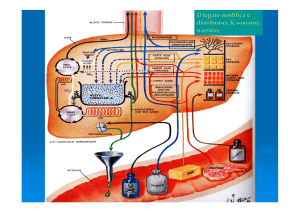
Utilizzo dei nutrienti dopo l'assorbimento
•
•
•
•
La velocità di assorbimento dei nutrienti dall'intestino non è costante, ma varia notevolmente in relazione all'ingestione di
cibo. La velocità di digestione dei pasti dipende dalla loro composizione chimica, indipendentemente dai fabbisogni nutritivi
dell'animale. La natura stessa della digestione prevede che l'assorbimento dei nutrienti sia rapido durante la digestione e si
arresti durante i periodi interdigestivi. In altre parole, l'intestino non è un deposito di nutrienti, e la digestione non è modulata
dal fabbisogno nutrizionale dell'animale. I fabbisogni nutritivi dell'animale non sono ben correlati ai notevoli intervalli
presenti tra gli assorbimenti dei nutrienti. Di contro, l'apporto costante di nutrienti che producano energia per mantenere le
funzioni metaboliche basali dell'organismo è di vitale necessità. Inoltre, i periodi in cui i fabbisogni metabolici dell'animale
sono particolarmente elevati spesso non coincidono con i tempi di veloce assorbimento di nutrienti dall'intestino. Di
conseguenza, l'animale deve essere in possesso di un sistema sofisticato per assicurare l'apporto costante di nutrienti,
particolarmente per quelli che sono fonte di energia, bilanciando l'effetto "abbondanza o carestia" a breve e lungo termine che
è associato al periodo digestivo pre e post assorbimento.
I MECCANISMI OMEOSTATICI REGOLANO L'APPORTO E LA RICHIESTA DI QUASI TUTTI I NUTRIENTI
Questo capitolo pone l'attenzione sulla regolazione dell'apporto dei principali nutrienti energetici; in ogni caso, anche altre
sostanze, tra cui vitamine e minerali, sono soggette a meccanismi di regolazione omeostatica. Anche se molti di questi
meccanismi coinvolgono direttamente il sistema digerente, per ragioni di spazio non saranno tutti descritti in questo libro. Per
la descrizione dei meccanismi omeostatici che regolano l'apporto di minerali e vitamine si può far riferimento ad alcune voci
bibliografiche riportate alla fine del capitolo.
I nutrienti energetici sono riportati come fonti metaboliche, ed i meccanismi fisiologici che mantengono l'apporto di sostanze
uniformandole alle richieste sono noti come omeostasi della fonte energetica. L'omeostasi della fonte energetica è mantenuta
da diversi meccanismi: l'asse insulina-glucagone, l'asse ipotalamo-ipofisi, ed il sistema nervoso centrale.
Fonti energetiche
•
•
•
•
•
•
•
•
II CICLO DEGLI ACIDI TRICARBOSSILICI (KREBS) È LA PRINCIPALE VIA ENERGETICA
DELL'ORGANISMO PER L'UTILIZZO DELLE FONTI ENERGETICHE
II ciclo di Krebs e le vie associate del metabolismo intermedio non sono solo sede di utilizzo della fonte energetica e di
produzione di energia, ma anche di trasformazione di una fonte energetica in un'altra. Queste trasformazioni sono importanti
nel quadro generale dell'omeostasi della fonte energetica.
LE PRINCIPALI FONTI ENERGETICHE METABOLICHE SONO: GLUCOSIO, AMINOACIDI, ACIDI GRASSI
E CORPI CHETONICI; PER QUESTI COMPOSTI ESISTONO DIVERSE FORME DI DEPOSITO E DI
TRASPORTO
II glucosio, prodotto della digestione dei carboidrati, è la fonte energetica metabolica di base dei monogastrici onnivori, come
cani e ratti. Nonostante la presenza di diverse importanti fonti energetiche nell'organismo, il glucosio assume un'importanza
particolare in quanto, nella maggior parte dei casi, è l'unica fonte energetica per il sistema nervoso centrale. Di conseguenza,
assicurare un apporto di glucosio per il metabolismo cerebrale è di fondamentale importanza per l'organismo. Non sorprende
quindi, che un raffinato sistema omeostatico regoli la disponibilità di glucosio al cervello ed agli altri tessuti. Il principale
obiettivo di questo capitolo è la descrizione di quei sistemi volti a garantire la disponibilità di glucosio per l'organismo.
IL GLUCOSIO È LA PRINCIPALE FONTE ENERGETICA NEL METABOLISMO DELLA MAGGIOR PARTE
DEGLI ANIMALI
II glucosio può essere conservato sotto forma di glicogeno, un amido altamente ramificato che si trova nel fegato e nel
muscolo scheletrico. Il glicogeno è l'unica forma diretta di deposito di glucosio nell'organismo, anche se il glucosio può
essere sintetizzato a partire da altri composti.
La principale funzione dell'omeostasi della fonte energetica è destinare il glucosio ai depositi o rimuoverlo dagli stessi. Il
processo di liberazione di glucosio dal glicogeno è denominato glicogenolisi.
•
•
•
•
•
•
Il principale sistema di utilizzo del glucosio ai fini energetici è la via di Embden-Meyerhof, denominata anche glicolisi. La
glicolisi è rappresentata da una serie di tappe biochimiche che prende il via dall'ossidazione del glucosio. La glicolisi porta
direttamente al ciclo di Krebs, sito della completa ossidazione della fonte energetica e principale via metabolico-energetica
dell'organismo. Per lo studio dell'omeostasi della fonte energetica bisogna tener presente che la glicolisi è un processo
reversibile, nel senso che il glucosio può essere prodotto a partire dai prodotti terminali della glicolisi. Dato lo stretto legame
tra glicolisi e ciclo di Krebs, ognuno dei prodotti intermedi del ciclo di Krebs può rientrare nuovamente nella via glicolitica
per produrre glucosio. La sintesi di glucosio dai prodotti terminali della glicolisi e da quelli intermedi del ciclo di Krebs,
definita gluconeogenesi, è parte fondamentale dell'omeostasi della fonte energetica. Anche se l'attività del ciclo di Krebs si
svolge virtualmente in tutti i tessuti, tranne i globuli rossi, il processo di gluconeogenesi si svolge solo nel fegato e,
limitatamente, nel rene.
Un'altra via di ossidazione del glucosio è quella dei pentoso-fosfati. Un sistema quantitativamente inferiore che non ha
grande impatto sull'omeostasi della fonte energetica. D'altro canto, essa è un'importante via metabolica per gli eritrociti, che
hanno un'assoluta necessità di glucosio, anche se la necessità energetica di queste cellule è ben poca rispetto a quella
dell'organismo.
GLI AMINOACIDI, OLTRE AD ESSERE I COSTITUENTI DELLE PROTEINE, SONO ANCHE UNA FONTE
ENERGETICA IMPORTANTE
Gli aminoacidi sono una fonte energetica importante. Oltre ad essere i mattoni delle proteine, contengono carbonio che può
fornire energia all'organismo. Inoltre, essi rappresentano un importante substrato per la gluconeogenesi, nel senso che (la
maggior parte) possono essere convcrtiti a glucosio quando l'apporto di questo è scarso. Anche se non viene riportato alcun
tipo di deposito di aminoacidi, le proteine dei muscoli scheletrici, oltre alle loro funzioni locomotrici, possono essere
considerate un sistema di deposito di aminoacidi.
GLI ACIDI GRASSI SONO LA PRINCIPALE FORMA DI DEPOSITO ENERGETICO DELL'ORGANISMO
ANIMALE
Gli acidi grassi vengono conservati nel tessuto adiposo sotto forma di trigliceridi (anche chiamati triacilgliceroli), che
consistono di tré molecole di acidi grassi legate ad una molecola di glicerolo mediante legame estere.
•
•
•
•
•
Per gli animali, i trigliceridi rappresentano la forma ideale di deposito di energia. Si tratta di molecole altamente ridotte (poco
ossigeno rispetto a carbonio ed idrogeno), il che le rende fonti di energia concentrata, con un valore calorico doppio rispetto a
carboidrati ed. aminoacidi. Inoltre, il tessuto adiposo contiene poca acqua rispetto a proteine e glicogeno, le altre forme di
deposito delle fonti energetiche. Di conseguenza, il tessuto adiposo, non risultando diluito dall'acqua interstiziale, rappresenta
una forma concentrata di energia che permette all'animale di ottenere la massima resa energetica portando un peso minimo.
Di contro, i grassi presentano uno svantaggio metabolico notevole, non sono idrosolubili, per cui sono necessari particolari
sistemi di trasporto per la loro distribuzione ai tessuti attraverso sangue e sistema linfatico. Ancora, gli acidi grassi non
possono essere convcrtiti in glucosio e quindi non possono, in condizioni fisiologiche, apportare energia al sistema nervoso
centrale, possono, però, essere convcrtiti in corpi chetonici.
I CORPI CHETONICI SONO METABOLICI IDROSOLUBILI DEI GRASSI CHE FUNGONO DA SOSTITUTI
DEL GLUCOSIO
Anche se la conversione dei grassi in glucosio non è possibile, i corpi chetonici presentano diverse caratteristiche simili a
questo monosaccaride. Ad esempio, i corpi chetonici possono attraversare la barriera emato-encefalica e, in condizioni di
digiuno prolungato, possono sostenere gran parte delle necessità energetiche del sistema nervoso centrale, almeno in alcune
specie. È anche vero però, che i corpi chetonici non possono sostituire completamente il glucosio, e quindi una minima
quantità di quest'ultimo è comunque necessaria per le funzioni del sistema nervoso centrale.
Nelle specie monogastriche, i corpi chetonici si formano esclusivamente nel fegato e vengono utilizzati da molti tessuti,
alcuni di questi, tra cui il muscolo cardiaco, li preferiscono al glucosio come fonte energetica. Nei ruminanti, il betaidrossibutirrato si forma dal butirrato nell'epitelio ruminale, quindi, in questa specie, i corpi chetonici non sono solo un
prodotto metabolico degli acidi grassi, ma anche prodotto dei normali processi digestivi.
L'aumento delle concentrazioni seriche dei corpi chetonici è frequente in diverse patologie associate ad alterazioni
dell'omeostasi della fonte energetica. Questa affermazione potrebbe far erroneamente pensare che i corpi chetonici siano
metaboliti anomali e potenzialmente tossici, mentre, a concentrazioni fisiologiche essi rappresentano un importante fonte
energetica che occupa un posto preciso nello schema omeostatico.
Utilizzo dei nutrienti durante la fase di assorbimento
•
•
•
•
•
•
•
Al momento dell'assorbimento, diversi eventi metabolici a livello epatico e tissutale si coordinano per dirigere i nutrienti
verso la formazione di molecole e dei siti di deposito. Lo schema generale del metabolismo durante la fase di assorbimento è
illustrato in Figura 31-3.
DURANTE LA FASE DI ASSORBIMENTO, IL FEGATO CAPTA IL GLUCOSIO E LO CONVERTE IN
GLICOGENO E TRIGLICERIDI
Durante l'assunzione di un pasto, la secrezione di insulina comincia prima che venga raggiunto il massimo assorbimento di
glucosio. Tale secrezione è stimolata dall'azione del peptide gastro-inibitore e forse da altri ormoni enterici. La secrezione
precoce di insulina assicura che fegato ed altri tessuti siano "pronti" all'arrivo del glucosio dall'intestino. Gran parte del
glucosio postprandiale viene preso dal fegato, in quanto questo è estremamente irrorato, e presenta un'elevata capacità di
captazione del glucosio. Sotto l'influenza dell'insulina, il glucosio nel fegato viene utilizzato per la sintesi di glicogeno.
L'effetto finale è quello di convertire in glicogeno, il glucosio, durante i periodi di assorbimento, evitando, in questo modo,
l'instaurarsi di iperglicemia. L'insulina stimola le vie metaboliche che portano alla formazione di glicogeno.
Questi effetti verranno discussi successivamente insieme all'effetto controbilanciante del glucagone.
La quantità di glicogeno che può essere conservata nel fegato è limitata e non supera, in condizioni normali, il 10% del peso
totale dell'organo. Nell'uomo, questo significa circa 100 g di glicogeno e sembra che una simile proporzione esista anche per
le altre specie. Questa quantità di glicogeno non rispecchia tutto il glucosio captato dal fegato durante la digestione e
l'assorbimento di un pasto ricco di carboidrati, quindi, devono esistere altri meccani smi per contenere l'eccesso di glucosio.
Se non esistessero vie alternative, la glicemia aumenterebbe eccessivamente una volta raggiunta la massima concentrazione
di glicogeno. La sintesi di acidi grassi rappresenta una via alternativa per lo smaltimento del glucosio.
LA CONVERSIONE DI GLUCOSIO IN ACIDI GRASSI È UN PROCESSO IRREVERSIBILE
La sintesi di acidi grassi dal glucosio comincia con la glicolisi. Questa via metabolica porta alla formazione di due molecole
di piruvato per ogni molecola di glucosio consumato. Il piruvato può quindi entrare nei mitocondri, essere convertito in acetil
coenzima A (acetil CoA) ed entrare nel ciclo di Krebs. La funzione del ciclo di Krebs è di generare energia e durante il
periodo di assorbimento, la quantità di acetil CoA è più che sufficiente; per tale motivo risulta necessario, quindi, allontanarlo
dal ciclo stesso. L'eccesso di acetil CoA si combina con l'ossalacetato per formare citrato, in pratica, la prima reazione del
ciclo di Krebs. La maggior parte del citrato abbandona i mitocondri e si porta nel citosol dove, da ogni molecola derivano due
molecole di carbonio per la sintesi di acidi grassi. La porzione rimanente rientra nei mitocondri per essere utilizzata
successivamente. Quindi, dato che l'acetil CoA non è in grado di attraversare la membrana mitocondriale, il citrato funge da
molecola di trasporto per due unità di carbonio acetato, originato dal glucosio, fuori dai mitocondri per la sintesi di acidi
grassi nel citosol.
•
•
•
•
•
Diverse tappe importanti di questa reazione sono stimolate dall'insulina e verranno ampiamente descritte successivamente. È
importante ricordare che la conversione di glucosio in acidi grassi è un processo irreversibile, quindi i carboidrati possono
formare grassi, ma i grassi non possono formare carboidrati. Il metabolismo epatico è strettamente implicato in questi
meccanismi ed il fegato è una sede importante per la formazione di acidi grassi in molte specie. La sintesi diretta di acidi
grassi avviene anche nel tessuto adiposo. L'importanza di fegato e tessuto adiposo per la sintesi di acidi grassi nelle varie
specie sarà discussa successivamente.
IL TRASPORTO DEGLI ACIDI GRASSI AL DI FUORI DEL FEGATO AVVIENE ATTRAVERSO PARTICELLE
SIMILI AI CHILOMICRONI, DENOMINATE LIPOPROTEINE A BASSISSIMA DENSITÀ
Una volta formati nel fegato, gli acidi grassi devono essere trasportati al tessuto adiposo come riserva, e ad altri tessuti, come
i muscoli, per fornire energia tramite il loro diretto utilizzo. Essendo insolubili nel sangue, gli acidi grassi hanno bisogno di
speciali sistemi di trasporto. Ciò avviene attraverso la formazione epatica di lipoproteine ricche di trigliceridi denominate
very low density lipoprotein o lipoproteine a bassissima densità o VLDL. Tale denominazione deriva da una densità molto
inferiore a quella delle altre proteine seriche. Nella sintesi delle VLDL, gli acidi grassi vengono primariamente esterificati a
trigliceridi, e questi successivamente sono intrappolati in uno strato di fosfolipidi, colesterolo e proteine specifiche. Si tratta
essenzialmente dello stesso meccanismo mediante il quale gli acidi grassi vengono trasportati al di fuori dell'enterocita dopo
l'assorbimento intestinale, solo che in quel caso le lipoproteine erano denominate chilomicroni. Le VLDL epatiche sono più
piccole dei chilomicroni, ma hanno struttura e funzioni simili. I meccanismi mediante i quali VLDL e chilomicroni vengono
distribuiti ai tessuti periferici saranno discussi successivamente.
GLI AMINOACIDI POSSONO ESSERE CLASSIFICATI IN GRUPPI SULLA BASE DELLE CARATTERISTICHE
METABOLICHE
La descrizione dell'assorbimento e del metabolismo degli aminoacidi è complicata per le diverse caratteristiche degli
aminoacidi stessi. Per questo motivo, gli aminoacidi vengono divisi in due gruppi, ognuno formato da due sottogruppi. I due
gruppi principali sono gli aminoacidi essenziali e quelli non essenziali. Tra quelli essenziali, glutammato, aspartato ed alanina
sono denominati aminoacidi di trasporto, tra quelli non essenziali, leucina, isoleucina e valina formano un gruppo
denominato aminoacidi a catena ramificata (BCCAAs). Gli aminoacidi di trasporto intervengono in diverse reazioni dove
aminogruppi vengono trasferiti da una molecola all'altra o da un organo ad un altro.
•
•
•
•
•
•
•
•
•
GLI AMINOACIDI SONO ESTREMAMENTE MODIFICATI DURANTE L'ASSORBIMENTO
II profilo aminoacidico nel sangue della vena porta è molto diverso da quello della dieta, il che indica una distruzione e
trasformazione degli aminoacidi durante i processi di assorbimento. Quasi tutto il glutammato, e la maggior parte
dell'aspartato, vengono rimossi dalle cellule dell'epitelio intestinale, durante l'assorbimento, cosicché a livello portale è
possibile trovare solo piccole quantità di aspartato. La maggior parte dell'azoto del glutammato ed aspartato viene trasferito al
piruvato per formare alanina, presente in grandi quantità nel sangue portale. Il metabolismo del trasporto degli aminoacidi
nell'epitelio intestinale è un buon esempio sia di come gruppi aminici possano essere persi e acquisiti, sia di come il
metabolismo degli aminoacidi si interface! con quello dei carboidrati.
Glutammato ed aspartato sono simili a due prodotti intermedi del ciclo di Krebs, l'a-chetoglutarato e l'ossalacetato, da cui
differiscono solo per la presenza di un aminogruppo o di un cheto-ossigeno. Carboidrati ed aminoacidi con tale caratteristica
vengono denominati analoghi; l'a-chetoglutarato è un cheto-analogo del glutammato ed il piruvato è un cheto-analogo
dell'alanina (Fig. 31-6).
Tutti gli aminoacidi possono formare cheto-analoghi e tutti i cheto-analoghi possono essere riconvertiti in aminoacidi.
MOLTI AMINOACIDI VENGONO RIMOSSI DAL FEGATO SEDUTA STANTE, SENZA MAI RAGGIUNGERE
LA CIRCOLAZIONE SISTEMICA
La circolazione portale epatica è organizzata in modo tale che tutti i nutrienti che lasciano l'intestino per via ematica, passino
attraverso il fegato prima di raggiungere la circolazione sistemica. Ciò attribuisce al fegato il ruolo di "sentinella" in grado di
modificare la
composizione dei nutrienti del sangue portale prima che questo giunga agli altri tessuti. Questa funzione del fegato è ben
rappresentata dal caso dell'assorbimento degli aminoacidi. Gran parte degli aminoacidi del sangue portale vengono rimossi
durante il loro transito epatico e non raggiungono mai il circolo sistemico.
ALCUNI AMINOACIDI CAPTATI DAL FEGATO VENGONO UTILIZZATI PER LA SINTESI DI PROTEINE
La posizione primaria del fegato nella captazione degli aminoacidi è giustificata dal fatto che esso rappresenta un'importante
sede per la sintesi di proteine. La Figura 31-7 mostra che circa il 20% del contenuto aminoacidico portale viene utilizzato a
livello epatico per la sintesi di proteme, anche se tale percentuale dipende dal tenore proteico della dieta. Quasi tutte le
proteine seriche vengono sintetizzate nel fegato, comprese alcune fondamentali quali albumine e fattori della coagulazione.
Le proteine seriche di origine epatica svolgono numerose funzioni, ma non quella di trasporto degli aminoacidi. Infatti,
l'apporto di aminoacidi per le sintesi proteiche extra epatiche è dato dagli aminoacidi liberi del sangue e non dalle proteine
seriche preformate.
•
•
•
•
•
LA MAGGIOR PARTE DEGLI AMINOACIDI CAPTATI DAL FEGATO VENGONO CONVERTITI IN
CARBOIDRATI
La maggior parte degli aminoacidi che arrivano al fegato vanno incontro a deaminazione, cioè alla rimozione di alcuni
aminogruppi, e conseguente conversione nel chetoanalogo specifico. I cheto-analoghi entrano nel metabolismo dei
carboidrati per essere utilizzati a scopo energetico, per la sintesi di acidi grassi, o convertiti a glucosio o glicogeno. Tutte
queste reazioni si svolgono come già descritto nella sezione relativa al metabolismo dei carboidrati. La deaminazione di
aminoacidi per la produzione di carboidrati, o energia, potrebbe sembrare una perdita delle proteine contenute nella dieta ma,
in alcune specie, questo processo è importante per mantenere Fomeostasi del glucosio e di altre fonti energetiche. Le diete
naturali dei veri carnivori, come gatti e visoni, per esempio, contengono molte proteine e pochi carboidrati e, dato che questi
animali hanno gli stessi fabbisogni di glucosio degli altri, è estremamente importante che il glucosio venga sintetizzato dagli
aminoacidi. I ruminanti presentano una situazione analoga, poiché la maggior parte del glucosio viene utilizzato per le
fermentazioni, ed assorbito come acidi grassi volatili. I ruminanti quindi, come i carnivori, dipendono dagli aminoacidi per
parte del fabbisogno in glucosio, anche se la quota maggiore di glucosio deriva dalla conversione del propionato. Per
permettere la produzione di carboidrati e la deaminazione degli aminoacidi in eccesso, le reazioni endocrine ad un pasto ricco
di proteine sono differenti da quelle che avverrebbero nel caso di un pasto a base di carboidrati. Durante la digestione di pasti
iperproteici, la secrezione di insulina e glucagone non avviene con le modalità normali. La secrezione di insulina è stimolata
sia dagli aminoacidi sia dal glucosio. La secrezione di glucagone, inibita dal glucosio, è stimolata dagli aminoacidi fin tanto
che la glicemia si mantiene entro livelli relativamente bassi. Ciò vuoi dire che, in caso di pasti iperproteici ed ipoglucidici, si
ha una stimolazione contemporanea di insulina e glucagone. Uno degli effetti dell'insulina è quello di aumentare sia la
captazione di aminoacidi sia quella di glucosio, per cui, l'effetto dell'insulina in questo caso è quello di aumentare il
trasferimento di aminoacidi ai tessuti.
Se la secrezione di insulina fosse il solo effetto stimolato dall'assorbimento di aminoacidi, l'animale rischierebbe
un'ipoglicemia insuline-dipendente in seguito ad un pa sto iperproteico ed ipoglucidico. Un'importante azione del glucagone
è quella di stimolare, a livello epatico, la gluconeogenesi tramite deaminazione degli aminoacidi; questo processo garantisce
una quantità di glucosio idonea a controbilanciare gli effetti della secrezione di insulina stimolata dagli aminoacidi.
NON TUTTI GLI AMINOACIDI SUBISCONO UN METABOLISMO EPATICO
Durante il periodo di assorbimento, gli aminoacidi per le sintesi proteiche periferiche (non epatiche) derivano da quella quota
aminoacidica che sfugge al metabolismo epatico. Tale quota corrisponde solo al 23% degli aminoacidi assorbiti a livello
intestinale.
•
•
•
•
•
•
•
Anche se questa può sembrare una percentuale minima per le sintesi proteiche di tutto l'organismo, alcune considerazioni
dimostreranno che non è così. In primo luogo, gli aminoacidi vengono captati dal fegato in maniera selettiva, per cui la
concentrazione dei singoli aminoacidi che lascia il fegato è diversa da quella che vi arriva. Gli aminoacidi essenziali,
specialmente i BCAA, non vengono avidamente utilizzati dal fegato, laddove alcuni degli essenziali, ad esempio l'alanina,
sono massivamente sequestrati dal tessuto epatico. Gli aminoacidi essenziali possono essere sintetizzati dai tessuti che
producono proteine, quindi, la concentrazione serica relativamente bassa dovuta alla captazione epatica non limita le sintesi
proteiche tissutali. In secondo luogo, la quota di aminoacidi captata dal fegato, ed il loro destino non sono costanti ma
variano in relazione alle necessità proteiche dell'organismo. Diete a basso contenuto proteico portano a riduzione della
captazione epatica di aminoacidi, della sintesi proteica e del metabolismo aminoacidico a livello epatico.
IL METABOLISMO A LIVELLO TISSUTALE È COORDINATO CON QUELLO EPATICO E DA LUOGO ALLA
DEPOSIZIONE DI FONTI ENERGETICHE NEI TESSUTI DI DEPOSITO DURANTE IL PERIODO DI
ASSORBIMENTO
Gli effetti generali del metabolismo epatico durante l'assorbimento di un pasto sono: la rimozione di glucosio ed aminoacidi e
la sintesi di proteine e grassi. Nei tessuti periferia si verificano cambiamenti complementari volti a rimuovere dal muscolo
scheletrico e tessuto adiposo gli aminoacidi ed il glucosio in eccesso. Inoltre, gli acidi grassi secreti dal fegato sotto forma di
trigliceridi VLDL si depositano nel tessuto adiposo, così come avviene per i chilomicroni.
L'INSULINA PROMUOVE LA SINTESI DI PROTEINE E LA DEPOSIZIONE DI GLICOGENO A LIVELLO
MUSCOLARE
II periodo di assorbimento è controllato dagli effetti dell'insulina. Nel muscolo scheletrico, che rappresenta la maggior parte
del corpo, l'insulina promuove la captazione di aminoacidi e glucosio, moderando l'aumento di questi nutrienti durante
l'assorbimento di un pasto. La captazione muscolare di glucosio è associata alla sintesi di glicogeno, come avviene nel fegato.
Il glicogeno muscolare, contrariamente a quello epatico, non può essere reso immediatamente disponibile per aumentare il
tasso ematico di glucosio in caso di ipoglicemia. Quindi, in questa sede il glicogeno viene utilizzato principalmente per il
metabolismo muscolare, anche se, in alcuni casi, esso può indirettamente rappresentare un substrato per la gluconeogenesi
epatica.
LA CAPTAZIONE MUSCOLARE DI AMINOACIDI STIMOLATA DALL'INSULINA PORTA AD UN NETTO
AUMENTO DELLE SINTESI PROTEICHE A LIVELLO MUSCOLARE
II termine aumento netto viene usato in relazione alle sintesi proteiche muscolari, perché le proteine muscolari, presentando
un flusso costante, sono un indice dell'equilibrio dinamico.
•
•
•
•
•
Le molecole proteiche vanno incontro a continuo catabolismo ed i loro aminoacidi entrano a far parte di un pool
aminoacidico intracellulare. Contemporaneamente, a partire da questo pool si formano costantemente nuove proteine. La
dimensione del pool aminoacidico dipende dalla quantità di aminoacidi che entrano ed escono. Gli aminoacidi entrano nel
pool dal sangue durante la fase di assorbimento e nello stesso tempo derivano dal catabolismo proteico. La fuoriuscita dal
pool avviene grazie alla sintesi proteica ed al catabolismo ossidativo. Durante la fase di assorbimento, legata alla digestione,
grazie alla notevole presenza di aminoacidi, le dimensioni del pool sono notevoli. Inoltre, pochi aminoacidi del pool vanno
incontro a catabolismo ossidativo, ciò grazie alla ampia disponibilità di glucosio per l'ossidazione e produzione di energia. Il
risultato è la presenza di un ampio pool aminoacidico, e tali aminoacidi sono utilizzati principalmente per la sintesi proteica.
Quando la velocità di sintesi delle proteine supera quella del loro catabolismo, si verifica un aumento netto delle proteine
muscolari. Quindi, durante la fase di assorbimento gli aminoacidi vengono conservati nelle proteine muscolari che, oltre a
svolgere un ruolo funzionale per locomozione e postura, fungono anche da deposito aminoacidico.
DURANTE LA FASE DI ASSORBIMENTO, L'ACCUMULO DI TRIGLICERIDI NEL TESSUTO ADIPOSO
AVVIENE MEDIANTE DUE MECCANISMI: CAPTAZIONE DALLE LIPOPROTEINE A BASSISSIMA DENSITÀ
E DIRETTA SINTESI LIPIDICA A PARTIRE DAL GLUCOSIO
I trigliceridi vengono trasferiti da chilomicroni e VLDL al tessuto adiposo grazie all'azione della lipoproteinlipasi (LPL).
Questo enzima si trova sulla superficie endoteliale dei capillari e, quando attivata, si lega a chilomicroni e VLDL, catalizza
l'idrolisi degli acidi grassi dai trigliceridi, permettendone il trasferimento ai tessuti circostanti. La sensibilità della LPL a
specifici ormoni varia a seconda dei tessuti. La LPL del tessuto adiposo è stimolata dall'insulina, quindi, durante la fase di
assorbimento, gli acidi grassi dei trigliceridi dei chilomicroni e le VLDL vengono trasferiti selettivamente alle cellule. A
questo punto, per l'influenza dell'insulina, durante la fase di assorbimento, aminoacidi e carboidrati in eccesso vengono
convcrtiti in acidi grassi a livello epatico, e questi vengono poi trasportati al tessuto adiposo tramite i trigliceridi delle VLDL.
Sempre per lo stimolo insulinico, i chilomicroni derivanti dall'assorbimento intestinale degli acidi grassi, raggiungono il
tessuto adiposo in maniera simile.
Oltre che da chilomicroni e VLDL, gli acidi grassi del tessuto adiposo possono anche derivare da una sintesi diretta a livello
locale. Le cellule del tessuto adiposo sono metabolicamente attive e, sotto lo stimolo dell'insulina, possono captare glucosio.
Negli adipociti, il glucosio può essere convcrtito in acidi grassi tramite lo stesso meccanismo che si realizza nel fegato.
Inoltre, anche l'acetato che deriva dai processi fermentativi intestinali può rappresentare un substrato per la sintesi di acidi
grassi nel tessuto adiposo (vedi le particolari considerazioni sulle fonti energetiche nei ruminanti). In conclusione, sono
presenti nell'organismo due sedi principali per la sintesi di acidi grassi: il fegato ed il tessuto adiposo. La relativa importanza
dei due tessuti è specie-specifica.
Utilizzo dei nutrienti durante
la fase successiva all'assorbimento
•
•
•
•
La fase successiva all'assorbimento è un periodo relativamente breve (generalmente poche ore) che intercorre tra un pasto e
l'altro. Essa è caratterizzata da cambiamenti a breve termine che mobilizzano i nutrienti dai pool di deposito per garantire la
disponibilità di fonte energetica ai tessuti metabolicamente attivi.
IL METABOLISMO EPATICO PERMETTE SIA L'UTILIZZO, SIA LA PRODUZIONE DI GLUCOSIO DURANTE
LA FASE SUCCESSIVA ALL'ASSORBIMENTO
Quando l'assorbimento di un pasto è completato, la quantità di glucosio riassorbito a livello intestinale si riduce, e la glicemia
si abbassa, inibendo lo stimolo per la produzione di insulina e stimolando la secrezione di glucagone. Quest'ultimo agisce
prevalentemente a livello epatico, inducendo notevoli cambiamenti metabolici. Tramite la stimolazione di specifici recettori
di membrana degli epatociti, il glucagone attiva l'adenilciclasi, portando alla fosforilazione di numerosi enzimi cellulari.
Alcuni enzimi vengono attivati mediante fosforilazione, altri inattivati e, se non si considera lo schema generale delle fonti
energetiche, l'intero sistema di fosforilazione e defosforilazione sembraavere poco senso. Se invece si considera l'azione dei
singoli enzimi, alla luce dei loro effetti sul flusso dei substrati energetici attraverso il fegato, ci si rende conto che si tratta di
un meccanismo elegante ed incredibilmente ben orchestrato atto al mantenimento dell'omeostasi della fonte energetica.
Gli enzimi che stimolano la mobilizzazione e l'utilizzo delle fonti energetiche vengono attivati tramite fosforilazione, mentre
quelli che ne stimolano il deposito vengono inattivati. Bisogna tener presente che molti enzimi del metabolismo intermedio
svolgono un ruolo passivo, catalizzando reazioni che possono andare in entrambe le direzioni a seconda della concentrazione
del substrato. Alcuni enzimi di regolazione sono spesso responsabili di vie metaboliche poiché determinano la
concentrazione di un substrato che si rende disponibile per altri enzimi privi di un sistema di regolazione.Tramite la sua
azione su alcuni enzimi chiave, il glucagone, uno stimolatore della fosforilazione, pone il fegato in uno stato di
mobilizzazione di fonte energetica. Di contro, l'insulina - inibitore della fosforilazione - promuove, a livello epatico, un
pattern metabolico volto a creare riserve, come descritto nel capitolo relativo alla fase di assorbimento.
•
•
•
•
•
L'azione opposta di insulina e glucagone a livello epatico si evince dai loro effetti su due paia di enzimi chiave: sintetasi e
glicogeno fosfatasi da un lato, fosfofruttochinasi e fruttosio 1-6 difosfatasi dall'altra. I primi regolano la sintesi di glicogeno
ed il catabolismo, i secondi, rispettivamente, glicolisi e gluconeogenesi.
La glicogeno sintetasi e la fosfofruttochinasi sono inibite dalla fosforilazione e quindi stimolate dall'insulina. La glicogeno
fosfatasi e la fruttosio 1,6 difosfatasi sono stimolate dalla fosforilazione e quindi dal glucagone. L'azione di insulina e
glucagone su queste coppie di enzimi antagonisti, enfatizza l'importanza del rapporto insulina/glucagone al quale il fegato
viene esposto. L'ormone non induce una reazione del "tutto o nulla", piuttosto, altera l'equilibrio di reazioni opposte,
influenzando l'attività relativa di enzimi antagonisti. Di conseguenza, la mobilizzazione o il deposito di fonti energetiche a
livello epatico dipende da quale è l'ormone dominante. Per queste ragioni e per quanto riguarda il fegato, il rapporto tra
insulina e glucagone sembra essere più importante delle concentrazioni ematiche di un singolo ormone. Sotto l'influenza del
glucagone, la glicogeno fosfatasi, attivata tramite fosforilazione, promuove la glicogenolisi ed un aumento della
concentrazione intracellulare di glucosio. Mentre il glucosio aumenta, ne viene impedita la riconversione a glicogeno, dato
che l'enzima responsabile di tale reazione, la glicogeno sintetasi, è bloccato dalla fosforilazione.
Inoltre, anche il flusso di glucosio verso la glicolisi è im pedito dal blocco della fosfofruttochinasi, sempre tramite
fosforilazione. Quindi, il normale utilizzo del glucosio all'interno dell'epatocita è impedito dal glucagone, il che permette un
accumulo intracellulare del glucosio derivante dal catabolismo del glicogeno.
All'occorrenza, il glucosio intracellulare può diffondere negli spazi interstiziali e quindi nel sangue. In questo modo, il
glicogeno epatico viene mobilizzato per aumentare e mantenere stabile la glicemia quando essa comincia a diminuire.
I depositi epatici di glicogeno sono relativamente limitati e non possono mantenere la concentrazione di glucosio ematico a
lungo. Si calcola che, nell'uomo, le riserve epatiche di glicogeno possano mantenere stabile la glicemia per 6-12 ore in
condizioni di esercizio leggero e per soli 20 minuti sotto sforzo. I valori negli animali sono probabilmente simili. Da quanto
detto, è evidente che devono esistere altri sistemi per garantire la disponibilità di glucosio nei periodi di sforzo o di digiuno
prolungato. Infatti, durante tali condizioni, l'apporto di glucosio è garantito dalla gluconeogenesi. La gluconeogenesi è
stimolata dall'enzima fruttosio 1,6 difosfatasi, attivato tramite fosforilazione. Questo enzima praticamente inverte la via
glicolitica, portando alla produzione di glucosio dalle stesse molecole che sono intermedi della sua demolizione ossidativa.
Sono substrati importanti il piruvato e tutti gli intermedi del ciclo di Krebs.
•
•
•
•
•
•
•
A questo punto, bisogna ricordare che il piruvato e la maggior parte degli intermedi del ciclo di Krebs possono derivare dalla
deaminazione degli aminoacidi. Il momento di entrata dei vari aminoacidi nello schema del metabolismo dei carboidrati è
illustrato nella Figura 31-8. Il piruvato e tutti gli intermedi del ciclo di Krebs possono rientrare attraverso la via ossidativa
(non tutte le reazioni di gluconeogenesi sono l'esatto contrario delle corrispondenti glicolitiche, ma il risultato netto della
gluconeogenesi è il contrario della glicolisi), portando alla produzione di glucosio. Quindi, dagli aminoacidi derivano gran
parte dei precursori per la formazione di glucosio tramite gluconeogenesi. Il risultato finale della stimolazione del glucagone
è la produzione di glucosio tramite glicogenolisi e gluconeogenesi, trasformando il fegato in un organo produttore di
glucosio.
LA MOBILIZZAZIONE DI FONTE ENERGETICA DAI TESSUTI PERIFERICI AVVIENE QUANDO
DIMINUISCONO I LIVELLI EMATICI DI INSULINA
Nella fase successiva all'assorbimento, i pattern metabolici dei tessuti periferici cambiano, per supportare la capacità del
fegato nel mantenere l'apporto di fonte energetica.
IL MUSCOLO REAGISCE ALLA RICHIESTA METABOLICA DI GLUCOSIO MOBILIZZANDO AMINOACIDI
PER SUPPORTARE LA GLUCONEOGENESI EPATICA
La mobilizzazione muscolare di aminoacidi sembra stimolata, per la maggior parte, da una relativa mancanza di insulina, per
cui, essa si verifica quando le concentrazioni ematiche di glucosio sono basse. Gli aminoacidi muscolari mobilizzati
provengono dal pool aminoacidico intracellulare precedentemente descritto. In ogni caso, le reazioni di mobilizzazione sono
complesse e la distribuzione degli aminoacidi che lasciano il muscolo non corrisponde a quella del pool, come sarà descritto
più avanti.
IL RILASCIO MUSCOLARE DI AMINOACIDI È CORRELATO AD UNA RIDOTTA CAPTAZIONE DI
GLUCOSIO ED AMINOACIDI
La diminuzione della concentrazione di insulina nel periodo di post assorbimento ha un doppio effetto sul muscolo:
diminuzione dell'entrata di aminoacidi dal siero nel pool intracellulare e ridotta entrata di glucosio nella cellula muscolare ai
fini energetici. La ridotta disponibilità di aminoacidi stimola la degradazione delle proteine, volta al mantenimento delle
dimensioni del pool intracellulare. La riduzione di glucosio porta ad un maggiore utilizzo degli aminoacidi del pool per la
produzione di energia.
•
•
•
•
•
Il pattern muscolare di utilizzo degli aminoacidi ai fini energetici può sembrare, inizialmente, inutilmente complesso, in
quanto richiede un uso selettivo ed una notevole trasformazione degli aminoacidi. I BCAA sono la principale fonte di energia
per la cellula muscolare durante la fase di post assorbimento, in quanto rappresentano circa un terzo della quota totale di
aminoacidi muscolari. Il catabolismo dei BCAA comincia con la deaminazione e la formazione dei corrispondenti alfa-chetoacidi, questi entrano nel ciclo di Krebs per la produzione di energia. La deaminazione dei BCAA richiede la disponibilità di
accettori per ricevere gli aminogruppi, rappresentati essenzialmente dal piruvato, che darà luogo alla formazione di alanina. Il
piruvato può derivare dal glicogeno muscolare, dal glucosio ematico, o dai prodotti metabolici degli alfa-cheto-acidi dei
BCAA. Quando il metabolismo degli alfa-cheto-acidi dei BCAA diventa fonte di piruvato per la sintesi di alanina, il risultato
finale è la conversione di BCAA in alanina (Fig. 31-13). In sostanza, l'attività metabolica generale del muscolo durante la
fase di post assorbimento è la demolizione dei BCAA e la formazione di alanina. L'alanina neoformata passa dalla cellula
muscolare al sangue e può essere captata dal fegato per la gluconeogenesi.
IL COMPLESSO MECCANISMO DI CATABOLISMO E RILASCIO MUSCOLARE DEGLI AMINOACIDI È
NECESSARIO PER SOPPERIRE ALLA LIMITATA CAPTAZIONE EPATICA DI AMINOACIDI RAMIFICATI E
PER FACILITARE LA RIMOZIONE DI AMINE AZOTATE DAL MUSCOLO
Si potrebbe pensare che per il trasferimento degli aminoacidi al fegato, possa essere sufficiente un sistema più semplice.
Perché gli aminoacidi del pool cellulare muscolare non passano semplicemente in circolo per raggiungere il fegato ed essere
utilizzati per la sintesi di glucosio?
La risposta è legata alla limitata capacità epatica di captazione dei BCAA, e nella necessità di allontanare azoto aminico dal
muscolo. I BCAA, aminoacidi dominanti nel muscolo, non vengono captati facilmente dal fegato per cui, se non venissero
trasformati in alanina, il loro trasferimento a livello epatico sarebbe limitato.
Inoltre, l'alanina è un utile mezzo per trasferire al fegato l'azoto che deriva dalla deaminazione degli aminoacidi muscolari.
Ciò è importante perché, se non rimossi, gli aminogruppi derivanti dal catabolismo degli aminoacidi muscolari potrebbero
portare alla formazione di livelli tossici di ammoniaca. L'ammoniaca viene detossificata tramite la formazione di urea e tale
reazione può avvenire solo nel fegato. Quindi, l'alanina è un precursore glucogenetico che trasporta azoto al fegato per la
sintesi di urea
•
•
•
•
•
La regolazione della mobilizzazione di proteine muscolari è per lo più dovuta alla mancanza di insulina, ma anche il cortisolo
svolge un ruolo importante nel catabolismo delle proteine e nella mobilizzazione di aminoacidi. Tramite la mobilizzazione
delle proteine muscolari e la stimolazione della gluconeogenesi epatica, il cortisolo svolge uno dei suoi effetti principali,
aumentando la concentrazione ematica di glucosio. In condizioni normali, il glucagone, l'altro principale ormone
gluconeogenico, agisce sul fegato e non sembra avere effetti diretti sul muscolo
LA REAZIONE DEL TESSUTO ADIPOSO DURANTE LA FASE SUCCESSIVA ALL'ASSORBIMENTO PORTA
ALLA MOBILIZZAZIONE DI ACIDI GRASSI
Gli acidi grassi vengono rilasciati dal tessuto adiposo per azione dell'enzima stimolato dalla fosforilazione lipasi
ormonosensibile (HSL). Tale enzima viene stimolato dalla relativa mancanza di insulina nel periodo post assorbitivo,
l'insulina, infatti, sopprime l'azione della HSL promuovendo la sua defosforilazione. Il glucagone potrebbe agire a livello del
tessuto adiposo stimolando il catabolismo dei trigliceridi e la fosforilazione, quindi l'attivazione della HSL. Verosimilmente
però, il glucagone agisce solo a livello epatico e la normale stimolazione del HSL avviene ad opera di adrenalina e
noradrenalina; la noradrenalina origina dalle fibre simpatiche del tessuto adiposo. La relazione tra sistema nervoso simpatico
del tessuto adiposo e disponibilità di fonti energetiche non è
chiara, ma le catecolamine ed i neurotrasmettitori sembrano rappresentare lo stimolo primario per il catabolismo dei
trigliceridi. In ogni caso, lo stimolo negativo legato alla mancanza di insulina potrebbe essere il principale sistema di
regolazione della mobilizzazione di grassi dal tessuto adiposo.
La stimolazione della HSL nel periodo post assorbitivi porta al passaggio di acidi grassi dal tessuto adiposo al sangue, dove,
essendo insolubili, si legano all'albumina in maniera reversibile. Gli acidi grassi legati all'albumina vengono definiti acidi
grassi non esterificati (NEFA), per distinguerli dai trigliceridi dei chilomicroni e dalle lipoproteine. I NEFA ematici possono
essere utilizzati direttamente ai fini energetici da molti tessuti, ma la maggior parte di essi viene captata dal fegato ed
utilizzata per la sintesi di corpi chetonici e VLDL, come descritto successivamente.
Utilizzo dei nutrienti durante periodi prolungati di
malnutrizione o di digiuno completo
•
•
•
•
•
DURANTE PERIODI PROLUNGATI DI DIGIUNO O IPONUTRIZIONE, GLUCOSIO ED AMINOACIDI SONO
CONSERVATI GRAZIE ALL'INTENSO UTILIZZO DI GRASSI E CORPI CHETONICI AI FINI ENERGETICI
Dalla precedente descrizione del metabolismo nella fase successiva all'assorbimento, si evince che gli aminoacidi
rappresentano un importante deposito di precursori del glucosio, e substrati per la produzione di energia. Durante digiuno o
iponutrizione prolungata però, l'animale non può attingere dal muscolo per la produzione di glucosio ed energia, in quanto
ciò porterebbe ad uno stato di debolezza legato all'esaurimento delle proteine muscolari.
Per tale motivo, esistono meccanismi volti a proteggere il muscolo durante il periodo di insufficiente apporto energetico. Di
conseguenza, per risparmiare le proteine, l'utilizzo delle fonti energetiche di deposito, si avvale, come substrato metabolico,
non del glucosio ma del tessuto adiposo.
GRAN PARTE DEGLI ACIDI GRASSI RILASCIATI DAL TESSUTO ADIPOSO VIENE CAPTATA
DIRETTAMENTE DAL FEGATO
Durante periodi prolungati di iponutrizione, la bassa disponibilità di glucosio porta alla rapida mobilizzazione di acidi grassi
dal tessuto adiposo sotto forma di acidi grassi non esterificati (NEFA). Anche se i NEFA possono essere utilizzati da molti
tessuti, gran parte di essi arriva al fegato, che riceve la maggior parte del flusso sanguigno, e che possiede un efficace sistema
di captazione dei NEFA dal sangue. Una volta raggiunti gli epatociti, i NEFA possono seguire tre potenziali vie metaboliche.
La prima è la completa ossidazione per la produzione di energia, ma le necessità energetiche del fegato sono tali che solo una
piccola quota degli acidi grassi del tessuto adiposo va incontro a tale reazione. La seconda è l'esterificazione con produzione
di trigliceridi e la terza è la produzione di corpi chetonici. La sintesi di trigliceridi sarà discussa successivamente; di seguito,
viene descritta la produzione di corpi chetonici.
•
•
•
•
•
LA FORMAZIONE EPATICA DI CORPI CHE TONICI È STIMOLATA DALLA BASSA DISPONIBILITÀ DI
GLUCOSIO, UN ELEVATO RAPPORTO GLUCAGONE/INSULINA, ED UN REGOLARE APPORTO DI ACIDI
GRASSI
La formazione di corpi chetonici avviene nei mitocondri dell'epatocita, e la velocità di formazione è controllata dal trasporto
di acidi grassi attraverso la membrana mitocondriale. Gli acidi grassi entrano nel mitocondrio combinati con una molecola
nota come carnitina, ed il trasporto dipende da un enzima denominato carnitina palmitoiltransferasi I (CPT I). L'attività di
questo enzima, con la disponibilità di acidi grassi, è il fattore determinante la velocità di produzione di corpi chetonici.
L'attività della CPT I è regolata in modo interessante, essendo inibita da un intermedio della sintesi degli acidi grassi, la
malonil CoA. Le concentrazioni di malonil CoA sono elevate quando è presente un eccesso di glucosio, quindi, quando il
fegato sta rispondendo all'insulina ed il glucosio viene utilizzato per la sintesi di acidi grassi. Quando l'apporto di glucosio è
scarso, o le concentrazioni di glucagone sono relativamente alte rispetto ai livelli di insulina, vengono sintetizzati pochi acidi
grassi a livello epatico. Ne deriva che le concentrazioni di malonil CoA sono basse e la CPT I è completamente attiva quando
il rapporto insulina/glucagone è basso.
Quando la CPT I è attiva, una maggior quantità di acidi grassi raggiunge il fegato per la sintesi di corpi chetonici. Questo
sistema di regolazione, ben orchestrato ma complesso, è necessario, in quanto il fegato può sia produrre sia consumare acidi
grassi. Se non ci fosse un modo per interrompere la degradazione degli acidi grassi durante la loro sintesi, si verificherebbe
un inutile ciclo di produzione e degradazione. L'inibizione della CPT I da parte del malonil CoA crea un sistema che blocca il
catabolismo degli acidi grassi neosintetizzati, ma permette l'utilizzo degli acidi grassi derivati dal tessuto adiposo nei periodi
di insufficiente apporto energetico. Il risultato finale è una relazione reciproca tra disponibilità di glucosio e produzione di
acidi grassi. Anche se prodotti dal fegato, i corpi chetonici non possono essere utilizzati in questa sede per la produzione di
energia, per cui essi devono raggiungere i tessuti periferici per l'utilizzo. Se la concentrazione di corpi chetonici nel sangue
diviene troppo alta, parte viene escreta con le urine.
IL GLUCAGONE SVOLGE UN RUOLO IMPORTANTE NELL'IPERPRODUZIONE DI CORPI CHETONICI
DURANTE IL DIABETE MELLITO
Se non trattato, il diabete mellito negli animali, e specialmente nel cane, porta ad un aumento delle concentrazioni di corpi
chetonici nel sangue. Il diabete mellito si verifica per mancanza di insulina, ma la produzione epatica di corpi chetonici
avviene per la notevole azione del glucagone. Anche se le concentrazioni di glucosio nel diabete sono alte, la mancanza di
insulina porta ad un basso rapporto insulina/glucagone, per cui il fegato viene stimolato unicamente dal glucagone. Il
glucagone inibisce la produzione di acidi grassi a partire dal glucosio, per cui le concentrazioni di malonil CoA sono basse e
l'attività della CPT I è alta. Inoltre, per la mancanza di insulina che sopprime la HSL del tessuto adiposo, la concentrazione di
NEFA nel sangue è elevata. La combinazione di alta disponibilità di NEFA ed attività elevata di CPT I inducono un rapido
trasporto di acidi grassi ai mitocondri con notevole produzione di corpi chetonici, anche se le concentrazioni ematiche di
glucosio sono alte.
•
•
•
•
•
•
•
•
GLI ACIDI GRASSI NON POSSONO ESSERE UTILIZZATI PER LA SINTESI DI GLUCOSIO
È importante comprendere che il metabolismo dei grassi all'interno dei mitocondri non può contribuire direttamente alla
gluconeogenesi. Una volta attraversata la membrana mitocondriale, gli acidi grassi vanno incontro a beta-ossidazione, che
porta alla successiva rimozione di due unità di carbonio dell'acetil CoA dalle catene carboniose degli acidi grassi. L'acetil
CoA può quindi entrare nel ciclo di Krebs tramite condensazione con ossalacetato. Dal momento che tutti gli intermedi del
ciclo di Krebs possono portare alla formazione di glucosio, si potrebbe pensare che ciò avvenga anche per l'acetil CoA
derivato dalla beta-ossidazione degli acidi grassi. Non si ha in questo caso una netta produzione di ossalacetato associata con
il consumo di CoA nel ciclo di Krebs. L'ossalacetato preesistente si combina con l'acetil CoA per formare citrato all'inizio del
ciclo. Alla fine del ciclo, l'ossalacetato si riforma, ed i due carbonio dell'acetil CoA vengono convcrtiti in anidride carbonica.
Questo processo non può produrre nuovo ossalacetato.
I CORPI CHETONICI SI FORMANO NEI MITOCONDRI A PARTIRE DALL'ACETIL COA
Non tutto l'acetil CoA mitocondriale deve entrare nel ciclo di Krebs. Infatti, quando gli acidi grassi entrano rapidamente nei
mitocondri, c'è più acetil CoA di quello necessario per il ciclo di Krebs, ed è da questo acetil CoA in eccesso che si originano
i corpi chetonici.
I corpi chetonici possono lasciare i mitocondri liberamente.
I corpi chetonici influenzano l'omeostasi della fonte energetica nei tessuti periferici, dove rappresentano un sostituto del
glucosio. In tal modo, si mantiene la disponibilità di glucosio riducendo la necessità di gluconeogenesi.
LE LIPOPROTEINE EPATICHE A BASSISSIMA DENSITÀ POSSONO ESSERE SINTETIZZATE A PARTIRE SIA
DAGLI ACIDI GRASSI DEL TESSUTO ADIPOSO DI DEPOSITO, SIA DA QUELLI NEOFORMATI
Nella sezione sul metabolismo della fase di assorbimento è stata descritta la produzione epatica di VLDL. Durante la fase di
assorbimento, i trigliceridi per la sintesi di VLDL derivano dagli acidi grassi sintetizzati a partire dal glucosio. Durante i
processi catabolici le VLDL possono essere ancora prodotte, ma gli acidi grassi derivano dai NEFA (vedi Fig. 31-5). Questo
può apparentemente sembrare un passaggio inutile ed insufficiente. Perché gli acidi grassi del tessuto adiposo devono essere
trasportati al fegato per la produzione di VLDL se possono essere direttamente metabolizzati dai tessuti per produrre energia?
La sintesi di VLDL è necessaria per garantire un miglior sistema di trasporto. La capacità di trasporto serica dei NEFA è
limitata in quanto essi viaggiano legati all'albumina, e la disponibilità di quest'ultima nel legare i NERA è insufficiente nei
periodi di rapida mobilizzazione dei grassi, in quanto saturata. Le VLDL rappresentano un sistema di trasporto degli acidi
grassi che, non dipendendo dall'albumina, è illimitato rispetto a quello esercitato dall'albumina.
•
•
•
•
L'ASSETTO ORMONALE REGOLA LA DISTRIBUZIONE ORGANICA DEGLI ACIDI GRASSI DELLE
LIPOPROTEINE A BASSISSIMA DENSITÀ
Durante la fase di assorbimento, le VLDL sono dirette al tessuto adiposo per l'azione della LPL, enzima stimolato
dall'insulina. Le LPL sono presenti anche nel tessuto muscolare, ma non sono insuline dipendenti. Quindi, durante i periodi di
bassa disponibilità di glucosio, la LPL del tessuto adiposo è inibita per la mancanza di insulina, mentre quella del muscolo è
pienamente attiva. Tale situazione porta selettivamente gli acidi grassi delle VLDL verso il tessuto muscolare nei periodi di
mobilizzazione del tessuto adiposo.
VARIAZIONI DI CONCENTRAZIONE DELL'ORMONE DELLA CRESCITA POSSONO RICHIEDERE, PER
L'UTILIZZO DELLA FONTE ENERGETICA PERIFERICA, CORPI CHETONICI ED ACIDI GRASSI, AL POSTO
DI GLUCOSIO E AMINOACIDI
Le variazioni del metabolismo epatico, indotte dalla mobilizzazione di grasso, riescono a risparmiare le proteine solo per i
cambiamenti di utilizzo di glucosio ed aminoacidi nei tessuti periferici. Nel momento in cui corpi chetonici, NEFA e
trigliceridi delle VLDL diventano la principale fonte di energia, c'è minore richiesta di glucosio ed aminoacidi come substrati
energetici. A livello tissutale, variazioni endocrine, oltre ai bassi livelli di insulina, possono favorire questo cambiamento di
utilizzo dei substrati a livello periferico. In molte specie, le concentrazioni di ormone della crescita aumentano durante
periodi prolungati di scarso apporto energetico. L'ormone della crescita è un antagonista dell'insulina, quindi promuove un
aumento delle concentrazioni di glucosio serico anche in presenza di livelli normali o quasi normali di insulina. Inoltre,
l'ormone della crescita può avere effetti diretti nel conservare proteine e mobilizzare lipidi.
Considerazione sulla peculiare fonte
energetica dei ruminanti
•
•
•
•
I RUMINANTI PRESENTANO UNO STATO COSTANTE DI GLUCONEOGENESI LEGATO AL LORO
PARTICOLARE PROCESSO DIGESTIVO
Nei ruminanti, la maggior parte della digestione dei carboidrati avviene nei prestomaci tramite digestione fermentativa. Il
risultato è che quasi tutti i carboidrati non raggiungono l'intestino per la digestione ghiandolare e l'assorbimento sotto forma
di glucosio. Di conseguenza, i ruminanti vivono in un costante stato di potenziale deficienza di glucosio. Per fronteggiare tale
situazione, il ruminante ha sviluppato un sistema efficiente per la conservazione e la produzione di glucosio.
Praticamente, nei ruminanti tutto il glucosio disponibile deriva dalla gluconeogenesi. Quantitativamente, il principale
precursore del glucosio è l'acido grasso volatile propionato. Il propionato contribuisce alla sintesi di glucosio entrando nel
ciclo di Krebs a livello di succinato. È da sottolineare che il succinato è un intermedio del ciclo di Krebs a quattro carbonio e
può portare alla formazione netta di ossalacetato, il primo metabolita della gluconeogenesi. Anche gli altri acidi grassi
volatili, acetato e butirrato, entrano nel ciclo di Krebs ma sotto forma di acetil CoA, come gli acidi grassi a catena lunga del
tessuto adiposo. Come già detto precedentemente, l'acetil CoA non può portare ne alla formazione di ossalacetato ne di
glucosio. Quindi, tra i principali substrati energetici dei ruminanti, solo il propionato può portare alla formazione di glucosio.
Quasi tutto il propionato assorbito dal rumine giunge per via portale al fegato, senza mai entrare nel circolo sistemico, ciò fa
si che tutto il propionato venga utilizzato per la gluconeogenesi. Oltre al propionato, anche gli aminoacidi assorbiti a livello
intestinaie rappresentano un substrato per la gluconeogenesi.
L'uso degli aminoacidi nella dieta per la gluconeogenesi è stato discusso nella sezione del metabolismo della fase di
assorbimento in relazione al rilascio di aminoacidi stimolato dal glucagone.
•
•
•
Oltre alla costante gluconeogenesi, i ruminanti fanno fronte alle proprie necessità di glucosio mediante la sua conservazione.
Gli acidi grassi, che in alcuni animali quali primati, ratti e cani vengono sintetizzati nel fegato, nei ruminanti lo sono
esclusivamente a livello del tessuto adiposo. Inoltre, il glucosio non viene utilizzato per la sintesi di acidi grassi, che derivano
essenzialmente dall'acetato, il principale substrato energetico dei ruminanti. Il tessuto adiposo utilizza il glucosio solo per la
sintesi del glicerolo per i trigliceridi. Durante la lattazione, gli acidi grassi prodotti nella mammella per il grasso del latte
derivano dall'acetato o dai corpi chetonici, ma mai dal glucosio.
Alcune importanti patologie metaboliche dei ruminanti si verificano quando l'omeostasi del glucosio è sotto stress. Le bovine
sono particolarmente vulnerabili durante il picco di lattazione perché la sintesi del lattosio, zucchero del latte, richiede
glucosio. Nelle bovine ad alta produzione, quasi tutto il glucosio prodotto viene utilizzato per la sintesi di lattosio e le
funzioni tissutali devono
utilizzare substrati energetici alternativi. La pecora presenta una simile situazione di stress da sintesi di glucosio alla fine
della gestazione. Le esigenze energetiche di feto e placenta possono essere soddisfatte solo dal glucosio (o lattato derivato dal
glucosio) e dagli aminoacidi. La pecora, rispetto a molti altri animali, ha un alto rapporto tra massa fetale e corporea, per cui i
meccanismi omeostatici energetici sono particolarmente intensi durante la gravidanza. In queste circostanze, il sistema
omeostatico del glucosio spesso è alterato provocando condizioni patologiche quali la chetosi nella bovina in lattazione e la
tossiemia gravidica nella pecora.