Angela Iaccio
Il metabolismo delle lipoproteine ad alta densità e l’embrione umano
Introduzione
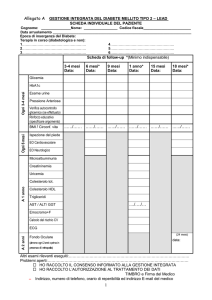
Le lipoproteine ad alta densità (HDL) si formano per l’aggregazione di vari componenti: colesterolo, trigliceridi, apolipoproteine e
fosfolipidi. In particolare, il fegato e l'intestino tenue secernono l'apolipoproteina apoA1, la quale riceve il colesterolo non
esterificato dei tessuti circostanti trasferitole da ABCA1 (ATP Binding Cassette A1). ApoA1 è
una molecola anfipatica che forma una cintura che si estende intorno al perimetro della
molecola sferica di HDL. Un deficit di apoA1 porta ad una riduzione dei livelli di colesterolo
nell’HDL, mentre un’apoA1 mutata riduce l’abilità di attivazione di lecitine indispensabili per
l’enzima colesterolo aciltransferasi (LCAT) e quindi comporta un abbassamento dei livelli di
HDL. Le HDL sono le molecole lipoproteiche dominanti nei fluidi follicolari (FF) umani durante
la follicolo genesi, in quanto serve al corpo luteo come substrato per la sintesi del
progesterone. Le proprietà anti-aterosclerogeniche e anti-infiammatorie dell’HDL sono state
attribuite in parte al trasportatore inverso del colesterolo, ovvero il recettore scavenger di
classe B tipo I (SR-BI).
Discussione
Il modello corrente della biogenesi di HDL è quello in cui il trasportatore ABCA1 trasferisce
colesterolo non esterificato alla molecola nascente detta HDL3 grazie all’interazione con apoA1. Successivamente il colesterolo non
esterificato viene esterificato dall’enzima LCAT che utilizza la lecitina come donatore dell’acido grasso che tramite l’azione di CETP
(apoD), scambia il colesterolo esterificato ricevendo in cambio trigliceridi mentre PLTP promuove lo scambio fosfolipidico tra HDL e
LDL ricchi di trigliceridi diventando HDL2. Queste lipoproteine neo sintetizzate raggiungono i tessuti steroidogenici (gonadi e
ghiandole surrenali) e il fegato a cui rilasciano il colesterolo esterificato tramite l’interazione con il recettore SR-BI, il cui omologo
umano è CLA-I. SR-BI è un recettore multi-ligando che lega apolipoproteine per mediare il trasferimento lipidico a livello cellulare.
Tali recettori sono situati in micro domini detti caveole utilizzate nell’endocitosi mediata da recettori del colesterolo come la
caveolina-1. Il meccanismo d’assorbimento lipidico-selettivo mediante SR-BI\CLAI è differente dal processo d’assorbimento che
effettuano i recettori dell’LDL. L’assorbimento del recettore lipidico di LDL attraverso l’endocitosi mediata da recettore avviene
tramite molecole di LDL intere mentre SR-BI interagisce con le lipoproteine per lo scambio lipidico selettivo. Il colesterolo, trasferito
al fegato, è trasportato, attraverso gli epatociti, nella bile mentre HDL povera in lipidi viene riciclata per un nuovo trasporto inverso
del colesterolo. La gonadotropina, agendo sul corpo luteo, media l’assorbimento selettivo del colesterolo attraverso un
meccanismo di AMP ciclico per la produzione di progesterone. Ulteriori prove sul ruolo fondamentale dell’HDL nella riproduzione
sono state fornite da topi femmine omozigote con una deficienza del trasportatore ABCAI in quanto hanno dimostrato una minore
fecondità, un HDL di dimensioni ridotte ed una modificazione significativa della composizione fosfolipidica. Un deficit familiare di
HDL è conosciuto anche come malattia di Tangier che causa un’aterosclerosi precoce per accumulo di colesterolo in vari organi e
una concentrazione plasmatica minore di HDL. La granulosa e le cellule della teca del follicolo ovarico acquisiscono grandi quantità
di colesterolo esterificato durante la peri-ovulazione che sotto stimolazione dell’ormone follicolo-stimolante (FSH) e dell’ormone
luteinizzante (LH) stimolano le cellule della granulosa a sintetizzare progesterone.
Conclusioni
Il deficit di ABCAI causa una carenza dell’efflusso di colesterolo che interferisce con l’omeostasi intra-follicolare del colesterolo tale
da danneggiare lo sviluppo normale dell’ovocita. La sfingosina1 associata con l’HDL nei FF induce proliferazione endoteliale e
angiogenesi con proprietà antiapoptotiche e antiossidanti, quindi le molecole di HDL provvedono allo sviluppo del follicolo. Queste
differenze suggeriscono che molecole HDL nei FF hanno una ridotta concentrazione di esteri del colesterolo nel core ma sono
arricchite sulla superficie da fosfolipidi. I processi che governano lo sviluppo della frammentazione embrionale in vitro possono
dipendere dal metabolismo del colesterolo, mentre i processi che regolano la divisione delle cellule embrionali possono essere
indipendenti dal metabolismo del colesterolo all’interno dello sviluppo del follicolo umano in quanto vi è un rimodellamento delle
particelle HDL all’interno del follicolo stesso. Si ipotizza che la relazione tra il colesterolo HDL e la frammentazione embrionale
rifletta l’alterazione dell’omeostasi del colesterolo intracellulare nell’oocita, in quanto influiscono sulla dinamica dei raft lipidici
coinvolti nel controllo dell’omeostasi del colesterolo e nella riorganizzazione dell’anello contrattile che facilita la citodieresi
embrionale.
Bibliografia
High-density lipoprotein metabolism and the human embryo. Fujimoto VY, Kane JP, Ishida BY, Bloom MS, Browne RW.
ABCA1 and ABCG1 synergize to mediate cholesterol export to apoA-I. Gelissen IC, Harris M, Rye KA, Quinn C, Brown AJ, Kockx M, Cartland
S, Packianathan M, Kritharides L, Jessup W.