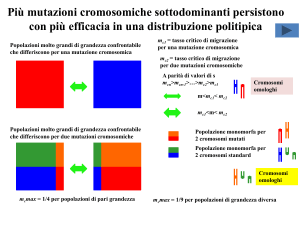
Più mutazioni cromosomiche sottodominanti persistono
con più efficacia in una distribuzione politipica
Popolazioni molto grandi di grandezza confrontabile
che differiscono per una mutazione cromosomica
mc1 = tasso critico di migrazione
per una mutazione cromosomica
mc2 = tasso critico di migrazione
per due mutazioni cromosomiche
A parità di valori di s
mcn>mcn-1>…>mc2>mc1
Cromosomi
omologhi
m<mc1< mc2
mc1<m< mc2
Popolazioni molto grandi di grandezza confrontabile
che differiscono per due mutazioni cromosomiche
Popolazione monomorfa per
2 cromosomi mutati
Popolazione monomorfa per
2 cromosomi standard
Cromosomi
omologhi
mcmax = 1/4 per popolazioni di pari grandezza
mcmax = 1/9 per popolazioni di grandezza diversa
L’espansione di mutazioni cromosomiche
sottodominanti in metapopolazioni
L’espansione casuale del
cromosoma mutato può
avvenire con processi di
estinzionericolonizzazione dei
demi (in rosso)
La metapopolazione in cui i demi
monomorfi per il cromosoma mutato
(rossi) sono fra loro adiacenti presentano
meno flussi migratori (arancio) che
possono destabilizzare i demi…
… rispetto a quelle in cui questi demi
sono dispersi perché sono meno
numerosi, quindi sono più stabili di
queste ultime.
Per questo si realizza la tendenziale
coalescenza degli areali occupati dai demi
monomorfi per il cromosoma mutato.
Accumulo di più mutazioni cromosomiche sottodominanti
mc2 = tasso critico di migrazione
mc1 = tasso critico di migrazione
per due mutazioni cromosomiche
per una mutazione cromosomica
Metapopolazioni monomorfe per un cromosoma presente solo nella sua forma standard (azzurro) ma politipiche per
un altro cromosoma, con alcuni demi monomorfi per la forma standard (verde), altri per la forma mutata (arancio).
Nella metapopolazione a
sinistra un deme
monomorfo per il
cromosoma standard verde
diviene monomorfo per il
cromosoma mutato rosso.
Nella metapopolazione a
destra un deme monomorfo
per il cromosoma mutato
arancio diviene monomorfo
per il cromosoma mutato
rosso.
Questa metapopolazione diventa più stabile quando tutti
i demi sono monomorfi per il cromosoma mutato rosso e
il cromosoma standard verde o per il cromosoma mutato
arancio e il cromosoma standard azzurro
Questa metapopolazione diventa più stabile quando tutti
i demi sono monomorfi per il cromosoma mutato rosso e
il cromosoma mutato arancio o per il cromosoma
standard verde e il cromosoma standard azzurro
In entrambi i casi l’aumento di stabilità è dovuto all’aumento del tasso critico di migrazione, legato al fatto che due
mutazioni cromosomiche invece di una si affacciano sugli stessi confini tra i demi; questo spiega il tendenziale
accumulo di diverse varianti cromosomiche negli stessi demi, come succede in popolazioni di Mus musculus
domesticus.
Le mutazioni cromosomiche sottodominanti
sono fattori di isolamento riproduttivo
Riduzione del flusso genico di un gene A con 2 alleli neutrali A1 e A2
(WA1A1=WA1A2=WA2A2) a causa di una mutazione cromosomica sottodominante.
Popolazione monomorfa per
l’allele A1
Popolazione monomorfa per
l’allele A2
Entrambe le popolazioni sono monomorfe per il cromosoma standard.
m
m*
Una popolazione è monomorfa per il cromosoma standard, l’altra per il cromosoma mutato.
m
Per tassi di migrazione molto bassi (m<< mc), la presenza di
una mutazione cromosomica sottodominante in una delle due
popolazioni riduce fra di esse il flusso genico di un gene
neutrale come se ci fosse un tasso di migrazione più basso
(m*) in assenza della mutazione cromosomica .
m*/m= R(1-s)/(s+R(1-s)), in cui R = frequenza di ricombinazione fra il gene con alleli neutrali e il sito della
mutazione cromosomica; se R=0,5 (geni non associati alla mutazione cromosomica), m*/m= (1-s)/(1+s).
Le mutazioni cromosomiche sottodominanti non bloccano ma riducono il flusso genico di tutti i geni.
Le inversioni sopprimono la ricombinazione
intracromosomica, le traslocazioni e le fusioni centriche
riducono la ricombinazione intercromosomica
INVERSIONI
paracentriche
A
C
a
b
A
C
D
B
B
c
D
d
b
c
TRASLOCAZIONI
Regione coinvolta
nell’inversione
Cromatidi bilanciati,
vitali senza crossing over
nella regione invertita
a
d
Cromosomi non omologhi
coinvolti nella traslocazione
Cromatidi instabili, non
vitali con crossing over
nella regione invertita
E
C
B
d
D
b
c
a
A
C
b
a
D
B
c
d
Cromatidi bilanciati,
vitali senza crossing over
nella regione invertita
Cromatidi sbilanciati, non
vitali con crossing over
nella regione invertita
Sono trasmessi alla progenie
vitale solo i cromatidi che non
hanno subito la ricombinazione
entro la regione invertita
N
I
e
L
H
i
f
l m
g
n
h
Segregazione alternata, che produce cromosomi bilanciati,
vitali con le combinazioni parentali dei cromosomi e dei geni
E
pericentriche
A
F G M
F G M
N
i
n
l m
e
f
g
h
I
L
H
Segregazione adiacente, che produce cromosomi sbilanciati, non
vitali con le combinazioni ricombinanti dei cromosomi e dei geni
E
i
F G M n
I L
H
e
f
l m
g
N
h
Segregazione alternata con ricombinazione dei geni distali al
sito della traslocazione o, sull’altro braccio, al centromero.
Sono trasmessi alla progenie vitale solo le combinazioni parentali dei
cromosomi e degli alleli, con possibile ricombinazione dei geni distali
Le inversioni: selezione
senza ricombinazione
Nella popolazione sono presenti solo 2 combinazioni degli alleli entro l’inversione:
A c
B1
d
A B2 C
d
Regione invertita
Gli alleli B1 e B2 del gene B sono neutrali
L’allele recessivo c del gene C è svantaggiato
rispetto all’allele dominante C
Le uniche combinazioni possibili di alleli
per i geni B e C sono c-B1 e B2-C (le
combinazioni ricombinanti non sono
vitali)
L’allele neutrale
B1 subisce la
stessa variazione
di frequenza ad
ogni generazione
dell’allele c, fino
all’eliminazione:
Dp= -sp2q/W’
Le inversioni: alleli coadattati e supergeni
Regione coinvolta nell’inversione
Nella regione coinvolta
nell’inversione si trovano due alleli
(indicate con i numeri 1 e 2) per
ogni gene di una serie (da “a” a “z”)
indichiamo con W’1=W’a1-b1-….y1-z1 e con W’2=W’a2-b2-….y2-z2
indichiamo con W’11=W’a1a1-b1b1-….y1y1-z1 z1,
con W’22=W’a2a2-b2b2-….y2y2-z2z2
e con W’12=W’a1a2-b1b2-….y1y2-z1z2
m1-n1
Le fitness delle combinazioni degli alleli dei
supergeni si combinano in modo
a1-b1-… -y1-z1
St
n2-m2
moltiplicativo con quella dei cromosomi
che li portano:
z2-y2-… -b2-a2
Se le inversioni sono
Inv
piccole, non avvengono
Le 2 serie di alleli possono essere rotte per ricombinazione W’11StSt=W’11xW’StSt
l’appaiamento e il crossing
solo dal crossing over doppio, molto raro: pertanto, dal
W’22InvInv=W’22xW’InvInv
over; quindi non si realizza
punto di vista della segregazione e della selezione, si
W’12StInv=W’12xW’StInv
la riduzione di fecondità
comportano come un solo gene (supergene).
degli eterozigoti.
a1-b1-…-m1-n1-… -y1-z1
St
Inv
z2-y2-…-m2-n2… -b2-a2
In questo caso le inversioni diventano intrinsecamente
neutrali e la fitness dei cromosomi è dovuta interamente a
quella dei supergeni in essi presenti:
W’11StSt=W’11; W’22InvInv=W’22; W’12StInv=W’12
Si trovano in natura inversioni multiple, che rendono
complesso l’appaiamento e sbilanciati anche i prodotti del
doppio crossing over; quindi le 2 serie di alleli non
possono essere mai separate per ricombinazione.
Effetti del crossing over nella regione compresa
fra il centromero e il sito della traslocazione
Gamete sbilanciato
A
A
B C
B C
d
d
b c
E
A
A
D
D
B C
B C
E
b c
Gamete bilanciato
Gamete sbilanciato
Gamete bilanciato
A
Gamete bilanciato
Gamete sbilanciato
C
B
c
b
c
b
d
A
Segregazione
adiacente
a
a
a
E
C
B
c
b
c
b
d
Segregazione
alternata
e
ED
e
e
D
D
E
a
b c
E
a
a
e
e
e
Gamete bilanciato
ED
d
d
b c
E
Gamete sbilanciato
I gameti
bilanciati prodotti
3) I gameti
bilanciati prodotti
Nelle
centriche
e2)nelle
traslocazioni
chedalla
coinvolgono
il centromero,
il dalla
1) Metàfusioni/fissioni
dei gameti prodotti,
sia
segregazione adiacente hanno
segregazione alternata non hanno
centromero
coincide
con
della
segregazione
adiacente
cheildisito della traslocazione, quindi non c’è ricombinazione
ricombinazione intercromosomica
ricombinazione intercromosomica
quella
alternata, sono bilanciati
intercromosomica
per i geni adiacenti
al
centromero.
per i geni adiacenti al centromero
per i geni adiacenti al centromero
La distribuzione delle mutazioni cromosomiche
sottodominanti legata alla fitness dei supergeni
In caso di eterogeneità ambientale, su grande o piccola scala, le combinazioni degli alleli dei supergeni presenti nel
cromosoma invertito possono conferire in omozigosi un vantaggio selettivo rispetto alle combinazioni presenti nel
cromosoma standard in una matrice ambientale e uno svantaggio in un’altra.
W’11> W’22 in ambiente 1; W’11< W’22 in ambiente 2
Se, come ci si aspetta, l’inversione è sottodominante,
la combinazione moltiplicativa delle fitness porta al
vantaggio di un omozigote rispetto all’altro in un
ambiente e lo svantaggio in un altro ambiente.
W’11StSt> W’22InvInv in ambiente 1
W’11StSt< W’22InvInv in ambiente 2
Se la fitness complessiva del’omozigote per il
cromosoma invertito è maggiore o uguale a quella
dell’eterozigote, nell’ambiente vantaggioso, si ricade
nella selezione direzionale; se è minore, si rimane nella
selezione diversificatrice.
W’11StSt< W’12StInv in ambiente 2 W’11StSt> W’12StInv in ambiente 2
W’11StSt> W’12StInv in ambiente 1 W’11StSt>W’12StInv in ambiente 1
Selezione direzionale.
Selezione diversificatrice.
Deme monomorfo per il
cromosoma standard e per la
combinazione 1 degli alleli
Deme monomorfo per il
cromosoma invertito e per la
combinazione 2 degli alleli
Ambiente 1
Ambiente 2
Migrazione
La distribuzione dei demi monomorfi per il cromosoma
invertito tende a coincidere con l’estensione dell’ambiente in
cui è avvantaggiata la propria combinazione di alleli; la
sottodominanza stabilizza ulteriormente questa distribuzione.
Il ruolo delle mutazioni cromosomiche sottodominanti
nell’anagenesi, nella cladogenesi, nella speciazione
Le mutazioni cromosomiche sottodominanti
agiscono come fattori di isolamento
riproduttivo post zigotico; per tassi ridotti di
migrazione il loro flusso genico è bloccato e il
flusso genico per tutti i geni è ridotto, in
particolare per i geni associati a tali mutazioni.
Le mutazioni cromosomiche sottodominanti
tendono a occupare areali compatti; più
mutazioni cromosomiche tendono a fare
coincidere i propri areali; questi fattori
rafforzano la loro capacità di agire come fattori
di isolamento riproduttivo.
Quindi, non ostante le difficoltà e le particolari condizioni che
ne aiutano la fissazione (p. es loro origine in demi piccoli e
isolati) svolgono un ruolo importante nella cladogenesi.
Le mutazioni cromosomiche sottodominanti
riducono o impediscono la ricombinazione
dei geni ad esse associati; questo favorisce la
formazione di combinazioni di alleli
coadattati per questi geni.
Le mutazioni cromosomiche sottodominanti
tendono ad occupare areali coincidenti con la
presenza di ambienti cui le proprie combinazioni
di alleli sono più adatte di quelle dei cromosomi
standard.
Quindi svolgono un ruolo importante sia nella cladogenesi che nell’anagenesi.
È documentato il loro ruolo nella divergenza evolutivasia tra popolazione della stessa
specie che fra specie affini; dunque svolgono un ruolo attivo nella speciazione.
Il ruolo delle duplicazioni
nell’anagenesi: l’origine di nuovi geni
L
M’: allele
funzionale di M
M N
Duplicazione
(“neutrale”)
L
M M N
Mutazioni
geniche
M
L
M M’ N
L
M m1 N
L
M m2 N
Diverse
mutazioni
geniche
L
m1, m2: alleli non
funzionali di M
Regione duplicata
m, m’: prodotti
del gene M
O: nuovo gene
m
Sostanza 1
L’individuo con il nuovo
gene O è avvantaggiato
o
Sostanza 3
Sostanza 2
m’
Sostanza 1
MO N
Sostanza 2
Sostanza 4
o: prodotto
del gene O
Esempi di mutazioni cromosomiche sottodominanti
che marcano la macroevoluzione dei primati
Il cromosoma 2 umano deriva dalla fusione centrica di 2 cromosomi acrocentrici intervenuta
dopo la divergenza dalle specie viventi più vicine (scimpanzè e bonobo).
Cromosomi non omologhi acrocentrici
ancestrali conservati in tutte le
Fusione centrica
catarrine
Cromosoma 2 umano
I cromosomi 14 e 15 umani, comuni alle altre scimmie antropomorfe derivano dalla fissione
centrica di un cromosoma metacrocentrico accompagnato da uno “spostamento” del centromero
intervenuta dopo la divergenza dalle altre catarrine.
Cromosoma metacrocentrico ancestrale
Fissione centrica +
spostamento del centromero
Cromosomi 14 e 15
umani, comuni alle altre
scimmie antropomorfe
Su una scala più ampia, tra il gibbone e l’uomo c’è una differenza cariotipica riconducibile a 21
diverse traslocazioni reciproche intervenute dopo la divergenza evolutiva.
La presenza di questi riordinamenti cromosomici testimoniano dell’occorrenza deelle condizioni
particolari compatibili con la fissazione e l’espansione di mutazioni cromosomiche
sottodominanti durante le diverse fasi della divergenza evolutiva, sulle quali è verosimile che tali
mutazioni abbiano esercitato un ruolo propulsivo.