PIANTE
Organismi pluricellulari fotosintetici costituiti
da cellule eucariotiche vacuolate e con pareti
cellulosiche.
Eventi principali nell’evoluzione delle piante
Le piante terrestri si sono evolute
Oltre 500 milioni di anni fa da
alghe caroficee
GIMNOSPERME
“Piante a seme nudo”, circa 700 specie, sono le
piante a seme più primitive
Conifere
ANGIOSPERME
Piante a fiore. 250.000 specie
Monocotiledoni e Dicotiledoni
Tutte le piante nonostante la grande variabilità di morfologia attuano processi
fondamentalmente simili e sono basate sullo stesso schema architettonico
Monocotiledoni (grano, mais, riso, orchidee, bambù)
Dicotiledoni (fagioli, rose,, girasole, querce)
Elementi unificanti
Autotrofia (fotosintesi)
Immobilità
Parete cellulare (sostegno meccanico; impermeabilità)
Traspirazione
Strutture di trasporto per l’acqua e nutrienti (xilema, floema)
Crescita indeterminata (meristemi)
Il corpo vegetativo delle piante
consiste di due parti:
Il sistema radicale
Il sistema di parti aeree
Sistema di parti aeree:
fusto primario, rami
Sistema radicale:
radice primaria e
radici secondarie e terziarie
Ogni organo vegetale consiste di diversi tessuti e ogni
tessuto contiene molti tipi di cellule
Gli organi vegetali consistono di tre diversi tessuti
DERMICO
VASCOLARE
FONDAMENTALE
In complesso questi tessuti contengono circa 40 diversi tipi cellulari
Il corpo umano contiene diverse centinaia di tipi cellulari
Piante
organismi più semplici
Organizzazione dei tre sistemi di tessuti nel corpo vegetativo della
pianta
Tessuti dermici
Epidermide:
in piante giovani: singolo strato
di cellule con parete cellulare ispessita rivestita dalla
cuticola (differenziamento in tricomi o cellule
di guardia nelle foglie e in peli radicali
nella radice)
Periderma: in piante mature,
comprende la corteccia;
compare all’inizio dell’ispessimento
e dopo la caduta dell’epidermide
Tessuti fondamentali
Parenchima: cellule con parete sottile,
si trovano in tutti i tessuti.
Foglie: fotosintesi (mesofillo)
Fusto e radice: accumulo di amido e saccarosio
Semi: amiloplasti, corpi proteici e corpi oleosi
Floema: cellule compagne
Collenchima: pareti cellulari più spesse, allungate,
raggruppate in file verticali al di sotto dell’epidermide,
con funzione di supporto meccanico
Sclerenchima: cellule morte con pareti ispessite
e lignificate.
Formano fibre che sostengono e proteggono
il floema nei fusti
Tessuti vascolari
Xilema: elementi dei vasi (tracheidi),
cellule allungate, morte con pareti ispessite e
lignificate;Trasporto di acqua e soluti
dalle radici alle foglie
Floema: elementi dei tubi cribrosi
cellule cribrose), cellule vitali prive
di nucleo e tonoplasto.
Trasporto dei fotoassimilati
nelle regioni sink della pianta.
LO XILEMA ED IL FLOEMA
tessuto vascolare
xilema
floema
responsabile
del trasporto di
H2O e nutrienti
dalle radici alle
foglie
responsabile
del trasporto
di H2O e di
vari composti
nella pianta
XILEMA: trasporto dell’acqua e dei sali minerali
FLOEMA: trasporto dei fotosintati
Sviluppo e differenziamento
Piante: immobilità
Maggiore capacità di adattamenti fisiologici
Minore complessità anatomica rispetto agli animali
Crescita indeterminata: mediante attività dei
meristemi durante tutto il ciclo vitale
Negli animali sviluppo stabilito essenzialmente
durante l’embriogenesi
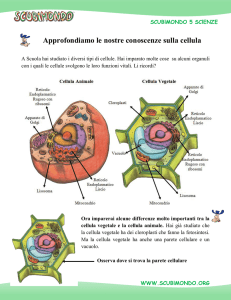
LA CELLULA VEGETALE
cellula eucariotica animale
cellula eucariotica vegetale
La cellula eucariota è caratterizzata dalla presenza di un
sistema esteso di endomembrane
Nella cellula vegetale:
Nucleo
Reticolo endoplasmatico
Golgi
Mitocondri
Cloroplasti
Vacuolo
Microcorpi (perossisomi, gliossisomi)
Oleosomi
Cloroplasti
Tilacoidi
PLASTIDI:
Cloroplasti: clorofilla; fotosintesi
Cromoplasti: carotenoidi; colorazione di frutti e
fiori
Leucoplasti:(amiloplasti); accumulo di amido
(nei tessuti di riserva del fusto, radice e seme)
Le cellule meristematiche contengono proplastidi, che mancano di
clorofilla e membrane interne. La luce innesca il differenziamento
in cloroplasti.
Semi germinanti al buio contengono ezioplasti che contengono i
corpi prolamellari
I cloroplasti (e i mitocondri ) sono organelli semiautonomi
Possiedono un proprio DNA e un sistema di sintesi proteica
(ribosomi, tRNA)
•Evoluti da batteri endosimbionti
•DNA in cromosomi circolari simili a quelli batterici
(localizzati in nucleoidi)
Genoma cloroplasto circa 145 kilobasi
Genoma mitocondrio 200 kilobasi
Il DNA del cloroplasto codifica per: rRNA, tRNA, Rubisco LS, e
molte altre proteine necessarie alla fotosintesi
GLICOSILGLICERIDI
captazione energia della luce
(reazioni alla luce)
produzione ATP e NADPH
utilizzazione NADPH e ATP per la riduzione di CO2 e la sintesi
di zuccheri
(reazioni al buio)
ORGANIZZAZIONE DELL’APPARATO FOTOSINTETICO
VACUOLO
•riserva
•digestione
•omeostasi ionica
•difesa da patogeni
Membrana del tonoplasto
Vacuoli
In cellule giovani provacuoli che si originano dal trans Golgi network
A maturità i provacuoli si fondono a formare grandi vacuoli
che occupano gran parte del volume cellulare
Raggiungimento di grandi dimensioni delle piante consentendo
economia biosintetica.
Contengono: acqua, ioni inorganici, zuccheri, enzimi, e
metaboliti secondari (difesa da organismi patogeni)
Ricchi di enzimi idrolitici: proteasi, ribonucleasi glicosidasi; ruolo litico
dei vacuoli, rilascio di tali enzimi nel citosol durante processi
degradativi come la senescenza
Vacuoli specializzati in accumulo di proteine –corpi proteicinei semi di alcune specie
L’accumulo di soluti garantisce al vacuolo la forza motrice
osmotica per l’assorbimento di acqua, necessaria alla distensione
cellulare vegetale
Pressione di turgore necessaria alla crescita generata per via osmotica
e responsabile del portamento ortotropo delle specie erbacee
che mancano di tessuti lignificati di sostegno
Microcorpi
Compartimenti circondati da una singola unità di membrana
specializzati in particolari vie metaboliche
Perossisomi:
presenti in tutti gli eucarioti nelle piante si trovano negli organi
Fotosintetizzanti
Rimuovono atomi di H da substrati consumando O2
RH2 + O2
R+ H2O2
H 2 O2
H2O + ½ O2
Il substrato è l’acido glicolico
(FOTORESPIRAZIONE)
Gliossisomi
presenti nei semi che accumulano grassi
contengono gli enzimi del ciclo del gliossilato che converte gli acidi
grassi di riserva in zuccheri, traslocati al germoglio per fornire
energia durante la germinazione.
RH2 + O2
H2O2
R + H2O2
H2O + ½ O2 (CATALASI)
FOTORESPIRAZIONE
GLIOSSISOMI
CORPI OLEOSI
Strutture di accumulo
dei trigliceridi
Apparato del Golgi
Rimozione mannosio; OH- glicosilazione (serina, treonina, tirosina)
Nei vegetali Golgi sito di sintesi delle emicellulose e pectine componenti della
parete cellulare
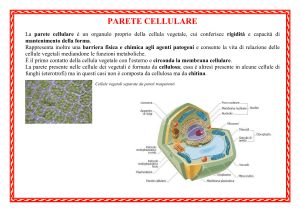
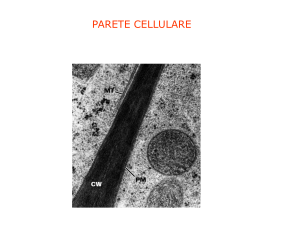
LA PARETE CELLULARE
FUNZIONI DELLA PARETE CELLULARE
•Conferimento della forza meccanica
•Mantenimento della forma
•Controllo dell’espansione
•Controllo del trasporto intercellulare
•Protezione da microorganismi patogeni
•Produzione di molecole segnale
forma cellulare
parenchima
fogliare
epidermide
petali
tracheide
tricoma
protoplasti
La parete cellulare non è una struttura uniforme
ma varia in composizione e aspetto nei vari tipi cellulari
LA PARETE CELLULARE
LAMELLA MEDIANA
Si forma nelle fasi finali della mitosi ed è
comune a cellule contigue
PARETE PRIMARIA
Si forma nelle cellule in crescita
Struttura simile in tutte le cellule
Spessore da 0.1 µm a 1 µm
PARETE SECONDARIA
Tipica delle cellule che hanno completato
il processo di sviluppo
È formata da più strati
Ha una composizione e struttura altamente variabile
(lignina)
parete
primaria
lamella
mediana
pareti
secondarie
Formazione della piastra cellulare (precursore parete cellulare)
fragmoplasto
PLASMODESMI
parete cellulare primaria
parete cellulare secondaria
Cellule che non hanno
completato l’accrescimento
Cellule che hanno
completato l’accrescimento
Componenti: Polisaccaridi (cellulosa, emicellulose, pectine) (85%)
proteine strutturali
Nella parete secondaria : Lignina
Componenti della parete cellulare
anomeria e
Principali zuccheri costituenti della parete cellulare
LA CELLULOSA
•polimero lineare di D(+)glucosio in legame (14)
•costituisce circa il 30% del peso delle pareti primarie
•struttura a microfibrille
•il grado di cristallizzazione e polimerizzazione è più elevato
nelle pareti secondarie
•l’orientamento delle microfibrille di nuova sintesi è
perpendicolare all’asse di crescita della cellula
lunghezza catene: da circa
2000 a circa 20000 residui
di glucosio
dimensioni microfibrille:
da circa 30 catene (alghe)
a circa 200 catene
diametro 5-15 nm)
LEGAMI IDROGENO
TRA LE CATENE DI
CELLULOSA
STRUTTURA DELL’AMIDO
complesso enzimatico della cellulosa sintasi
famiglia multigenica CesA
(1-4) -D-glucano (n) + UDP-glucosio
(1-4) -D-glucano (n+1) + UDP
Saccarosio sintasi: Saccarosio + UDP
UDP-Glucosio + Fruttosio
MATRICE
fase amorfa della parete
alto contenuto in H2O
polisaccaridi
emicellulose
pectine
proteine
HRGP (estensine)
GRP
PRP
AGP
EMICELLULOSE
gruppo eterogeneo di polisaccaridi
alta variabilità (tessuti, specie)
XILOGLUCANI
XILANI
GLUCANI
MANNANI
GLUCOMANNANI
non formano fibrille perché ramificati o non lineari
Sintetizzate nel Golgi
da glucosiltrasferasi
famiglia multigenicaCsI
XILOGLUCANI
9-3000 kDa
20% peso secco dicotiledoni
2% peso secco monocotiledoni
polimero di unità di D(+)glucosio in legame (14), con
residui di xilosio legati in posizione 6
allo xilosio è legata una molecola di galattosio in posizione 2
spesso al galattosio è legata una molecola di fucosio
La presenza di catene laterali negli xiloglucani impedisce
la formazione di fibrille come nella cellulosa
La lunghezza delle catene di xiloglucano è tale (50-500 nm)
che essi possono legare Insieme diverse fibrille di cellulosa
(distanza 20-40 nm)
XILANI
emicellulose più abbondanti nella
parete primaria delle
monocotiledoni
polimero lineare di D-xilosio in legame (14)
diverse catene laterali in posizione O-2 e O-3
(arabinosio e acido 4-ossi-metil glucuronico)
GLUCANI
tipici delle graminacee
polimeri di D(+) glucosio in legame (13) e (14)
GLUCOMANNANI
tipici delle conifere
D(+)glucosio e D(+)mannosio in rapporti variabili, in legame
(14)
galattosio in legame (16)
MANNANI
endosperma dei semi
mannosio in legame (14)
PECTINE
Gruppo eterogeneo di polisaccaridi contenenti uno
zucchero acido, l’acido galatturonico, e residui di ramnosio,
arabinosio, galattosio e xilosio
Il gruppo acido può
essere esterificato
formano gel in presenza di ioni bivalenti
(gelatine di frutta)
Omogalatturonano (acido poligalatturonico)
polimero lineare di unità di acido -D galatturonico unite da legami 1-4
(circa 200 unità, 100 nm)
Ramnogalatturano Tipo I
è la pectina più abbondante. Polimero composto da unità ripetute
del disaccaride 2)-D-Rha-(14) -D-GalA-(1
catene laterali legate al Rha (15 o 14)
Ramnogalatturonano Tipo II
struttura molto complessa (almeno 10 zuccheri differenti)
forma dimeri (legami con boro)
I gruppi carbossilici delle pectine possono essere esterificati
(CH3O-) enzimaticamente in genere nel Golgi durante la biosintesi
prevenendo così l’interazione con il calcio e riducendo la
gelificazione; una volta secreti parte dei gruppi esterici
possono essere rimossi dalle pectina metil esterasi
PROTEINE DELLA PARETE
ESTENSINA (HRGP) motivo Ser-(Hyp)4
residui di idrossiprolina glicosilati (arabinosio)
alcune serine glicosilate (galattosio)
forma legami intermolecolari insolubile
FUNZIONE STRUTTURALE
limitazione dell’estensione cellulare
resistenza a patogeni
ESTENSINA (HRGP)
Localizzate nel cambio, nel parenchima del floema e negli sclerenchimi
PROTEINE DELLA PARETE
PRP (proline-rich protein)
GRP (glycine-rich protein)
AGP (proteine arabino
galattaniche)
GRP : 70% di glicina; localizzate nei vasi xilematici
PRP: unità ripetitiva Pro-Pro-X-X-Lys; localizzate nei vasi xilematici, nelle fibre
e nel cortex
La quantità varia a seconda del tipo cellulare la maturazione o diversi stimoli
p es l’attacco di patogeni
AGP: altamente glicosilate, arabinogalattano (90%)
COME INTERAGISCONO I DIVERSI COMPONENTI DELLA PARETE?
legami tra pectine
modello trama-ordito
ESPANSIONE CELLULARE
Determinata dal TURGORE CELLLULARE
La direzionalità è determinata dalla struttura della parete
ORIENTAMENTO DELLE MICROFIBRILLE DI CELLULOSA
Orientamento isotropo
Orientamento anisotropo
(trasversale all’asse di crescita)
LA CRESCITA PER DISTENSIONE
AUMENTO DELLE DIMENSIONI SENZA DIVISIONE CELLULARE
CONSENTE ALLE PIANTE DI RAGGIUNGERE DIMENSIONI NOTEVOLI
CON RISPARMIO ENERGETICO E BIOSINTETICO
(espansione della superficie fogliare per la cattura dell’energia luminosa)
Il vacuolo accumula sostanze
aumenta la concentrazione osmotica
determinando una diminuzione del
potenziale idrico
vacuolo
vacuolo
aumento della P di turgore
(0,3-1 MPa)
Se la parete si rilassa, sotto
la spinta del turgore la cellula
aumenta di volume
CRESCITA PER DISTENSIONE
IL FATTORE DI RILASSAMENTO ELLA PARETE E’ IL PROTONE
INFATTI LA CRESCITA PER DISTENSIONE E’ CHIAMATA
ANCHE CRESCITA ACIDA
LA CRESCITA E’ ACCOMPAGNATA DA UN ABBASSAMENTO
DEL pH DELLA PARETE CELLULARE
crescita per distensione
Le espansine a valori di pH acidi sono in grado di interferire con la
formazione dei legami idrogeno tra i polisaccaridi di parete
H+
H+
H+
H
H+
H
+
+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H
+
H+
H+
Enzimi in grado di modificare la parete cellulare
Espansine: rompono i legami idrogeno (in arabidopsis
2 famiglie multigeniche EXPA; EXPB)
Egasi (endoglicosidasi) : tagliano (1-4)-d glucani
(xiloglucani e cellulosa) dall’ interno
XET (xiloglucanoendotraniglicosilasi): tagliano e
riformano legami tra catene di xiloglucani
Glicosidasi: rimuovono monosaccaridi dalle estremità
non riducenti di polisaccaridi o oligosaccaridi
XET: associate al meccanismo di crescita per distensione
PARETE SECONDARIA
cessazione crescita ispessimento della parete
primaria per stratificazione di materiale
•forma cellulare
•sostegno meccanico della pianta
•difesa
•riduzione della traspirazione
componenti:
cellulosa (in strati sovrapposti)
cuticola (cutina e cere)
suberina
lignina
Organizzazione della parete cellulare in cellule con parete secondaria (tracheidi)
LIGNINA
polimero di natura fenolica
costituenti:
alcol coniferilico
alcol sinapilico
alcol cumarilico
la via metabolica inizia dalla fenilalanina,
che si forma dalla via dell’acido scichimico
PAL= fenilalanina ammoniaca liasi
enzima regolatore del metabolismo secondario
sotto il controllo (enzima inducibile) di:
•luce
•ormoni
•ferite
•infezioni fungine
la polimerizzazione avviene mediante un meccanismo
ossidativo che comporta la formazione di radicali liberi
perossidasi: emoproteina H2O2-dipendente
laccasi: ossidasi O2-dipendente
non esiste una unità ripetitiva
Parete secondaria
Lignina
Xilani invece di xiloglucani
Maggiore percentuale di cellulosa
Microfibrille orientate parallelamente
Protezione
da microorganismi patogeni
Fonte di molecole segnale