Questo articolo di Cornish-Bowden , che ho riprodotto solo in parte , contiene una
analisi dei risultati ottenuti da Harden e Yuong nello studio della glicolisi che
vengono spiegati secondo le attuali conoscenze . Così le conseguenze dell'aggiunta di
fosfati , l'accumulo di esoso fosfati , l'effetto dell'aggiunta di arseniati , che furono
investigati da Harden e Young nel succo di lievito o sistema a cell-free , sono chiariti
in modo sintetico .
Reprinted from:
New Beer in an Old Bottle: Eduard Buchner and the Growth of Biochemical Knowledgeed.
A. Cornish-Bowden,
Universitat de València, Valencia, Spain, 1997
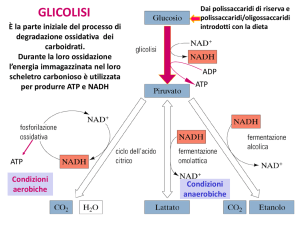
La glicolisi in un sistema di cell-free
[Ndt: per sistema a cell-free si intende l'estratto di lievito privato delle sue cellule ]
La scoperta di Eduard Buchner del metabolismo di un sistema a cell - free cento anni
fa è considerata una pietra miliare della biochimica ; si può dire che rappresentò il
punto di partenza per uno studio dettagliato del metabolismo . Il fatto che una
frazione solubile del lievito , che egli chiamò "zimase" , fosse capace di produrre
etanolo dal glucosio rappresentò un evento rivoluzionario e alla fine liquidò l'idea
che le cellule vive fossero essenziali alla fermentazione ( Buchner , 1897). Più tardi
venne riconosciuto che questo estratto contenente proteine era una miscela
complessa che conteneva enzimi responsabili del processo che noi ora chiamiamo
glicolisi , e durante il successivo mezzo secolo furono definiti tutti i metaboliti e gli
enzimi della glicolisi. Questa biochimica classica che utilizza un estratto di cell-free
per catalizzare un processo biologico è stata alla base di molte dei miei lavori di
ricerca compiuti durante i trenta anni passati . Uno delle caratteristiche degli
esperimenti di Buchner era che il sistema di cell-free non produceva mai qualcosa di
uguale alla trasformazione dello zucchero ad alcol che poteva realizzarsi con le
cellule di lievito intere . Persino il suo estratto molto concentrato , equivalente ad un
denso impasto di cellule di lievito , produceva diossido di carbonio ed alcol ad una
velocità che noi possiamo stimare meno dell'1% della potenziale attività enzimatica.
Tipicamente nella produzione del vino , il liquido di partenza contiene circa il 20% in
peso di zucchero , che è convertito completamente a etanolo, CO2 , biomassa di
cellule di lievito e tutti gli importanti componenti aromatici minori . Ma con i primi
esperimenti con estratti di lievito , soltanto una piccola quantità dello zucchero
addizionato poteva essere convertito in etanolo prima che il processo si fermasse
dopo un giorno o due . Una delle principali ragioni è che il controllo dell'entrata del
glucosio nella via glicolitica è , almeno in parte, stabilito dalla membrana della
cellula del lievito , la quale naturalmente è perduta nel fare l'estratto cellulare . Noi
possiamo ora valutare che il controllo del metabolismo era dovuto principalmente
alla fosforilazione incontrollata del glucosio, e lo sbilanciamento dei metaboliti che
questo causa . Benchè non fossero in grado di spiegare i fatti in questo modo ,
Harden e Young nel 1906 riconobbero effettivamente la stessa cosa quando
scoprirono l'accumulo di esoso fosfati nell'estratto di lievito in fermentazione ........
Addizionando glucosio a un sistema glicolitico ricostruito del lievito abbiamo una
fosforilazione del glucosio non controllata , e un accumulo di esoso fosfati . Questo
effettivamente ripete gli esperimenti di Harden e Young quasi 80 anni fa , i quali
furono capaci di individuare l'esoso difosfato come un intermedio della glicolisi ,
sebbene fosse stato pienamente identificato come fruttosio 1,6-difosfato ben venti
anni più tardi . Che cosa era successo può essere spiegato in modo semplice .
L'esochinasi [ Ndt: enzima che catalizza la rima reazione della glicolisi ] è presente
nell'estratto di lievito piuttosto ad alti livelli e non ha un significante controllo
allosterico . Così addizionando glucosio questi è fosforilato velocemente , con
rapido aumento dei livelli di glucosio /fruttosio 6-fosfato . Viene rilevata anche una
inibizione della fosfofruttocinasi , con conseguente accumulo di fruttosio difosfato .
La glicolisi è portata a termine , con la formazione di due ADP dalle due esoso
chinasi e la risintesi da parte della fosfogliceratocinasi e piruvato cinasi . Ma la
risintesi produce quattro molecole di ATP , in confronto alle due che sono state
usate dalla esoso cinasi . In assenza di qualsiasi ATPase (Negli esperimenti di
Buchner e di Harden e Young la maggior parte della ATPase era stata rimossa con il
materiale particolato cellulare ) , questo nuovo ATP formato è utilizzato ancora in
un ulteriore fosforilazione del glucosio .
[Ndt: L'ATPase sono una classe di enzimi che catalizzano la decomposizione di ATP in ADP e fosfato
Pi . Questa reazione rilascia energia che viene utilizzata per svariati processi , in particolate per
movimenti trans membrana di molecole e ioni ]
Come risultato abbiamo un netto accumulo di intermedi fosforilati . Sono costituiti
principalmente da esoso fosfati e questi esteri fosforici si formano a spese del
fosfato inorganico che decresce rapidamente . Quando il fosfato inorganico si
avvicina a zero , non c'è più nessuna attività della gliceraldeide fosfato deidrogenasi,
così non può formarsi altro ATP . Si ha una mancanza finale di ATP , e il metabolismo
di fatto si ferma perché non c'è ne fosfato inorganico ne ATP. Harden e Yuong
furono capaci di far ripartire la glicolisi aggiungendo un eccesso di fosfato , il quale
viene tutto incorporato negli esoso fosfati . La reazione netta ha come risultato una
molecola di diossido di carbonio e una di etanolo prodotte per ogni ione fosfato
utilizzato , una stechiometria che essi hanno riconosciuto 80 anni prima dei nostri
studi .
Net:
Glc + 2ADP + 2Pi →2C2H5OH + 2CO2 + 2ATP
Glc + 2ATP →Fru-1,6-P2 + 2ADP
------------------------------------------------------------2Glc + 2Pi →Fru-1,6-P2 + 2C2H5OH + 2CO2
( In questa equazione la specie indicata come fruttosio 1,6-difosfato comprende le
quantità dei trioso fosfati in equilibrio con esso nella reazione catalizzata dalla
aldolasi e trioso fosfato isomerasi ).
Harden e Young scoprirono inoltre che sostituendo il fosfato con l'arseniato , invece
di avvelenare il sistema come ci si poteva aspettare , in realtà era stimolata la
glicolisi . Essi conclusero correttamente che l'arseniato prendeva il posto del
fosfato, e che l' arsenio-estere instabile che si formava si decomponeva , con
l'effetto netto di mantenere disponibile sufficiente fosfato inorganico che era
richiesto per il processo completo della fermentazione . Noi ora interpretiamo
l'effetto dell'arseniato come una attività pseudo -ATPase la quale non solo baypassa
il sequestro di fosfato inorganico , ma elimina anche ogni netta sintesi di ATP .
Questo è dovuto alla decomposizione rapida e spontanea dell'1-arsenio-3fosfoglicerato formata dalla gliceraldeide fosfato deidrogenasi , che abbiamo
stimato accadde con un tempo di vita medio che non supera pochi millisecondi .
Allo scopo di ottenere una glicolisi del sistema a cell-free che si protragga per lungo
periodo è chiaro che dobbiamo sia bilanciare in modo accurato l'attività della
ATPase con una alimentazione accurata di glucosio o trovare una via alternativa per
la decomposizione dell'ATP . L'esperimento con l'arseniato fu la soluzione . Usando
arseniato non fu necessario aggiungere alcun enzima ATPase . Fu possibile
conseguire una prolungata fermentazione , con la completa conversione del glucosio
in etanolo . Noi riuscimmo ad ottenere la conversione di glucosio 1 mole a 2 moli di
etanolo più CO2 in 7 ore . La fermentazione non fu "naturale", poiché si verificò ad
una concentrazione di ATP molto bassa dovuto alla efficienza con la quale il sistema
composto da gliceraldeide fosfato deidrogenasi e fosfoglicerato cinasi abbatteva
l'ATP in presenza di arseniato . Ma il livello costante di ATP di poco più di 0,1 mM fu
sufficiente a far girare al massimo la glicolisi ( questo è probabilmente l'ordine di
concentrazione di ATP che esisteva nell'esperimento di Buchner ) . La via glicolitica ,
precisamente la via di Embden- Meyerhof , è stata al centro di molti studi di
biochimica . Fu la prima via metabolica studiata , venne chiarita pienamente nei
termini delle attuali reazioni catalizzate dal 1940. Ma dopo di allora ciascun enzima
venne studiato in tutti i dettagli e il controllo del sistema continuamente investigato.
E' possibile simulare il processo in un computer , ma una soddisfacente simulazione
della glicolisi non è stata ancora realizzata perché i parametri esatti di ciascun
enzima non sono conosciuti sufficientemente in dettaglio in condizioni fisiologiche .
E' sempre possibile aggiustare i parametri per aver al risposta corretta , ma questo è
difficilmente convincente . Un problema è che la cinetica convenzionale di Michaelis
- Menten dovrebbe essere modificata per tener conto del fatto che in molti casi la
concentrazione degli enzimi è dello stesso ordine di quella del substrato , o anche di
più alta . L'assunzione iniziale da cui viene derivata l'equazione di Michaelis Menten , che la concentrazione del substrato è molto più grande di quella dell'
enzima , non è valida . Molti dei metaboliti nella glicolisi cellulare a ogni istante
sono per lo più legati all'enzima che liberi nel citoplasma . Un secondo problema è
che la concentrazione degli enzimi in vivo può essere 1000 volte più alta di quella
che abbiamo in vitro negli studi di cinetica , e l'estrapolazione della cinetica in vitro
al di sopra di tre ordini di grandezza non è sempre valida . Per ultimo gli esperimenti
riportati in letteratura per determinare i parametri della cinetica sono raramente
eseguiti a pH fisiologico , forza ionica o anche la temperatura , così che ciascun
dato deve essere considerato con prudenza .