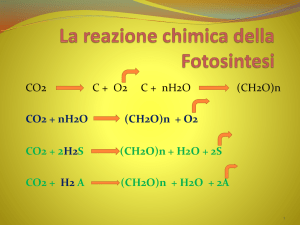
Fotoassimilazione della CO2
La nutrizione di un organismo autotrofo si basa sulla funzione
dei cloroplasti
La produzione di ATP e NADPH sono il presupposto per tutte le
altre attività del cloroplasto, che consistono nel produrre
intermedi C3 e idrati di carbonio, nel ridurre composti dell'azoto
ad ammine e dello zolfo a gruppi -SH
Il cloroplasto ha inoltre la proprietà di sintetizzare
autonomamente un grande numero di sostanze come i lipidi e
gli amminoacidi
In questo senso il cloroplasto assomiglia ad un organismo
procariotico fotosintetico come la Chlorella, un'alga unicellulare
Per
fotoassimilazione si intende una complessa sequenza
di reazioni attraverso le quali la CO2 viene trasformata in
zuccheri
Dapprima si passa attraverso un pool di molecole C3 dalle quali si
formano zuccheri di riserva e sostanze strutturali per nuove cellule
La sintesi degli zuccheri può svolgersi in tre direzioni:
1) sintesi di amido come riserva intermedia nel cloroplasto
2) trasporto degli intermedi nel citoplasma con lo scopo di
fornire i composti per la sintesi delle membrane
3) esporto degli intermedi nel citoplasma per la produzione di
sostanze di trasporto per trasportarle verso altre cellule
Per la fissazione della CO2 nel cloroplasto si susseguono tre
fasi: carbossilazione, riduzione e rigenerazione del substrato
Il processo di FOTOASSIMILAZIONE della CO2 è detto Ciclo di
Calvin dal nome di Melvin Calvin che nel 1961 vinse a questo
proposito il premio Nobel ed avviene nello stroma dei
cloroplasti
Il ciclo è caratterizzato da tre fasi distinte:
- FASE DI CARBOSSILAZIONE
-FASE DI RIDUZIONE
-FASE DI RIGENERAZIONE
FASE DELLA CARBOSSILAZIONE
Il primo step consiste nell’addizione di una molecola di CO2
ad un composto a cinque atomi di carbonio (RIBULOSIO1,5-BISFOSFATO, RuBP)
Questa reazione chiave è catalizzata dall’enzima Rubisco
(Ribulosio-1,5-bisfosfato carbossilasi/ossigenasi), uno degli
enzimi più diffusi sulla terra
L'enzima RUBISCO coordina i substrati RuBP e CO2 e
favorisce la formazione di un composto intermedio ipotetico
che non si libera, ma si scinde in 2 molecole di un
composto C3, l’ACIDO 3-FOSFO GLICERICO (PGA)
La RUBISCO è uno degli enzimi più importanti sulla Terra e
sicuramente uno dei più diffusi. E’ inoltre l’unico enzima che è
capace di fissare la Co2 atmosferica per la formazione di
biomassa
RAPPRESENTA IL PREREQUISITO ESSENZIALE PER
L’ESISTANZA DELLA VITA SULLA TERRA
Costituisce il 15% delle proteine totali del cloroplasto ed è stato
stimato che nel mondo se ne trovino circa 40 milioni di
tonnellate
Dal nome si comprende come questo enzima svolga anche
un’azione ossigenasica della quale tratteremo in seguito
Adesso concentreremo la nostra attenzione sulla funzione
carbossilasica che svolge nell’ambito del ciclo di Calvin
L’enzima Rubisco è un eteropolimero (circa 560
kDa) formato da otto subunità identiche (51-58
kDa) (unità LSU) codificate da un gene del
cloroplasto e da otto subunità identiche più
piccole (12-18 kDa) (unità SSU) codificate da
un gene nucleare
Su ogni subunità maggiore esiste un sito attivo
che lega i substrati CO2 e RuBP e l’attivatore
Mg2+, mentre l’azione delle unità SSu non è
ancora chiara in quanto non sembra essenziale
per la fissazione della CO2
Rubisco
Il processo di carbossilazione è veramente lento; infatti, al
livello di saturazione solo 3 moli di CO2 ed una di RuBP sono
convertite per secondo al livello del sito catalitico della Rubisco
A causa di questo lento ritmo di catalisi una grande quantità di
molecole di Rubisco sono necessarie per garantire il flusso
della fotosintesi
Si stima che la Rubisco può rappresentare anche il 50% delle
proteine totali solubili fogliari
Attivazione della Rubisco
La Rubisco è attiva solo quando un gruppo amminico della
lisina (posizione 201) reagisce con la CO2 per formare
CARBAMMATO a cui è legato il magnesio:
E-lys-NH3+ + CO2 ↔ E-lys-NH-COO- + 2H+ + Mg2+↔ E-lys-NHCOO-Mg2+
L’attivazione è determinata da cambiamenti conformazionali
della LSU ed è stabilizzata dallo ione magnesio
L’attivazione della Rubisco richiede ATP ed è catalizzata
dall’enzima Rubisco attivasi
L’attività della Rubisco è
dipendente dalla luce
Optimum di pH (circa 8)
È raggiunto nello stroma del cloroplasto a seguito
del trasporto elettronico durante il quale si ha un
trasporto netto di H+ dallo stroma al lumen dei
tilacoidi
Necessità della presenza di Mg2+
La concentrazione aumenta a seguito del
trasporto elettronico cloroplastico come
controione degli H+
• L’attivazione della Rubisco alla luce è mediata da una proteina
la Rubisco attivasi (RCA), la cui attività è a sua volta massima in
presenza di ATP e di RuBP e la sua funzione è quella di
catalizzare la carbammilazione della Rubisco da parte della
molecola attivatrice della CO2
•Un potente inibitore della Rubisco, il 2-carbossiarabinitolo-1fosfato (CA1P), raggiunge alte concentrazioni solo di notte
mentre è degradato alla luce da una fosfatasi che stacca il
fosfato dal carbonio 1
L'equilibrio
HCO3- + H +
H 2O + CO 2
è regolato, al livello delle membrane dei tilacoidi, dall’enzima
CARBONICO ANIDRASI
Gli ioni Mg++ sono essenziali per la carbossilazione oltre al
pH: valori elevati di pH favoriscono l’affinità per il Mg 2+ e la
formazione di un complesso carbammato-Mg 2+ che è il
presupposto per l’aggancio del ribulosio difosfato
Al buio la concentrazione di Mg2+ ed il pH sono subottimali
per la Rubisco
Alla luce si ha contemporaneamente un trasporto di H
dentro i tilacoidi ed un trasporto di Mg 2+ nello stroma
+
FASE DELLA CARBOSSILAZIONE
FASE DELLA RIDUZIONE
Il ciclo di Calvin o via riduttiva del pentosofosfato può svolgersi
solo alla luce in quanto dipende dalla disponibilità di ATP e
NADPH a forti dosi.
La riduzione del PGA avviene in due fasi catalizzate da una
chinasi (fosforilazione) (ENZIMA 1) e da una deidrogenasi NADPH
dipendente (ENZIMA 2) :
1) FOSFOGLICERATO-CHINASI (ATP-dipendente)
2) GLICERALDEIDE-3FOSFATO-DEIDROGENASI
(NADPH dipendente)
Il senso della reazione è determinato dalle concentrazioni di ATP
e NADPH (provenienti dal trasporto elettronico)
FASE DELLA RIGENERAZIONE
Una parte dei triosifosfati sono esportati dal cloroplasto per la
sintesi di molecole più complesse
In un processo noto come la fase di rigenerazione, una parte dei
triosi-fosfati vengono utilizzati per la sintesi del ribulosio-1,5bisfosfato necessario per la reazione iniziale del Calvin
Questo processo richiede energia che viene fornita dall’ATP.
Tredici sono gli enzimi coinvolti nell’intero ciclo di Calvin e la sua
efficienza di conversione dell’energia è di circa il 90%. Le reazioni
non coinvolgono la trasduzione dell’energia ma piuttosto
riarrangiamenti dell’energia chimica
Rigenerazione dei pentosi
1) Triosofosfato isomerasi
2) Aldolasi
3) Fruttosio-1,6-bisfosfato fosfatasi
4) Transchetolasi
5) Sedoeptulosio-1,7-bisfosfato fosfatasi
3
5
2
3
6
3
3
3
4
7
5
2
5
Attivazione degli enzimi della fase oscura
da parte della luce
Oltre la Rubisco anche altri enzimi del ciclo di Calvin sono regolati
dalla luce:
-3-FOSFO GLICERALDEIDE DEIDROGENASI
-FRUTTOSIO-1,6-BISFOSFATO FOSFATASI
-SEDOEPTULOSIO-1,7-BISFOSFATO FOSFATASI
-RIBULOSIO-1,5-BISFOSFATO CHINASI
I meccanismi di attivazione di questi enzimi sono indiretti in quanto
l’energia luminosa non è direttamente assorbita dagli enzimi che sono
incolori
E’ coinvolta l’energia luminosa assorbita dal PSII e dal PSI del
trasporto elettronico
Questi enzimi contengono nella loro molecola ponti disolfuro (SS) che alla luce sono ridotti a due gruppi sulfidrici (SH)
La riduzione del ponte disolfuro determina una importante
modificazione nella struttura proteica per cui la sua attività
aumenta
Nella riduzione sono utilizzati gli elettroni che provengono dalla
fotolisi dell’acqua nel PSII e che non vengono utilizzati per la
riduzione del NADP+
In particolare il processo vede coinvolte le tioredoxine attraverso
l’azione dell’enzima FERREDOSSINA-TIOREDOXINA RIDUTTASI
Nei cloroplasti intensamente illuminati nei quali le riserve del
NADP+ sono ridotte, si accumula Fd(Fe2+) che determina
l’attivazione degli enzimi del ciclo di Calvin
Bilancio energetico della fotosintesi
L’ATP ed il NADPH necessari per il ciclo di Calvin vengono
rilasciati nello stroma durante la fase luminosa del processo
Occorrono 2 fotoni perché un elettrone passi attraverso il PSII
ed il PSI; 2 sono gli elettroni richiesti però per ridurre una
molecola di NADP+ quindi saranno necessari 4 fotoni per ridurre
una molecola di NADPH
Ciò corrisponde anche ad 8 fotoni per molecola di O2 evoluta
Ridurre una mole di CO2 a zucchero (CH2O)n richiede 2 moli di
NADPH e 3 di ATP. Se consideriamo una molecola di glucosio
prodotta, 6 molecole di CO2 devono essere fissate ed assimilate
e questo richiederà 12 molecole di ATP e 12 di NADPH
12 NADP+ + 18 ADP + 18 Pi + 6H+ 48 fotoni 6O2 + 12
NADPH + 18 ATP + 6H2O
6H2O + 6CO2 + 48 fotoni C6H12O6+ 6O2
Fotorespirazione
La Rubisco come abbiamo visto può comportarsi come
un’ossigenasi anziché come una carbossilasi
Le cellule vegetali contengono i mitocondri e quindi respirano
per produrre ATP. In questo processo viene consumato
ossigeno e prodotta CO2. Il processo, noto con il nome di
respirazione, avviene nelle piante indipendentemente dalla luce
Tuttavia, in molte specie vegetali un altro tipo di respirazione
avviene solo in presenza di luce ed è noto con il nome di
FOTORESPIRAZIONE.
Questo
processo
comporta
la
cooperazione di cloroplasti, perossisomi e mitocondri
La reazione di ossigenasi operata dalla Rubisco determina la
formazione dell’acido 3-fosfoglicerico e di un prodotto a due
atomi di carbonio, l’acido fosfoglicolico
Il perché della fotorespirazione
La fotorespirazione potrebbe essere considerato un processo
inutile
Infatti:
• il ribulosio-5-P viene sottratto dal ciclo di Calvin
• viene consumato O2 e liberata CO2
• solo una parte degli atomi di C ritorna al cloroplasto
• si consuma ATP
Alcuni Autori hanno ipotizzato che la fotorespirazione che
diminuisce l’efficienza della fotosintesi, costituisca un
mezzo per eliminare NADPH ed ATP prodotti in eccesso
quando i livelli di irradianza sono eccessivamente elevati
L’eliminazione di potere riducente potrebbe preservare dal
danneggiamento i pigmenti presenti nel cloroplasto
Fissazione della CO2 nella piante C4
(via degli acidi bicarbossilici)
In alcune specie la reazione primaria di carbossilazione è
diversa da quella a cui partecipa il ribulosio 1,5-difosfato ed il
primo prodotto della reazione non è il 3PGA, bensì una acido a 4
atomi di carbonio
Le specie che hanno questo tipo di metabolismo vengono
comunemente dette specie C4, mentre quelle che fissano
inizialmente la CO2 nel 3PGA sono dette specie C3
La maggior parte delle specie C4 sono monocotiledoni, in
particolare graminacee, anche se appartengono al gruppo più
300 dicotiledoni
Tra le piante coltivate si annoverano fra le C4 il mais, il sorgo e
la canna da zucchero
Tutte le piante arboree ed arbustive sono piante C3
Nelle specie C4 esiste anche una divisione di compiti tra due
diversi tipi di cellule fotosintetiche: le cellule del mesofillo e
quelle della guaina del fascio (ANATOMIA KRANZ)
Il processo delle piante C4 è considerato più evoluto del
sistema C3 ed è fortemente connesso alle condizioni
ambientali
Le alte temperature favoriscono l’attività ossigenasica rispetto
a quella carbossilativa della Rubisco e questa ha come
conseguenza una minore efficienza fotosintetica. Questo
sembra dovuto in parte a cambiamenti nella solubilità di O2 e
della CO2
La via comprende la fissazione dello ione HCO3- nelle cellule
esterne del mesofillo, la fissazione di questo in un acido a 4 atomi
di carbonio, il trasferimento di questo nelle cellule della guaina del
fascio ed quindi il rilascio della CO2 che viene ri-fissata mediante
il ciclo di Calvin-Benson
Nelle piante C4 infatti gli enzimi del ciclo di Calvin sono localizzati
esclusivamente nelle cellule della guaina del fascio, mentre
l’acido organico (ossalacetato) prodotto dalla fissazione dello
ione HCO3- è presente sono nel citosol delle cellule del mesofillo
L’enzima chiave per la fissazione è rappresentato
dalla
fosfoenolpiruvato
carbossilasi
(PEP
carbossilasi), la quale utilizza lo ione HCO3- e non
la CO2
Questo determina una maggiore efficienza in
quanto in soluzione acquosa è favorita la
formazione dello ione HCO3- rispetto alla CO2
gassosa
La PEP carbossilasi inoltre non può fissare
l’ossigeno che ha un struttura tridimensionale
simile alla CO2 ma non all’HCO3La reazione iniziale catalizzata dalla PEPcase è la
combinazione con il substrato di partenza, il
fosfoenolpiruvato (PEP)
con
lo
ione
carbonato per formare
ossalacetato. Questa
reazione è irreversibile
Sono conosciute tre varianti della fotosintesi nelle
piante C4, che differiscono per:
- acido C4 trasportato nel cellule della guaina
- meccanismo di decarbossilazione che avviene
nelle cellule della guaina
FOTOSINTESI C4 TIPO ENZIMA
MALICO NADP+-DIPENDENTE
FOTOSINTESI C4 TIPO ENZIMA
MALICO NAD+-DIPENDENTE
FOTOSINTESI C4 TIPO
CARBOSSICHINASI
PEP
UNA DELLE REAZIONI CHIAVE DEL METABOLISMO DELLE
PIANTE C4 E’ LA SINTESI DEL PEP CATALIZZATA DALL’ENZIMA
PIRUVATO-ORTOFOSFATO DICHINASI
PIRUVATO + ATP + Pi PEP + AMP + PPi
LA REAZIONE RICHIEDE QUINDI LA CONVERSIONE DI 2 MOLI DI
ATP PER OGNI MOLECOLA DI CO2 RILASCIATA NELLE CELLULE
DELLA GUAINA DEL FASCIO
SI HA QUINDI UN CONSUMO SUPERIORE DI ENERGIA RISPETTO
AL METABOLISMO C3
OCCORRE TUTTAVIA SOTTOLINEARE L’ASSENZA IN QUESTE
PIANTE DEL PROCESSO DI FOTORESPIRAZIONE CHE
TIPICAMENTE CONSUMA ATP
E’ INFATTI ACCERTATO CHE I CLOROPLASTI
DELLE CELLULE DELLA GUAINA DEL FASCIO
NON
HANNO
MEMBRANE
IMPILATE
E
MOSTRANO UNA BASSISSIMA ATTIVITA’ DEL PSII
QUESTO ESCLUDE IL PROCESSO FI FOTOLISI
DELL’ACQUA CON LIBERAZIONE DI OSSIGENO
FAVORENDO
QUINDI
L’ATTIVITA’
CARBOSSILATIVA DELLA RUBISCO
ANCHE L’ATTIVITA’ DI ALCUNI ENZIMI DEL CICLO
C4 SONO REGOLATI DALLA LUCE ANCHE SE
CON SISTEMI DIVERSI:
- Malato Deidrogenasi NADP-dipendente
- PEPcase
- Piruvato- Ortofosfato Dichinasi
MALATO DEIDROGENASI NADPDIPENDENTE
La sua regolazione avviene attraverso il ciclo delle
tioredoxine
PEPcase
Sono entrambe regolate per fosforilazione
La PEPcase NON FOSFORILATA presente nelle
foglie adattate al buio è inibita dal malato ed ha
affinità bassa per il PEP: è quindi una forma
essenzialmente inattiva al buio
Alla luce vie attivata una CHINASI che fosforila un
residuo serinico della PEPcase ed in questa forma
l’enzima è meno sensibile alla concentrazione di
malato
Meno attivo
PEP carbossilasi
chinasi
Luce
Buio
Più attivo
PEP carbossilasi
ATP
chinasi
ADP
OH
PEPcase
OP
Ser
PEPcase
Ser
PIRUVATO- ORTOFOSFATO DICHINASI
(PPDK)
La regolazione di questo enzima è piuttosto
complessa e coinvolge diverse reazioni che danno
un enzima fosforilato inattivo al buio ed un enzima
defosforilato alla luce
La sorgente del gruppo fosfato è l’ADP la cui
concentrazione aumenta al buio quando la
concentrazione di ATP è bassa ed aumenta quindi
la fosforilazione della PIRUVATO-ORTOFOSFATO
DICHINASI
La proteina di regolazione è bicatalitica e regola
quindi sia la fosforilazione che la defosforilazione
NON E’ ANCORA CHIARO IL RUOLO DELLA LUCE
OH
ADP
AMP
Thr
OP
Thr
PPDK
PROTEINA
REGOLATRICE
Attivo
LUCE
PPDK
Inattivo
PPi
Pi
BUIO
Metabolismo acido delle crassulacee
(PIANTE CAM)
E’ una ulteriore fissazione della CO2 ed è tipica di specie che
vivono in climi aridi. Esse sono caratterizzate da habitus
xerofitico con rapporti superficie/volume relativamente bassi, una
spessa cuticola e conseguente basso ritmo di traspirazione
La maggior parte delle cellule fotosintetizzanti delle foglie o del
fusto forma un tessuto lacunoso e presenta vacuoli molto grossi.
Anche se poco distinguibili sono presenti anche le cellule della
guaina del fascio
Il metabolismo attraverso il quale queste piante fissano la CO2 è
peculiare ed è stato definito come metabolismo acido delle
crassulacee (CAM)
Si ha una suddivisione non solo spaziale ma anche
temporale del processo di assimilazione e fissazione della
CO2
Per questo motivo esse aprono gli stromi durante la notte (la
temperatura è più bassa e l’umidità relativa più elevata) e
fissano la CO2 nell’acido malico
L’acido malico viene “raccolto” negli enormi vacuoli presenti in
queste piante. Nel metabolismo CAM è ancora poco chiaro
perché queste piante fissino la CO2 attraverso la
PEPcarbossilasi, dato che entrambi gli enzimi sono presenti.
Una spiegazione possibile è data dal fatto che la PEP
carbossilasi durante il giorno si converte in una forma inattiva,
che ha affinità minore per il PEP ed è fortemente inibita
dall’acido malico; ma si verificano anche variazioni di altri
enzimi che favoriscono la fissazione della CO2 da parte della
PEP carbossilasi durante la notte e quella della Rubisco durante
il giorno
IL MECCANISMO CHE TIPICAMENTE INIBISCE LA PEPcase
NELLE PIANTE C4 AL BUIO NON AGISCE OVVIAMENTE
NELLE PIANTE CAM
L’ATTIVAZIONE DELLA PEPcase ALLA NOTTE E LA SUA
DISATTIVAZIONE
FURANTE
IL
GIORNO
AVVIENE
ATTRAVERSO UN RITMO CIRCADIANO ENDOGENO
LA FORMA DIURNA DELL’ENZIMA E’ INIBITA DAL MALATO,
MENTRE LA FORMA NOTTURNA E’ INSENSIBILE AL MALATO
Differenze tra piante C3
e piante C4
I meccanismi di fissazione della CO2 delle C4 e delle CAM sono
considerati una evoluzione adattativa del metabolismo C3; nelle
prime a condizioni di alta irradianza, elevate temperature e
carenza idrica, nelle seconde ad ambienti decisamente desertici
Piante C4:
• sviluppo di un meccanismo di fissazione più efficiente, con
velocità di assimilazione della CO2 mediamente più elevate delle
piante C3
• una fissazione più efficiente anche a basse concentrazioni di
CO2, perché la PEP carbossilasi ha localizzazione
citoplasmatica (minori resistenze al flusso di CO2), utilizza
come substrato l’HCO3-, non subisce la competizione dell’O2
rispetto alla CO2 ed ha una affinità per il substrato maggiore
della Rubisco. Infatti il punto di compensazione per la CO2 (cioè
la concentrazione di CO2 esterna per la quale il flusso
fotosintetico eguaglia quello respiratorio) è nella C4 più basso
(intorno a 10 ppm) rispetto alle C3 (50-60 ppm)
• non presentano fotorespirazione apparente sia perché
nei cloroplasti delle cellule più interne (guaina del
fascio), dove interviene la Rubisco, la competizione
dell’O2 è minore, sia perché non è presente il PSII e non
si ha quindi splitting dell’acqua e liberazione di O2
• la possibilità di fotorespirazione nelle piante C4 non è
da escludersi completamente; sono presenti infatti tutti
gli enzimi del ciclo fotorespiratorio. Tuttavia l’eventuale
CO2 liberata per fotorespirazione può essere rifissata
dalla PEP carbossilasi al livello delle cellule del
mesofillo più esterne. Da notare che l’assenza del
trasporto elettronico non ciclico nei cloroplasti delle
cellule più interne rappresenterebbe un problema se gli
equivalenti riduttivi in forma di NADPH non venissero
ricostituiti nella reazione di decarbossilazione
ossidativa dell’acido malico catalizzata dall’enzima
malico
In definitiva il sistema di fissazione delle C4
realizza fra le cellule del mesofillo e quelle della
guaina una pompa per la concentrazione della CO2
ed uno shuttle per il trasferimento degli
equivalenti riduttivi (NADPH) entrambi mediati
dall’acido malico