Ricerca e sviluppo nei processi industriali
• Screening di biocatalizzatori con migliori proprietà
• Miglioramento dei ceppi industriali
• Ottimizzazione della composizione dei terreni di coltura
• Miglioramento delle procedure di recupero del prodotto
Massimizzazione delle
rese di produzione!
Miglioramento dei ceppi industriali
Il ceppo, risultato di una selezione per la capacità di produrre il prodotto, è
immediatamente oggetto di uno studio di fattibilità per il suo miglioramento
• versatilità utilizzo di substrati
• produzione sottoprodotti
• resistenza a stress durante il processo produttivo
• vitalità (ricircolo della biomassa)
• parametri fisiologici (µ, richiesta di O2, temperatura ottimale ecc.)
OBIETTIVO: ridurre ulteriormente i costi di produzione SENZA
aumentare il capitale investito
Tecniche di ingegneria cellulare e/o metabolica per esaltare alcune
proprietà del ceppo o per introdurre nuove funzioni
Miglioramento dei ceppi industriali
Gli strumenti impiegabili per il miglioramento sono legati a
quante e quali informazioni possediamo sul microrganismo e sul
processo.
informazioni
•
•
•
•
Tecniche di mutagenesi random
Ricombinazione (naturale o indotta) del materiale genetico
Metodi ricombinanti
Metodi –omici
Alterare i controlli di regolazione.
Eliminare i “colli di bottiglia” (reazioni lente).
Metodi di mutagenesi non ricombinanti
Metodi che
agiscono sul
DNA in
assenza di
replicazione
Analisi del fenotipo dei mutanti
Un buon schema di mutagenesi deve ottimizzare alcuni parametri al ceppo
Tasso di sopravvivenza, % di individui che sopravvivono alla mutagenesi (< 1%)
Frequenza di mutazione, n. di mutanti/n. di sopravviventi (massima possibile)
Numero di mutazioni introdotte
Aploide mononucleato
Per osservare il fenotipo
associato a mutazioni recessive
(quelle più spesso ricercate) è
necessario attendere il “ritardo
fenotipico”.
La giustificazione molecolare è
schematizzata a lato
Batteri in crescita
esponenziale
Mutazione a carico
di ribosomi
Mutazione a carico di
proteine di membrana
La mutagenesi random può non dare i risultati attesi perché:
• frequenza di mutazione non omogenea lungo tutto il genoma
• scarsa efficienza dei sistemi di riparo (tossicità elevata del mutageno)
• ulteriori mutazioni a soppressione
soppressori intragenici (nello stesso gene in cui è avvenuta la mutazione primaria)
soppressori extragenici (i più frequenti generano t-RNA mutati)
Metodi basati sulla ricombinazione per ottenere ceppi migliorati
Ricombinazione naturale
es. starter di Lactococcus per l’industria casearia resistenti alle infezioni fagiche, ottenuti
con l’impiego di plasmidi coniugativi
Ricombinazione sessuale
Fusione di protoplasti
E’ utile per quei ceppi che mancano di un ciclo sessuale o parasessuale ben definito!
Due tipologie cellulari diverse possono essere indotte a fondersi ed a rimescolare il loro
patrimonio genetico. Prima produzione dei protoplasti mediante idrolisi del rivestimento
cellulare, quindi fusione indotta da
PEG, polietilen glicole
Elettrofusione, mediante l’applicazione di campi elettrici pulsanti
Metodi ricombinanti
Azione mirata su geni specifici coinvolti nel processo produttivo:
•
•
•
•
•
•
•
Aumento del pool di precursori
Rimozione di vie metaboliche competitive o non necessarie
Inserimento, modifica o delezione di geni regolatori
Alterazione/modifica regioni di regolazione trascrizionale
Aumento del n. di copie di geni che codificano enzimi dei passaggi lenti
Aumento della efficienza di secrezione del prodotto
(per la produzione di antibiotici) Aumento della resistenza alla molecola
Metodi -omici
Corynebacterium glutamicum
Gram-positive, non-pathogenic, fast growing soil bacterium
Pathways leading to amino acids are much simpler
in their regulation when compared to E. coli:
-- very few enzymes are feedback controlled
-- good secretion
-- genome sequenced
Problems:
1) vector system are far from ideal
(E.coli vectors will not work)
2) Transformation rates are low
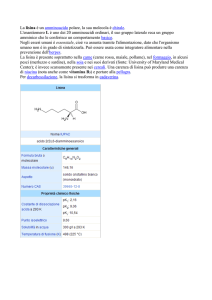
La lisina fa parte di
quel gruppo di
amminoacidi che
vengono sintetizzati
a partire da
ossalacetato; tali
amminoacidi sono
detti della famiglia
dell’aspartato
poiché condividono
l’intermedio
aspartato.
L-lisina
La prima tappa della via metabolica
prevede
la
fosforilazione
dell’aspartato ad aspartilfosfato ad
opera dell’aspartato chinasi
riduzione ad aspartato semialdeide
condensazione con una molecola di
piruvato e la conseguente sintesi
del diidropicolinato
riduzione a tetraidropicolinato
La via biosintetica della lisina in C.
glutamicum giunti al tetraidropicolinato
si biforca: una direzione prevede una
serie di quattro passaggi per giungere
alla
formazione
del
diamminopimelato
(via
della
succinilasi), nell’altra si ha il
passaggio
diretto
da
tetraidrodipicolinato
a
diamminopimelato mediato dalla
diamminopimelato deidrogenasi
Strain improvement
C. glutamicum, come tutti i batteri produttori di amminoacidi, normalmente non
produce grosse quantità di lisina quindi, per ottenere quantità di amminoacido
compatibili con le richiesta dell’industria, sono state sviluppate, nel tempo,
diverse procedure di miglioramento del ceppo microbico.
•il primo approccio utilizzato è stato quello di indurre delle mutazioni casuali (per
mutagenesi chimica o per irradiazione UV) nel ceppo selvatico di C. glutamicum e
di selezionare i mutanti iperproduttori di lisina.
•Una progettazione più razionale dei mutanti è iniziata negli anni ottanta
quando i primi studi sui flussi metabolici hanno consentito una conoscenza
più approfondita della via metabolica che conduce alla lisina
Con il termine Metabolic Flux Analysis (MFA) si intende il calcolo dei flussi intracellulari
utilizzando un modello stechiometrico che descriva
•le reazioni intracellulari più rilevanti
•i bilanci di materia
Dati il modello e le relazioni di bilancio, il calcolo dei flussi intracellulari si basa sulle misure
sperimentali dei flussi extracellulari, ovvero sulle velocità (misurabili sperimentalmente) di
consumo dei substrati e di secrezione dei prodotti Il risultato finale del calcolo dei flussi è
una mappa metabolica in cui ad ogni reazione biochimica è associata una stima della velocità
di reazione allo stato stazionario
Strain improvement
I primi studi sui flussi metabolici ( basati
sul bilancio di massa dei prodotti
extracellulari) oltre a caratterizzare i
principali nodi (punti di incontro di
diverse vie metaboliche) della via
metabolica dimostrarono che la resa di
lisina
dipendeva
dal
nodo
fosfoenolpiruvato/piruvato e dalla
resa di produzione dell’ossalacetato
da parte dei pathways anaplerotici.
Strain improvement
Il limite principale del metodo basato sul bilancio di massa dei prodotto
extracellulari, che non tiene conto degli intermedi intracellulari, è che non
consente di avere informazioni sulla reversibilità di certe reazioni poiché
tiene conto solo del flusso netto tra substrato e prodotto. Questo metodo,
inoltre, non consente di risolvere le singolarità strutturali di certe vie
metaboliche nel caso della lisina ad esempio non consentiva di discriminare
tra i due possibili meccanismi anaplerotici che conducono alla sintesi di
ossalacetato: la via della piruvato carbossilasi e quella della fosfoenolpiruvato
carbossilasi.
?
13C
Metabolic Flux Analysis (MFA)
13C
based flux balance model
•Use 13C-label to track the carbon flow
•Calculate the actual fluxes in metabolism.
A
v1
v2
C
D
H
CO2
B
EMP/PPP/ED
Kpathway
PGA
G
E
F
Glycerate
PEP
CO2
1
2
3
Phe
0.93 0.04
Lactate
0.88 0.10
Hydroxypyruvate
Pyruvate
0.96 0.02
CO2
0.81 0.19
Serine
CO2 or Formate
Acetate
Acetyl-CoA
0.81 0.19
0.39 0.43 0.17
Asp
0.06 0.35 0.58
Glycine
0.65 0.35
CIT
OAA
1
CO2
ICT
Glyoxylate
MAL
CO2
2
OXO
FUM
3
SUC
SUC
from
ICT
CO2
SUC
from
OXO
0.11 0.84
0.44 0.56
Formate
Glu
4
0.04 0.95
C1 pool
0.15 0.80
CO2
Strain improvement
Per superare limiti dei primi studi sui flussi metabolici è stato necessario acquisire
informazioni sui metaboliti intracellulari
Questo tipo di studi sui flussi metabolici hanno confermato l’importanza dei
pathways anaplerotici per la produzione di lisina e in particolare hanno stabilito
che il rifornimento dell’ossalacetato si deve sostanzialmente alla piruvato
carbossilasi (pyc), che rifornisce l’ossalacetato dalla carbossialzione del
piruvato.
Strain improvement
Con questo tipo di analisi dei flussi metabolici è stata
analizzata la distribuzione degli intermedi tra le due
vie per la biosintesi della lisina, quella della
diamminopmelato deidrogenasi e quella della succinilasi.
Tali studi hanno stabilito che via diretta rappresenta circa
il 40% del flusso verso la formazione della lisina e che la
scelta della via è funzione della fase di crescita. Infatti,
mentre all’inizio della crescita circa il 75% della lisina è
prodotto mediante la via della deidrogenasi, nella fase
finale del processo di fermentazione, essa è prodotta
quasi esclusivamente dalla via della succinilasi. Ciò è
giustificato dal fatto che la deidrogenasi ha una debole
affinità per uno dei suoi substrati ovvero l’ammonio, per
cui, quando esso è presente in basse concentrazioni la
deidrogenasi non può contribuire alla formazione della
lisina, in queste condizioni è favorito il flusso nella via
della succinilasi.
Controllo della concentrazione di
ammonio durante tutta la fermentazione
Strain improvement
Sono stati quindi progettati dei mutanti per
valutare l’effetto della sovraespressione
di pyc sulla crescita cellulare e sulla
produttività specifica di lisina in C.
glutamicum.
Sorprendentemente si è osservato che la
sovraespressione del gene codificante la
piruvato
carbossilasi
determina
un
sostanziale aumento di biomassa a cui non
corrisponde alcun effetto positivo sulla
produzione di lisina!!
Strain improvement
L’assenza dell’attesa iperproduzione di lisina ha suggerito
l’esistenza di altri potenziali “colli di bottiglia” nella via
metabolica che minimizzano l’effetto della sovraespressione
del gene pyc.
Già dai primi studi sui flussi metabolici era emerso che la
fosforilazione dell’aspartato rappresenta un punto
nevralgico della sintesi della lisina ma la sovraespressione
del gene ask aveva determinato solo una riduzione della
crescita accompagnata da un minimo aumento della
produttività specifica della lisina, questo risultato è dovuto
alla mancanza di equilibrio tra l’aumentata attività aspartato
chinasica, che devia il flusso metabolico verso la lisina, e
l’attività nativa della piruvato carbossilasi che contribuisce al
rifornimento di ossalacetato.
Strain improvement
Quindi la creazione di un ceppo capace di sovra produrre contemporaneamente
sia la piruvato carbossilasi che l’aspartato chinasi ha consentito di ottenere un
aumento del 250% della produttività specifica di lisina, senza influenzare la
velocità di crescita o la densità cellulare finale della coltura.
Strain improvement
Il sequenziamento del genoma di C. glutamicum ha aperto la strada a nuovi
approcci di ingegneria metabolica come ad esempio l’ingegneria metabolica
inversa definita dagli autori “genome-based strain recostruction”.
Strain improvement
Genome-based strain recostruction
Con questo approccio sono state identificate tre mutazioni puntiformi associate da
aumento di produttività del ceppo C. glutamicum:
la prima è stata individuata nel gene codificante l’aspartato chinasi
la seconda è a carico della piruvato carbossilasi
la mutazione introdotta nell’enzima omoserina deidrogenasi, che compete con la
diidropicolinato sintetasi per aspartato semialdeide, ne ha determinato una ridotta attività
catalitica.
Queste mutazioni introdotte per scambio allelico nel ceppo selvatico hanno avuto sulla
produzione del ceppo ingegnerizzato non un effetto addizionale ma sinergico dando vita
ad un batterio capace di produrre 80g/l di lisina in sole 27 ore.
Strain improvement…verso la system biology