Comunicazione Cellulare
S. Beninati
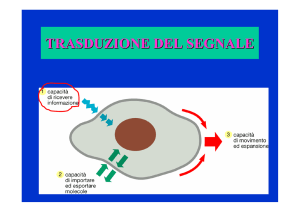
Trasduzione del segnale
in tutti i metazoi o organismi pluricellulari,
una complessa rete di comunicazione tra
cellule
coordina
la
crescita,
il
differenziamento e il metabolismo.
In alcuni casi la comunicazione cellula-cellula e'
diretta (vedi le giunzioni serrate o comunicanti
che consentono a cellule adiacenti di
scambiarsi piccole molecole).
Le cellule devono poter comunicare anche a distanze
superiori di quelle che possono essere coperte da
catene di contatti cellula-cellula. I questi casi i segnali
sono costituiti da prodotti extracellulari. Queste
sostanze sono prodotte e liberate da cellule adibite alla
comunicazione, dette cellule segnale.
I loro prodotti sono liberati nello spazio extracellure nel quale
diffondono liberamente raggiungendo distanze elevate.
La
comunicazione
o
trasduzione
intracellulare del segnale è la catena di
reazioni che si attuano quando i segnali emessi
da cellule dette segnale, interagiscono tramite
recettori proteici della superficie cellulare o del
nucleo, di cellule dette bersaglio di vario tipo.
Tali segnali attivano o disattivano l'espressione
genica di fattori di trascrizione proteica, i quali
sono
essenziali
per
la
regolazione
dell'espressione genica .
La risposta delle cellule bersaglio e' altamente specifica,
poiche' solo loro sono in grado di riconoscere il segnale in
quanto fornite di recettori specifici, presenti sulla
membrana plasmatica,
segnale.
per il prodotto delle cellule
Segnali extracellulari.
I segnali extracellulari
possono essere classificati in:
Endocrini - Paracrini
Autocrini.
Comunicazione Endocrina
Nella segnalazione endocrina le cellule
degli organi endocrini rilasciano
ormoni, cioe' sostanze segnale che
agiscono su gruppi di cellule bersaglio
lontane.
Comunicazione paracrina
Nella segnalazione di tipo paracrino, la cellula
bersaglio si trova in prossimita' della cellula
segnale e la sostanza che funge da segnale
agisce sul gruppo di cellule bersaglio adiacenti.
paracrina
Sono un esempio di segnalazione paracrina la
conduzione di un impulso elettrico da una
cellula nervosa a un'altra o da una cellula
nervosa ad una muscolare. Tale meccanismo
comporta la liberazione di sostanze segnale
dette neurotrasmettitori.
Comunicazione autocrina
La segnalazione autocrina e' caratterizzata
da
cellule
che
agiscono
contemporaneamente da segnale e da
bersaglio. Le cellule rispondono alle
sostanze che esse stesse hanno liberato.
Autocrina
Le cellule in coltura rispondono a fattori di crescita
che esse stesse producono. Molte cellule tumorali
presentano una iperproduzione di fattori di
crescita che stimolano l'accrescimento delle cellule
tumorali stesse e inducono le cellule normali
adiacenti, a proliferare trasformandole in cellule
tumorali.
Comunicazione endocrina
Il sangue e' la sede di rilascio di tali sostanze.
SEGNALI: gli ormoni
Gli ormoni possono essere
classificati in tre categorie:
1)piccole molecole lipofile
2) grandi molecole lipofile
3) molecole idrofile
piccole molecole lipofile
diffondono
attraverso
la
membrana
plasmatica
e
interagiscono
con
recettori
presenti nel citoplasma o nel
nucleo (steroidi, la tiroxina e
l'acido retinoico)
STEROIDI
CICLOPENTANOPERIDROFENANTRENE
ORMONI TIROIDEI
GRUPPO ACIDO
ACIDO RETINOICO
TRASPORTO EMATICO
• Questi ormoni (steroidi e tiroxina) siccome sono poco solubili
in acqua, nel sangue sono trasportati da molecole proteiche
carrier
• Gli ormoni steroidei, a differenza di quelli peptidici, sono
sintetizzati solamente da pochi organi e non vengono
immagazzinati
in
tessuti
di
riserva,
bensì
all'occorrenza e prontamente liberati nel plasma.
prodotti
• In questa sede, in virtù della loro scarsa solubilità in acqua,
devono necessariamente legarsi a proteine di trasporto:
• specifiche, come le SHBG (proteine di trasporto degli
ormoni
sessuali)
e
le
CBG
corticosteroidi)
• ed aspecifiche, come l'albumina.
(globuline
leganti
i
grandi molecole lipofile
si legano ai recettori della
membrana
plasmatica
(prostaglandine)
Le prostaglandìne sono
acidi
ciclopentanoici
derivati
dall'acido
arachidonico,
rivestono
che
un
biologico
ruolo
importante
come mediatori flogistici
(mediatori
dei
derivanti
infiammazioni).
processi
dalle
INIBIZIONE DELLE
PROSTAGLANDINE
L'inibizione della sintesi delle prostaglandine è
il meccanismo d'azione di una classe di farmaci
antiflogistici (antinfiammatori), antipiretici
(antifebbrili), e analgesici (antidolorifici) molto
diffusi: i FANS* a cui appartengono comuni
farmaci da banco come Ibuprofene, Nimesulide
(Aulin), Acido Acetilsalicilico (Aspirina) e
Ketoprofene (OKI)
*farmaci antiinfiammatori non steroidei
molecole idrofile
si legano a recettori sulla
superficie cellulare (insulina,
glucagone, epinefrina)
Ormoni idrofili
Molti ormoni non sono in grado di
diffondere attraverso la membrana poiché
non sono liposolubili.
Interagiscono quindi con recettori di
superficie.
Questa categoria di ormoni comprende
polipeptidi, come l’insulina e il
glucagone,
prodotti
del
pancreas
endocrino.
Meccanismi della Trasduzione
del segnale
Diversi meccanismi cellulari sono
coinvolti nella comunicazione del
segnale
I meccanismi innescati dall’attacco del segnale
(ligando) al recettore, sono:
1. formazione del secondo messaggero
2. apertura di una proteina canale e ingresso di
ioni
3. fosforilazione di una proteina
4. defosforilazione di una proteina
La funzione della trasduzione del
segnale è quella di poter regolare
il metabolismo cellulare e quindi
le funzioni dell’organismo.
Un esempio è il controllo della
glicemia
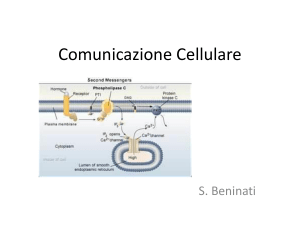
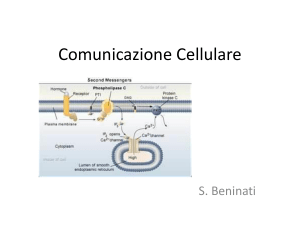
IL SECONDO MESSAGGERO
Per molti recettori, il legame con il ligando genera
un incremento di breve durata nella concentrazione
di un composto intracellulare che funge da secondo
messaggero.
Tipi di recettori della superficie
cellulare
Tipi diversi di recettori di superficie
inducono tipi diversi di meccanismo come
risposta cellulare.
RECETTORI DI SUPERFICIE
Sono stati definiti cinque tipi di recettori di
superficie:
1. canali ionici attivati dal calcio
2. protein chinasi attivata dal ligando
3. protein fosfatasi di residui tirosinici attivati
dal ligando
4. guanilato ciclasi attivata dal ligando
5. proteina G attivata dal ligando.
1. Canali ionici attivati dal calcio
La formazione del complesso ligando-recettore,
induce nel recettore della membrana plasmatica
una variazione conformazionale della proteina,
portando all’apertura di un canale ionico specifico.
Il flusso di ioni che ne deriva, altera la differenza
di potenziale elettrico tra i due lati della membrana
plasmatica. Il calcio ad esempio è un elemento
importante per la funzione di alcuni enzimi detti
calcio-dipendenti
Ligando
CALCIO
ENZIMA
Attivazione
2. protein chinasi attivata dal
ligando
La formazione del complesso ligando-recettore
induce l’attivazione dell’attività protein-chinasica
del recettore (capacità di fosforilazione).
In tali condizioni il recettore opera, nella sua
porzione citoplasmatica, la fosforilazione di una
proteina bersaglio, alterandone la struttura e quindi
l’attività.
Ligando
Porzione enzimatica
del recettore
ATTIVATA
INATTIVA
Il recettore proteico riceve il segnale. Il segnale provoca il mutamento
conformazionale della porzione intracellulare del recettore.
Tale porzione ha funzione enzimatica e il mutamento conformazionale provoca
l’esposizione del sito attivo.
Tale sito ora è pronto a ricevere il substrato da trasformare.
L’esposizione del sito attivo e la trasformazione del substrato, provocano
l’attivazione della proteina substrato. La porzione intracellulare del recettore
riassume la conformazione originale mentre il segnale lascia il sito di legame.
Il recettore riassume la forma inattiva. La proteina attivata a sua volta, a
cascata, attiva altre proteine
Legame del segnale al recettore
ORMONE
RECETTORE
fosforilazione
TRASDUZIONE DEL SEGNALE
Attivazione di proteine per mezzo della
fosforilazione
3. protein fosfatasi
Al contrario altri recettori sono invece
delle fosfatasi, la cui attività è indotta dal
ligando.
La
risposta
si
esplica
rimuovendo un gruppo fosforico da una
proteina bersaglio intracellulare.
3. protein fosfatasi di residui
tirosinici attivati dal ligando
Il ligando induce nella porzioni citoplasmatica del
recettore l’attività fosfatasica (scissione idrolitica
del legame fosforico), che rimuove un residuo
fosforico legato ad una tiroxina (amminoacido)
presente in una proteina bersaglio.
Il mutamento strutturale induce un cambiamento di
attività della proteina stessa.
Ligando
Porzione enzimatica
del recettore
4. guanilato ciclasi attivata dal
ligando
il legame del ligando al recettore permette
l’inizio della sintesi del secondo messaggero
GMP ciclico a partire da GTP.
Ligando
Porzione enzimatica
capace di scindere il
GTP
Secondo messaggero
5. proteina G attivata dal ligando.
Le proteine G sono proteine della membrana plasmatica
che agiscono da carrier per trasmettere i segnali generati
da un recettore al suo bersaglio cellulare (trasduttori
intracellulari).
Proteina G
Mentre nei casi precedenti il recettore
presenta anche la funzione di enzima nella
sua porzione intracellulare, nel caso della
proteina G sono presenti tre proteine sulla
membrana plasmatica:
1. Il recettore
2. La proteina G
3. L’enzima
1. Recettore
2. proteina G
3. enzima
PROTEINA G
Le proteine G sono polimeri trimerici con attività GTPasica*,
costituite quindi da tre subunità proteiche.
Le subunità α, β e γ sono disposte in una subunità α e un
complesso βγ.
*scinde il GTP
Proteina G
Queste proteine possono direttamente associarsi a
recettori di membrana, conosciuti come recettori a
serpentina o recettori collegati a proteine G.
Allo stato di riposo, la proteina G si trova libera nella forma di
trimero e con il GDP legato al sito specifico della subunità α
enzima inattivo
Nello stato inattivo la subunità α che lega GDP è strettamente legata al complesso βγ
(1) ma non legata al recettore.
Il legame di una molecola segnale* con il recettore provoca un cambiamento
conformazionale dello stesso, che porta all'associazione del trimero (α e βγ) con il
recettore. Questo si riflette direttamente sulla subunità α che a sua volta cambia
conformazione(2) e determina il rilascio del GDP legato e la sua sostituzione con il
GTP. Questa modificazione, a sua volta, causa la dissociazione del trimero dalla
proteina G con il rilascio di α-GTP e delle subunità βγ .
segnale
1
Forma inattiva
*ligando
2
L’unità α e la βγ libere, sono le forme attive della proteina G, che diffondono
nella membrana e possono legarsi con enzimi e canali ionici, causando, a
seconda dei casi, l'attivazione o l'inattivazione.
Il processo termina con l'idrolisi del GTP a GDP da parte della subunità α , che
possiede attività GTP-asica. La α-GDP che così si forma, si dissocia
dall'effettore* e si combina con βγ , completando in tal modo il ciclo.
Forma attiva
* Enzima o canale ionico
In conclusione entrambe le unità (α e βγ)
dissociate dal recettore, possono interagire con
proteine bersaglio, che sono generalmente
canali ionici oppure degli enzimi, quali
adenilciclasi,
fosfolipasi,
fosfodiesterasi.
Funzioni
Schematicamente le proteine G possono:
1).agire da tramite tra recettori a sette segmenti
transmembrana
ed
effettori
enzimatici
intracellulari (azioni a livello della membrana
plasmatica o delle membrane interne)
2).agire da tramite diretto (senza secondi
messaggeri) tra recettori e canali ionici
3).agire da tramite tra recettori ad attività
enzimatica e i loro effettori
Esempio di produzione di un secondo messaggero da parte dell’adrenalina attraverso
la proteina G.
adrenalina
Adenilato ciclasi
Fosforilazione delle
proteine operata da
una chinasi attivata
dal secondo
messaggero
Formazione
del secondo
messaggero