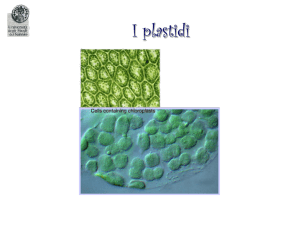
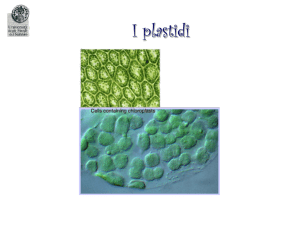
PLASTIDI
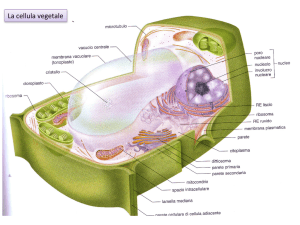
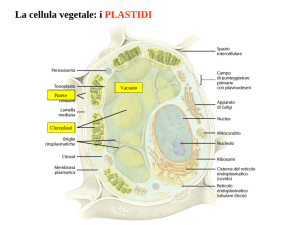
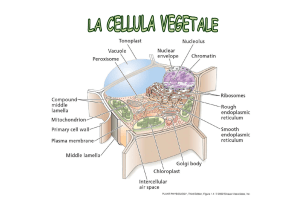
I PLASTIDI, insieme con i vacuoli e la parete
cellulare, sono gli organelli caratteristici delle
cellule vegetali e sono coinvolti nei processi di
fotosintesi e di accumulo.
I plastidi dei tessuti maturi derivano –per trasformazione di
altri tipi di plastidi o direttamente - da piccoli organelli
indifferenziati definiti proplastidi
presenti nelle cellule meristematiche
di 0,5-1 mm
Nei meristemi il numero di proplastidi per cellula è costante
attorno a 10-20.
Prima di ogni divisione cellulare vi è la
duplicazione dei proplastidi
attraverso un meccanismo di scissione binaria
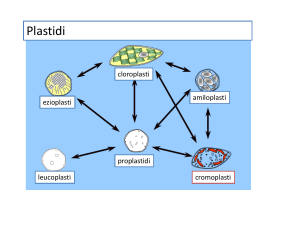
Tipi di plastidi
Cloroplasti: svolgono la fotosintesi
Cromoplasti: contengono grandi quantità di pigmenti
lipidici; si trovano nei fiori e nei frutti
Amiloplasti: accumulano amido
Ezioplasti: uno stadio specifico della trasformazione dei
proplastidi in cloroplasti; si formano in tessuti determinati
a divenire fotosintetici che crescono in assenza di luce
Leucoplasti: termine descrittivo, indica i plastidi incolori
Proplastidi: plastidi piccoli indifferenziati
Differenziamento dei plastidi
Il tipo, il numero e le dimensioni dei plastidi di una cellula
matura dipendono dallo specifico programma di sviluppo della
cellula nel tessuto a cui appartiene
Ad es. nell’ epicarpo (buccia ) dei frutti maturi di pomodoro,
la popolazione di cromoplasti è molto più numerosa (oltre
1000/cellula) di quella delle sottostanti cellule del mesocarpo
Il numero dei cromoplasti è più alto di quello dei cloroplasti da
cui derivano, dimostrando che i plastidi possono dividersi anche
in fase adulta
• I plastidi sono capaci di interconversione,
cioè un tipo può convertirsi in un altro tipo,
es. cloroplasti in ezioplasti.
CROMOPLASTI
I cromoplasti sono plastidi provvisti di pigmenti. Sono
di forma variabile, sono privi di clorofilla ma
sintetizzano e accumulano pigmenti carotenoidi,
questi ultimi sono per lo più sciolti in gocce di grasso
(es. pomodori e peperoni maturi).
Conferiscono il colore giallo, arancione o rosso ai
petali di alcuni fiori, alle foglie senescenti
(in
alternativa alle antociane).
Possono differenziarsi da cloroplasti preesistenti, che
invecchiando perdono la clorofilla, degradano i
tilacoidi
e
contemporaneamente
accumulano
carotenoidi (es. durante la maturazione dei frutti).
I cromoplasti a seconda delle modalità di
sequestro dei carotenoidi
sono differenti e si classificano:
• Globulari
(es. fittone
della carota)
• Fibrillari
(es. petali di
alcuni fiori)
• Membranosi
(es.
frutto del peperone)
• Cristallini (es. frutto
del pomodoro)
Proplastidio
Cloroplasto
Amilo-Leucoplasto
Cromoplasto
• Localizzazione: frutti, petali e parenchima di riserva della carota
• Stimoli: Ormonali, ambientali (temperatura, luce, stress)
• Differenziamento: organogenesi
• Funzione: vessillare
• Contenuto: pigmenti, proteine e lipidi, accumulo di carotenoidi con
l’attivazione di geni nucleari
Leucoplasti (plastidi incolori)
Possono essere specializzati per biosintesi particolari,
come ad es. biosintesi di acidi grassi e metaboliti secondari
come gli oli essenziali
I leucoplasti possono contenere corpi proteici
GLI AMILOPLASTI
ACCUMULO DI AMIDO
Amido in cellule parenchimatiche
(tubero della patata)
Granuli
di
amido
in
cellule
del
parenchima
midollare di
tabacco
Amido = polimero dell’ alfa glucosio
Amilosio (ha struttura
elicoidale) e le unità di
glucosio sono unite
mediante ponti alfa 1-4
I granuli negli amiloplasti sono di amido secondario
o di riserva
Quando tutto il PLASTIDIO si è riempito di
amido e resta solo la membrana delimitante
il plastidio originario si chiamerà
GRANULO D’AMIDO
Si può trovare nelle radici, tuberi, rizomi,
semi, midollo del fusto
Granuli d’amido
Incluso solido, si accumula
sotto forma di grossi granuli
di forma varia (sferica,
ovoidale, poliedrica,
lenticolare, a bastoncino con
dimensioni che variano da 1 a
170 mm)
L ’ILO è il punto di aggregazione
dell’ amido
I granuli sono semplici o
composti
La forma del granulo ha
significato tassonomico
I granuli di amido nelle cellule della cuffia radicale funzionano da
statoliti (apparato di percezione della gravità necessario per la crescita
della radice in profondità nel suolo)
Gli ezioplasti sono
plastidi con corpi
prolamellari. In seguito ad esposizione alla luce
i corpi prolamellari si trasformano in tilacoidi e
l’ezioplasto in cloroplasto
Anche gli ezioplasti sono capaci di divisione
binaria
CLOROPLASTO
plastidio specializzato per il processo fotosintetico
provvisto di:
“Doppia” membrana
esterna
Stroma
Membrane
tilacoidali
Le dimensioni sono di circa 5-10 mm con forma ovoidale (nelle
piante superiori)
La pricipale funzione dei cloroplasti è la
FOTOSINTESI
Per svolgere al meglio questa funzione i cloroplasti si
dispongono, in genere con la superficie più ampia
parallella alla parete cellulare. Tuttavia, all’interno della
cellula, sotto l’influenza della luce possono riorientarsi.
Ontogenesi del cloroplasto
•Da un proplastidio
•Da un leucoplasto
•Da un cloroplasto preesistente
•Da un ezioplasto posto alla luce
Luce e biogenesi del cloroplasto
La luce è il fattore ambientale più importante per
il differenziamento dei
cloroplasti
Agisce sull’espressione genica nucleare e plastidica
Il precursore della clorofilla, la protoclorofillide viene
convertita in clorofilla grazie ad un enzima attivato dalla luce
L’importanza della luce risulta evidente quando un germoglio od
una coltura cellulare vengono mantenute al buio. Dai proplastidi
si differenziano gli ezioplasti
La divisione dei proplastidi è seguita da una fase di
crescita dimensionale dell’organello, fino al valore
necessario per la successiva divisione
Il cloroplasto è dotato di DNA di tipo procariotico
e durante la sua fase di crescita si ha anche la
replicazione di questo DNA
Alla prima fase di divisione dei proplastidi segue una seconda
fase di divisioni degli organelli in via di differenziamento o
differenziati
Questa fase di divisione porta ad un aumento del numero di
plastidi per cellula ed è correlata con il differenziamento
dell’organello proprio del tipo cellulare
Da una cellula del meristema apicale del germoglio si
potrà differenziare una cellula fotosintetica con
100 e più cloroplasti.
Da una cellula del meristema apicale della radice si
potrà differenziare una cellula parenchimatica con
un numero molto inferiore di amiloplasti.
Le membrane plastidiali sono caratterizzate da
acidi grassi con un elevato grado di insaturazione e da
un alto contenuto in galattolipidi
L’unico fosfolipide è il fosfatidilglicerolo, questo nella
membrana esterna è sostituito dalla fosfatidilcolina
Vi sono poi lipidi apolari, principalmente composti
ossidoriduttivi come i chinoni e tocoferoli
Nelle membrane dei tilacoidi sono inclusi le clorofille
ed i pigmenti carotenoidi (di natura liposolubile)
responsabili della cattura della luce che dà il via
al processo fotosintetico (fase luminosa).
Nello stroma, invece, sono presenti gli enzimi che
catalizzano la fase oscura della fotosintesi.
Un pigmento è una sostanza che assorbe luce
selettivamente
Un pigmento ha il colore della luce che riflette e
trasmette.
Es. la clorofilla dà il colore verde alle foglie
perché assorbe luce nel VIOLETTO, AZZURRO e
ROSSO, però riflette e trasmette la luce VERDE
Tutti i pigmenti sono molecole organiche non
polimeriche, con una serie di doppi legami alternati.
Stroma
Sede del DNA (di tipo procariotico), ribosomi 70S ed
il corredo enzimatico per vari metabolismi,
quali per es. il processo fotosintetico,
per la biosintesi di carotenoidi, degli acidi grassi
Granuli d’amido
Plastoglobuli (costituiti di chinoni, carotenoidi, galattolipidi e
scarse proteine)
Nelle prime fasi del differenziamento le membrane tilacoidali
sono di tipo stromatico
Successivamente ai nuclei centrali dei 2 fotosistemi, vengono
sintetizzati i complessi antenna LHC dei fotosistemi I e II.
Parallelamente è necessaria la sintesi di pigmenti che legandosi
alle proteine le stabilizzano
In questa fase avanzata del differenziamento dei cloroplasti
iniziano a formarsi i grana con la progressiva produzione di
membrane tilacoidali e l’aumento del numero e dimensioni
degli impilamenti granali
La pricipale funzione dei cloroplasti è la
FOTOSINTESI
Per svolgere al meglio questa funzione i cloroplasti
si dispongono, in genere con la superficie più ampia
parallella alla parete cellulare. Tuttavia, all’interno
della cellula, sotto l’influenza della luce possono
riorientarsi.
Nelle membrane dei tilacoidi sono inclusi le
clorofille ed i pigmenti carotenoidi (di natura
liposolubile) responsabili della cattura della luce
che dà il via al processo fotosintetico (fase
luminosa).
Nello stroma, invece, sono presenti gli enzimi che
catalizzano la fase oscura della fotosintesi.
Nelle membrane tilacoidali sono presenti due tipi di
COMPLESSI ANTENNA, detti
FOTOSISTEMI I e II
Cosa sono ???
complessi di natura proteica associati ai
pigmenti fotosintetici, essenziali o accessori
quali clorofille e carotinoidi o altri pigmenti nelle alghe
La clorofilla a è il pigmento essenziale degli eucarioti
fotosintetici e dei cianobatteri
A che servono I FOTOSISTEMI???
I fotosistemi catturano l’energia luminosa.
I plastidi delle Alghe
I plastidi delle alghe hanno forme diverse
I tilacoidi possono essere isolati e paralleli come nelle rodofite
e dotati di ficobilisomi
I ficobilisomi contengono ficoeritrina, ficocianina,
alloficocianina sono pigmenti necessari per far vivere queste
alghe nella profondità del mare (fino al limite della zona
eufotica, circa -200 m in acque pulite)
Spesso i tilacoidi decorrono uniti a 2, 3 o 4. Nelle alghe eteroconte è
presente un tilacoide circolare parallelo alla membrana interna del
cloroplasto (lamella periferica)
Nelle alghe verdi compare l’ultrastruttura granale ed intergranale.
Teoria endosimbiontica
• E’ dell’inizio del ‘900 e fu ripresa alla fine degli
anni 60 del novecento, è basata
sull’osservazione che mitocondri e cloroplasti
posseggono DNA proprio e di tipo batterico e
ribosomi 70S, come i procarioti, e sono sensibili
agli stessi antibiotici che interferiscono con i
ribosomi dei procarioti.
• Ad oggi questa teoria è largamete accettata
L’evento endosimbiontico all’origne
dei mitocondri sembra avvenuto
per primo
L’origine del cloroplasto sarebbe
dovuta ad un evento successivo tra
un eucariote capace di fagocitosi ed
un cianobatterio
• Il cianobatterio inglobato NON venne
digerito, come si trasformò in
cloroplasto??
• Perse la parete cellulare e trasferì più del
90 del suo genoma al nucleo dell’ospite.
• Comparvero nel genoma residuo i geni per
l’rRNA del cloroplasto
• Da questo evento di endosimbiosi
nacquero le prime cellule fotosintetiche
con cloroplasto avvolto da due membrane,
la più interna corrispondente alla
membrana citoplasmatica del
cianobatterio e la più esterna alla
membrana del vacuolo digestivo del
fagotrofo
E tutte le altre divisioni algali???
Endosimbiosi secondaria
• Alghe come le euglene, le dinophyta, le
eteroconte, ecc., avrebbero acquisito i
cloroplasti per una simbiosi secondaria fra
un protozoo eterotrofo ed un’alga
eucariotica rossa o verde
E le mebrane delimitanti il
cloroplasto quante diventano???
• L’alga eucariote di solito presenta 4
membrane, due membrane RE (una,
l’interna, il plasmalemma originario
dell’alga entrata in simbiosi, l’altra,
l’esterna, membrana vacuolare dell’ospite
fagotrogo) e le due membrane del
cloroplasto, residuo dell’evento
endosimbiontico primario
Nelle euglene e nelle dinofite le
membrane che circondano il
cloroplasto sono tre, perchè??
Una delle membrane RE è stata
persa
La TEORIA ENDOSIMBIONTICA è sostenuta da
1. Presenza di DNA non organizzato in cromosomi (circolare a doppia elica ),
non compartimentato ed in grado di duplicarsi
2. Presenza di ribosomi che effettuano sintesi proteica, con caratteristiche
simili a quelle dei procarioti ( 70 S e un rRNA 16S con sequenze omologhe
in plastidi e cianobatteri)
3. Divisione per scissione binaria
4. Omologia di alcune proteine, ad es.:
4a. Presenza di una RNA-polimerasi (PERP, Plastidial Encoded RNA
Polymerase) con un alto grado di omologia con quella dei cianobatteri
5. In alcune alghe i cloroplasti sono circondati da una o più altre membrane,
probabile “relitto” del fenomeno simbiontico)
Cooperazione genoma
nucleare e
plastidiale porta ad
es.alla formazione
della RUBISCO,
l’enzima essenziale
per la fissazione
della CO2