trova solo nei vertebrati inferiori ed è frequente
negli invertebrati.
MODALITA’ DI ACCRESCIMENTO DELLA CARTILAGINE
A differenza degli altri tessuti connettivi, il tessuto cartilagineo, ad eccezione
del tessuto fibrocartilagineo, è sprovvisto di nervi e di vasi e quindi si nutre
per diffusione in quanto i vasi si arrestano al pericondrio.
L’accrescimento
diversi:
del tessuto
cartilagineo
avviene
secondo
due meccanismi
Accrescimento interstiziale in cui gli elementi già differenziati si dividono
ripetutamente dando origine ad una progenie di cellule che elabora e deposita
nuova sostanza intercellulare. Le cellule figlie derivanti dalla proliferazione di
ogni cellula iniziale formano un clone di elementi accostati tra loro denominati
gruppi isogeni.
Accrescimento per apposizione che consiste nella differenziazione di cellule
periferiche mesenchimali in condroblasti. Le cellule mesenchimali derivano o dal
tessuto mesenchimale circostante il centro cartilagineo o dallo strato
condrogenico del pericondrio. I condroblasti sono cellule di forma
rotondeggiante che presentano un abbondante REG ed uno sviluppato Golgi,
indice della funzione secernente in cui sono coinvolti.
TESSUTO OSSEO
Embriologicamente deriva dal mesenchima, dal mesoderma parassiale e dalle
creste neurali (ossa della testa).
Cellule del tessuto osseo
Cellule osteoprogenitrici: derivano da elementi mesenchimali e manifestano una intensa
attività mitotica. Sono generalmente situate sugli strati superficiali del tessuto osseo e, in
rapporto alle esigenze locali e sistemiche, rimangono quiescenti o si differenziano in
osteoblasti (osteociti e cellule di rivestimento).
Cellule di rivestimento: elementi fusiformi localizzati sulle superfici temporaneamente
inattive del tessuto osseo.
Osteoblasti: cellule piuttosto voluminose che svolgono intensa attività osteogenica in quanto
producono la matrice organica e regolano la deposizione di quella inorganica, mostrano
quindi RER e Golgi ben sviluppati, ed una intensa esocitosi. La loro superficie presenta
tozzi prolungamenti. Gli osteoblasti sono presenti soprattutto in corrispondenza delle
superfici in via di espansione delle ossa e nello strato osteogenico del periostio e
dell’endostio durante tutto il periodo di morfogenesi dell’osso.
Osteociti: sono le principali cellule dell’osso che ha completato il suo sviluppo. Sono
essenzialmente osteoblasti che, dopo aver elaborato la sostanza ossea, rimangono
imprigionati nella matrice calcificata nell’interno di lacune ossee. Secondo alcuni autori,
tuttavia, l’osteocita non è uno stadio differenziativo terminale, in quanto nel processo di
riassorbimento dell’osso può riprendere le funzioni osteoformative assumendo di nuovo la
forma di osteoblasto, oppure ,essere incorporato in un osteoclasto polinucleato.
Gli osteociti presentano prolungamenti citoplasmatici lunghi e ramificati, sono localizzati
all’interno di cavità denominate lacune ossee dalle quali si dipartono i canalicoli che
accolgono i prolungamenti. I canalicoli sono anastomizzati con quelli provenienti dalle lacune
circostanti; alcuni si aprono sulle superfici vascolari del tessuto (canali di Havers e spazi
midollari). I prolungamenti degli osteociti si connettono tra loro per mezzo di gap
junctions. Rispetto agli osteoblasti dai quali derivano,gli osteociti presentano una sensibile
riduzione del RER e Golgi. Partecipano alla osteolisi osteocitica (liberazione dei minerali
dall’osso) finalizzata al mantenimento omeostatico del livello di calcio nel sangue.
Osteoclasti: sono cellule ossee deputate alla degradazione della matrice ossea, fenomeno
denominato osteolisi osteoclastica e finalizzato al rimodellamento dell’osso; hanno notevoli
dimensioni e sono polinucleate. Sono localizzati sulle superfici ossee in via di
riassorbimento e spesso determinano la formazione di cavità di erosione dette fossette di
Howship. Sul versante di erosione presentano un orletto striato, contengono numerosi
lisosomi, mitocondri, polisomi liberi, Golgi multipli, il RER è ridotto. Il riassorbimento della
matrice avviene in seguito al rilascio di acidi organici (citrico, lattico) che determina un
abbassamento del pH ed il conseguente rilascio degli enzimi lisosomiali che vengono
esocitati dalla cellula. Derivano dalla fusione di macrofagi e quindi dai monociti.
La sostanza intercellulare è costituita da una matrice organica e da una inorganica.
La matrice organica rappresenta il 35% della sostanza intercellulare ed è costituita da:
Fibre collagene (in quantità rilevanti e sono responsabili della acidofilia della matrice
ossea e della resistenza dell’osso alla trazione),
Proteoglicani (0,2-1% peso secco),
Glicoproteine (responsabili della Pas positivà della matrice ossea),
Lipidi,
Proteine.
La matrice inorganica rappresenta il 65% della sostanza intercellulare ed è costituita da:
Fosfato di calcio (sotto forma di cristalli di idrossiapatite lunghi 20-40m e spessi 1.53m),
Carbonato di calcio,
Fosfato di magnesio,
Fluoruro di calcio, responsabile della durezza e rigidità dell’osso, può presentarsi sotto
forma di aggregati cristallini rotondeggianti (negli spazi interfibrillari) o formazioni
cristalline allungate (in corrispondenza delle fibre collagene).
La componente fibrillare è rappresentata da fibre collagene.
A seconda che la sostanza fondamentale sia disposta o meno a costituire
lamelle, il tessuto osseo si suddivide in:
tessuto osseo non lamellare,
tessuto osseo lamellare.
TESSUTO OSSEO NON LAMELLARE
Si distingue in:
a fibre intrecciate, costituisce lo scheletro definitivo di vertebrati inferiori e l’osso
primario dei mammiferi. Si tratta di un tessuto attraversato da cavità anastomizzate
(spazi di Havers contenenti vasi, nervi e midollo osseo) delimitati da trabecole nelle quali
fascetti più o meno cospicui di fibre collagene si intersecano. Varietà di questo tipo di
osso sono rappresentate dall’osso follicolinico (tessuto estrogeno-dipendente), e dal
cemento del dente che si differenzia per la mancanza di vasi.
a fibre parallele, è un tessuto caratteristico negli uccelli; raro e transitorio nei
mammiferi. Le fibre collagene, riunite in fasci piuttosto cospicui, presentano andamento
parallelo. Anche in questo caso sono presenti ampie cavità anastomizzate.
TESSUTO OSSEO LAMELLARE
La matrice si dispone in lamelle che hanno orientamento, rapporti e struttura tali da
garantire, nelle diverse parti dello scheletro, la massima resistenza (alla pressione,
trazione e torsione) con il minimo impiego di materiale.
Le lamelle (4.5-11m) si dispongono a formare sistemi particolari (tessuto osseo
compatto), oppure a costituire trabecole che si intrecciano ed anastomizzano (tessuto
osseo spugnoso).
Tessuto osseo spugnoso
Costituisce la maggior parte delle ossa
brevi, delle epifisi delle ossa lunghe e della
diploe delle ossa piatte. E’ formato da
strati di lamelle associate in trabecole più
o meno spesse e variamente anastomizzate
a delimitare spazi intercomunicanti (cavità
midollari contenenti midollo osseo, vasi e
nervi). Cellule di rivestimento si trovano
sulle superfici delle trabecole e tappezzano
le cavità midollari. Le trabecole sono
orientate in modo da meglio sopportare
pressioni e trazione.
Tessuto osseo compatto
Forma i tavolati (superficiale e profondo)
delle ossa piatte, lo strato superficiale
delle ossa brevi, della diafisi e delle epifisi
di quelle lunghe (molto sottile a livello delle
superfici articolari). Oltre alle lacune
ossee ed ai relativi canalicoli questo
tessuto presenta dei canali che nella
diafisi delle ossa lunghe dei mammiferi
decorrono parallelamente all’asse maggiore
dell’osso (canali di Havers), oppure hanno
decorso trasversale (canali di Volkmann).
Sistemi di lamelle:
sistema
concentrico
o
Harves(costituisce l’osteone);
di
sistema delle lamelle circonferenziali o
limitanti (esterno, interno);
sistema interstiziale.
Osteone.
E’ la formazione tipica del tessuto osseo
compatto, ha forma cilindrica ed altezza di
0.9-1.2mm.
E’ attraversato, in lunghezza, dal canale di
Havers (calibro di 20-110m) che contiene
vasi e fibre nervose amieliniche.
Le lamelle dell’osteone si dispongono
concentricamente intorno al canale di
Havers, in numero da 8 a 15 (si può
raggiungere un minimo di 4 ed un massimo
di 24); generalmente circoscrivono il canale
ma possono anche essere incomplete.
La lamella più profonda è quella di più
recente formazione (deposizione).
All’esterno l’osteone è delimitato da una linea frastagliata
detta linea cementante di Ebner, più mineralizzata.
Nell’ambito di ogni lamella,le fibre hanno decorso tra loro
parallelo, ma l’andamento del collagene di una lamella è
ortogonale, o quasi, a quello della lamella vicina.
Gli osteociti, di forma ellissoidale, possono avere sede
intra- o inter-lamellare con asse maggiore parallelo alle
fibre collagene ed asse minore perpendicolare alle lamelle.
I canicoli si dipartono dalle lacune ossee e:
quelli che si spingono fra le lamelle sono pochi e terminano a fondo cieco,
quelli che attraversano le lamelle sono lunghi, ramificati ed anastomizzati con quelli
provenienti da altre lacune.
I canalicoli che si dipartono dalle lacune più profonde terminano nel canale di Havers.
La disposizione di lacune e canalicoli è ampiamente intercomunicante e garantisce da un
lato la possibilità di scambi metabolici (gap junctions) per tutti gli osteociti e dall’altro
rappresenta un fattore limitante lo spessore della parete dell’osteone.
Le lamelle del sistema interstiziale colmano gli spazi interposti tra gli osteoni.
I canali da Volkmann si distinguono da quelli di Havers in quanto non sono delimitati da
lamelle concentriche. Contengono connettivo e vasi che collegano i vasi di calibro
maggiore situati nel canale di Havers con quelli del periostio.
In corrispondenza dell’inserzione di tendini i canali di Havers terminano a fondo cieco.
PERIOSTIO ED ENDOSTIO: i rivestimenti dell’osso
Periostio: lamina di connettivo fibroso denso, strettamente aderente
alla superficie esterna dell’osso, nel quale si possono distinguere:
lo strato più superficiale che è relativamente povero di cellule
(fibroso);
lo strato più profondo che è più ricco di capillari e cellule alcune delle
quali hanno capacità osteoformativa latente o manifesta (osteoblasti).
L’ancoraggio del periostio all’osso è garantito da grossi fasci di fibre
collagene che si spingono in profondità attraversando il sistema
limitante esterno e quello interstiziale più periferico.
Endostio:
sottile lamina di tessuto connettivo lasso che riveste il
canale midollare, i canali di Havers, i canali di Volkmann e contiene
cellule a varia potenzialità evolutiva (osteogenica, ematopoietica).
DENTINA
Rappresenta la varietà filogeneticamente più antica del tessuto osseo ed è
costituita da odontoblasti, cellule specializzate fornite di lunghi
prolungamenti poco ramificati (fibre del Tomes) e da matrice mineralizzata
nella quale sono scavati i canalicoli che accolgono i prolungamenti degli
odontoblasti. La sostanza intercellulare è costituita dal 22% di matrice
organica e dal 78% di matrice inorganica.
Tra la superficie delle fibre di Tomes e la matrice mineralizzata è
presente la guaina di Newmann.
La dentina si differenzia dal tessuto osseo per il fatto che gli odontoblasti
non sono situati all’interno del tessuto osseo stesso ma sulla sua superficie
interna a contatto con la polpa dentaria. Le fibre collagene decorrono
perpendicolarmente ai canalicoli dentali.
SANGUE
Il sangue è un tessuto connettivo che assolve a diverse funzioni:
respiratoria (trasporto di O2 e CO2),
nutritizia (trasporto delle sostanze nutritizie assorbite ),
escretrice (rene, ghiandole sudoripare, polmoni),
termoregolatrice
mantenimento del tasso idrico,
coordinamento e regolazione (ormoni, enzimi),
difesa (aspecifica e specifica),
regolazione dell’emostasi,
mantenimento della pressione osmotica (minerali) ed oncotica (proteine del plasma).
Il sangue è formato da elementi figurati per il 46% (eritrociti 45%, leucociti e piastrine 1%) e
plasma per il 54% (sostanza intercellulare liquida), ha pH 7,4 e densità 1048-1066; scorre in un
sistema di vasi chiusi, spinto dal cuore che è l’organo propulsore.
Il plasma è un fluido leggermente alcalino dal colore giallino, ha peso specifico 1029-1034, viscosità
1.8-2.1, è composto da:
H2O per il 90%,
proteine (albumine, globuline, fibrinogeno) per il 7%,
componenti organici di altra natura (glico e lipo proteine, urea, aminoacidi, glucosio, grassi) per
circa lo 0.1%,
componenti inorganici (Na+, Ca++, K+, Cl-, bicarbonato, fosfati, ferro, iodio) per circa l’1%,
ormoni,
enzimi,
vitamine,
anticorpi.
Gli elementi figurati comprendono eritrociti,
leucociti e piastrine. Si differenziano a
livello degli organi emopoietici (sacco
vitellino, fegato e midollo osseo nell’embrione
e nel feto, e midollo osseo nell’adulto) e
vengono distrutti negli organi emocateretici
(principalmente nella milza).
Eritrociti o globuli rossi o emazie
Trasportano O2 e CO2, nei Mammiferi
hanno forma biconcava atta ad ampliare la
superficie di scambio, mentre negli Uccelli
e nei Pesci hanno forma ellittica e sono
nucleati. Variazioni della forma ricadono
sotto il fenomeno definito poichilocitosi.
Le dimensioni variano da specie a specie,
variazioni delle dimensioni ricadono nel
fenomeno della anisocitosi. Nei Mammiferi
sono privi di nucleo e di altri organuli,
essendo il loro citoplasma costituito per il
96% da emoglobina; fanno eccezione:
i reticolociti, che rappresentano emazie
giovani che per la presenza di mRNA
stabili
continuano
a
sintetizzare
emoglobina,
mostrano
ribosomi
e
mitocondri,
gli .eritrociti con residui nucleari
rappresentati dai Corpi di Jolly o dagli
Anelli di Cabot,
gli eritrociti con residui citoplasmatici
denominati Siderociti in quanto portano
aggregati di ferritina o Eritrociti con
Corpi di Heinz con aggregati di emoglobina
denaturata.
Leucociti o globuli bianchi
Possono avere funzione di difesa o di secrezione. Il loro numero varia da specie a
specie, il loro aumento numerico viene indicato come leucocitosi, mentre la
diminuzione viene indicata come leucopenia. Si differenziano a livello degli organi
emopoietici, possono presentare attività ameboide e sono incapaci di dividersi.
Sono suddivisi in:
Granulociti o leucociti polimorfonucleati con nucleo lobato, e Agranulociti o cellule
mononucleate
I granulociti sono distinti in:
Granulociti neutrofili, rappresentano il 50-70% dei leucociti, presentano un nucleo plurilobato. Le
granulazioni sono rappresentate da granuli A o primari o azzurrofili (contengono perossidasi,
fosfatasi acida, idrolasi acide, collagenasi, elastasi) che si differenziano dalla faccia concava del
Golgi allo stadio di promielocito, da granuli B o secondari (contengono fosfatasi alcalina, lisozima,
fagocitina, catalasi) che si differenziano dalla faccia convessa del Golgi allo stadio di mielocito e
da granuli terziari. Secondo alcuni autori i granuli secondari e terziari devono essere classificati
come granuli specifici. I granulociti neutrofili possiedono uno pseudopodio che permette loro la
diapedesi (movimento tra le cellule endoteliali) attratti per chemiotassi, grazie a questo
intervengono nei processi infiammatori fagocitando batteri e sostanze estranee.
C ane
Gatto
Cavallo
Bovino
Granulociti acidofili o eosinofili, rappresentano il 2-4% dei leucociti, presentano un nucleo con
numero di lobi inferiore a quello dei neutrofili e con cromatina meno condensata. Le granulazioni
sono costituite da una parte amorfa granulare che contiene enzimi lisosomiali e da un cristalloide
contenente proteine basiche. I granulociti acidofili sono dotati di movimento ameboide ed
intervengono nella fagocitosi del complesso antigene-anticorpo, nelle malattie parassitarie e nei
fenomeni allergici.
Granulociti basofili, rappresentano lo 0.5-1% dei leucociti, hanno un nucleo con diverse lobature e
cromatina addensata. Le granulazioni contengono eparina (anticoagulante), istamina
(vasodilatatrice), serotonina (vasocostrittrice) solo nei roditori. Legano sulla superficie cellulare
IgE prodotte dai linfociti B ed in grado di interagire con gli antigeni che ne hanno determinato la
produzione. I granulociti basofili hanno movimento ameboide a scarsa capacità fagocitaria.
C ane
Gatto
Cavallo
Bovino
Gli agranulociti sono distinti in:
Linfociti, sono cellule che si trovano nel sangue, nella linfa e negli organi linfoidi primari (timo,
midollo osseo) e secondari (milza, linfonodi, tonsille palatine, placche di Peyer). Rappresentano il
20-40% dei leucociti. Nel sangue, allo stato inattivo, sono presenti come piccoli linfociti e
mostrano un grosso nucleo con lieve intaccatura e poco citoplasma. Nei tessuti e negli organi
linfoidi, allo stato attivo, hanno dimensioni maggiori, mostrano un nucleo irregolare con evidente
intaccatura; sono distinti in linfociti medi, grandi e linfoblasti. Sono i costituenti principali del
sistema immunitario che è deputato al riconoscimento e alla neutralizzazione di ogni sostanza
estranea (antigene). I linfociti si dividono in:
Linfociti B, localizzati nelle aree B-dipendenti degli organi linfoidi (follicoli della corticale dei
linfonodi, parte centrale della polpa bianca della milza), determinano l’immunità umorale o
anticorpo mediata particolarmente efficace contro le infezioni batteriche,
Linfociti T, localizzati nelle aree T-dipendenti degli organi linfoidi (paracorticale dei linfonodi,
parte periferica della polpa bianca della milza),
C ane
Gatto
Cavallo
Bovino
I linfociti T si distinguono in:
Linfociti con il corpo di Gall, che contengono
lisosomi associati ad una gocciola lipidica,
Linfociti granulari, che contengono lisosomi
sparsi in prossimità dell’apparato di Golgi,
presiedono all’immunità cellulare o cellulo
mediata che comporta una interazione diretta
tra linfocito ed antigene ed è particolarmente
efficace contro funghi, parassiti, cellule
neoplastiche, cellule infettate da virus.
I linfociti T e i linfociti B riconoscono gli
antigeni grazie alla presenza sulla loro
membrana di molecole glicoproteiche che
costituiscono i recettori per gli antigeni.
Nel sangue periferico e negli organi linfoidi
sono presenti cellule NK (natural killer) che
sono morfologicamente simili ai linfociti
granulari e contengono poliperforine, ossia
fattori citotossici la cui azione è diretta
contro cellule neoplastiche, infettate da virus,
opsonizzate (ricoperte da anticorpi) alle quali
determinano lesioni che portano alla lisi
osmotica. Le cellule NK mancano di recettori
per gli antigeni e producono anche fattori di
regolazione emopoietici.
Monociti, rappresentano il 3-8% dei leucociti, sono i precursori dei macrofagi. Hanno forma
variabile da rotonda ad ovoidale con contorni lisci o irregolari per la presenza di corti
pseudopodi che utilizzano per il movimento ameboide. Presentano nucleo reniforme o a ferro
di cavallo con cromatina dispersa. Nel citoplasma sono presenti granuli azzurrofili assimilabili
a lisosomi primari. I monociti maturi vengono immessi nel torrente circolatorio dal midollo
osseo, entro 24-36 h migrano nel tessuto connettivo.
C ane
Gatto
Cavallo
Bovino
Piastrine, sono coinvolte nella coagulazione del sangue (emostasi), in vivo hanno forma ovale o
tondeggiante, stellata nei preparati fissati. Possono presentare una zona centrale o
cromomero ed una periferica o ialomero, sono prive di nucleo in quanto rappresentano
porzioni citoplasmatiche di precursori chiamati megacariociti. La loro membrana presenta un
rivestimento di mucopolisaccaridi e lipoproteine coinvolti nella coagulazione, nel citoplasma
sono contenuti granuli denominati (fibrinogeno, fattori piastrinici) e lisosomiali (idrolasi
acide).
Linfa
E’ un liquido alcalino trasparente e leggermente giallognolo, che circola nei vasi
linfatici. Si genera per confluenza del liquido intercellulare, alla periferia degli
organi e dei tessuti, in capillari linfatici che originano a fondo cieco. I vasi linfatici
di calibro maggiore che ne derivano giungono alle stazioni linfonodali e da qui la linfa
confluisce nel torrente circolatorio venoso. E’ formata da:
porzione liquida, contente piccole quantità di sali, colesterolo, lecitina, CO2, O2 in
piccole quantità, fibrinogeno, numerosi prodotti del ricambio dei tessuti,
parte corpuscolata, rappresentata quasi esclusivamente da linfociti.
EMOPOIESI
L’emopoiesi è il processo che porta alla formazione degli elementi figurati del sangue.
Per spiegare tale processo sono state avanzate diverse teorie.
Teoria unitaria o monofiletica: una cellula mesenchimale denominata emocitoblasto,
morfologicamente simile ad un medio o grande linfocito, ha la capacità di riconoscere
gli organi emopoietici e di popolarli specificamente, dando origine a tutti gli elementi
del sangue.
Teoria dualistica, le cellule del sangue derivano da due specifici capostipiti:
una cellula capostipite per la serie bianca e per le piastrine
una cellula capostipite per la serie rossa
oppure
un mieloblasto da cui derivano granulociti, monociti, eritrociti e piastrine
un linfoblasto da cui derivano i linfociti
Teoria pluralistica o polifiletica: si hanno tante cellule staminali quanti sono gli
elementi del sangue.
Secondo la teoria più accreditata, durante l’emopoiesi, l’emocitoblasto subisce
trasformazioni graduali che danno origine alla formazione di cellule che evolvono
gradualmente negli elementi differenziati del sangue.