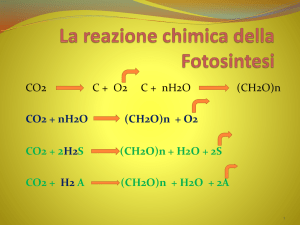
ogni anno 200 miliardi di tonnellate di CO2
vengono convertite in biomassa
La maggior parte di questo carbonio viene
incorporato in composti organici attraverso le
reazioni di riduzione del carbonio associate alla
fotosintesi
Tutti gli eucarioti fotosintetici,
dall’alga più primitiva
all’angiosperma più evoluta,
riducono la CO2 in carboidrati.
CICLO DI CALVIN
ciclo riduttivo dei pentosi fosfati, RPP
riduzione fotosintetica del carbonio, PCR
Approccio cinetico
14CO
2
Chlorella
analisi intermedi
marcati
dopo 5 s
dopo 30 s
Qual è l’accettore della CO2?
La susseguente scoperta che i pentosi
monofosfati e un pentoso difosfato (il
ribulosio) partecipassero al ciclo suggerì la
possibilità che l’accettore iniziale della CO2
fosse un composto a cinque di carbonio
che, dopo aver reagito con la CO2,
generava due molecole di 3-fosfoglicerico.
Questo sconvolgimento concettuale portò
rapidamente all’identificazione del
ribulosio 1,5-difosfato come accettore
della CO2.
Il ciclo di Calvin procede attraverso
tredici tappe divise in tre fasi
il ciclo di Calvin è autocatalitico
rigenera più substrato di quanto ne consumi
5 RuBP + 5 CO2 + 9 H2O + 16 ATP + 10 NADPH
→ 6 RuBP + 14 Pi + 6 H+ + 16 ADP +10 NADP+
stechiometria generale
6 CO2 + 11 H2O + 18 ATP + 12 NADPH →
fruttosio 6-P +12 NADP+ + 6 H+ + 18 ADP +17 Pi
il ciclo di Calvin consuma 3 ATP e 2 NADPH per
ogni molecola di CO2 fissata
Rubisco
ribulosio 1,5 bisfosfato carbossilasi/
ossigenasi
uno dei tre enzimi esclusivi del ciclo di Calvin
La Rubisco è la proteina solubile
più abbondante del cloroplasto e
probabilmente una delle più
abbondanti della biosfera
enzima multimerico (560 kDa)
8 subunità grandi (55 kDa)
8 subunità piccole (14 kDa)
(L8S8)
Le subunità L sono codificate dal
genoma cloroplastico, le subunità
S dal genoma nucleare.
luce
genoma nucleo → rbcS (precursore subunità piccola)
genoma cloroplasto → rbcL (subunità grande)
La subunità grande è
codificata dal DNA del
c l o r o p l a s t o, m e n t r e l a
subunità piccola e trascritta
dal nucleo, tradotta nel
citoplasma e trasportata nel
cloroplasto. L’assemblaggio
di Rubisco richiede la
partecipazione attiva di un
complesso traduzionale
come la chaperonina
cloroplastica altro prodotto
a localizzazione plastidica di
un gene nucleare.
40% della frazione proteica
totale delle foglie
Affinità dell’ enzima per la
CO2 è sufficientemente
elevata
nello stroma [rubisco] = 4
mM 500 volte > [CO2]
Carbossilazione
3-PGA
Acido 3-fosfoglicerico
ΔG = -51 KJ
Favorisce il progredire della reazione
Primo
intermedio
stabile della
fotosintesi
Processo di
riduzione
L’enzima gliceraldeide-3-fosfato deidrogenasi è simile
a quello della glicolisi, ad eccezione dell’utilizzo come
coenzima del NADP, al posto del NAD. Durante lo
sviluppo dei cloroplasti (inverdimento) viene
sintetizzata una forma dell’enzima legata al NADP e
nelle reazioni biosintetiche è preferibilmente utilizzata
la forma di enzima che richiede NADP.
rigenerazione del RuBP
La continua fissazione della CO2
ha bisogno che l’accettore della
CO2, il ribulosio 1,5-difosfato,
venga continuamente rigenerato.
Per evitare la mancanza di
intermedi nel ciclo vengono
formate 3 molecole di ribulosio,
recuperando gli atomi di
carbonio dalle 5 molecole di
triosi fosfati.
una molecola di gliceraldeide 3-fosfato è convertita a
diidrossiacetone fosfato da una trioso fosfato isomerasi
il diidrossiacetone fosfato va incontro a
condensazione aldolica con una molecola di
gliceraldeide 3-fosfato
il fruttosio 1,6-bisfosfato viene defosforilato a fruttosio 6-fosfato
il fruttosio 6-fosfato reagisce con la gliceraldeide 3-fosfato per
formare eritrosio 4- fosfato e xilosio 5-fosfato
L’eritrosio 4-fosfato si combina con una molecola di trioso fosfato
(sotto forma di deidrossiacetone 3-fosfato) per formare uno
zucchero a 7 atomi di C sedoeptulosio 1,7-difosfato
idrolizzato
Il septulosio 6 fosfato
dona una unità a due
atomi di C a una
m o l e c o l a d i
gliceraldeide 3-fosfato
producendo ribosio 5fosfato e xilulosio 5fosfato
regolazione del ciclo di Calvin
efficienza termodinamica indica la presenza di qualche
forma di regolazione
90% (vitro)
80% (vivo)
in modo da assicurare
tutti gli intermedi sono presenti in condizioni adeguate
il ciclo è spento quando le condizioni non sono favorevoli
REGOLAZIONE QUANTITÀ ENZIMI
controllo espressione genica
REGOLAZIONE ATTIVITA’ ENZIMI
la luce regola l’attività di numerosi enzimi del ciclo
di Calvin
Alcuni enzimi sono in comune con la via glicolitica e
quella dei pentosi fosfati è essenziale che, alla luce
l’apparato sintetico sia “acceso” e che l’apparato
degradativo sia “spento”
regolazione Rubisco
una molecola di CO2 reagisce con la Lys nel
sito attivo
reazione favorita dall’aumento di pH nello
stroma associata alla illuminazione del
cloroplasto
Complesso
carbammato
Mg2+
pH 7 → 8
[Mg2+] 1-3 mM → 3-6 mM
La luce determina cambiamenti reversibili degli ioni presenti nello
stroma. A seguito di illuminazione,i protoni sono pompati dallo stroma
nel lume dei tilacoidi. Questo efflusso di protoni, accoppiato
all’assorbimento di Mg2+ ,porta all’aumento del pH dello stroma da circa
7a8
Attivazione della rubisco
in vitro [CO2] = mM
in vivo [CO2] = µM
il RuBP lega la Rubisco → no carbammilazione
Il ruolo della rubisco attivasi è di riattivare la
rubisco che è stata disattivata da zuccheri
difosfati. La rubisco attivasi accelera il rilascio di
zuccheri fosfati legati che inibiscono la forma
carbammilata della rubisco.
Alcuni enzimi del ciclo di Calvin sono attivi solo alla luce
gliceraldeide 3-fosfato deidrogenasi
fruttosio 1,6-bisfosfatasi
sedoeptuloso 1,7-bisfasfatasi
Rubisco attivasi
inattivo
attivo
SISTEMA
FERREDOSSINA -TIOREDOSSINA
Costituisce un meccanismo
per l’attivazione o
disattivazione degli enzimi in
dipendenza della luce. Il
trasferimento fotosintetico di
elettoni da parte del PSI
porta alla riduzione della
ferrodossina proteina Fe-S.
La ferredossina ridotta, a sua
volta, riduce la tioredossina,
una proteina di regolazione a
disolfuri. La tioredossina
ridotta può ridurre i legami
disolfuro di numerose
proteine bersaglio,
modulando la loro attività
Gli enzimi del ciclo di Calvin
sottoposti a regolazione dalla
tioredossina ridotta vengono
attivati per riduzione alla luce e
disattivati per ossidazione al
buio
FOTORESPIRAZIONE
il ciclo PCO (photorespiratory carbon
oxidation) recupera parte del carbonio
perduto nella reazione di ossigenazione
del ribulosio 1,5-bisfosfato
Il funzionamento del ciclo per
l’ossidazione fotorespiratoria del
carbonio implica l’interazione
cooperativa fra tre organuli: i
cloroplasti, i mitocondri e i
perossisomi.
La traslocazione dei metaboliti fra
questi organuli è dedotta dalla
localizzazione di numerosi enzimi
il glicolato esce dai cloroplasti
tramite un trasportatore specifico
entra nei perossisomi (diffusione?)
acido glicolico
acido gliossilico +H2O2
contiene FMN
Acido gliossilico viene transaminato, il donatore del gruppo
amminico è probabilmente l’acido glutammico
e il prodotto della reazione è aa glicina
la glicina passa dal perossisoma al mitocondrio
2 glicina
serina,
CO2
la serina passa
nel perossisoma
Perossisoma
la serina è deaminata a idrossipiruvato
(l’accettore dell’NH2 è l’acido α-chetoglutarico)
l’idrossipiruvato è ridotto a glicerato
il glicerato passa nel cloroplasto dove viene
fosforilato per dare acido 3-fosfoglicerico
glicerato
3-fosfoglicerato
CO2
il ciclo PCO recupera il 75% del C
Il ciclo di Calvin è in
grado di funzionare
i n
m a n i e r a
autonoma, mentre il
PCO (ossidazione
fotorespiratoria del
carbonio) necessita
di un continuo
funzionamento del
ciclo di Calvin per
poter usufruire del
m a t e r i a l e d i
p a r t e n z a , i l
ribulosio-1,5difosfato
a 25°C
carbossilazione
ossigenazione
=
3
1
cosa succede quando la temperatura aumenta?
legge di Henry
[gas] µM = Pgas x
αx
106
V0
α = coefficiente Bunsen di assorbimento
(volume gas assorbito da un volume di acqua
alla pressione di 1 atmosfera)
V0 =volume normale di un gas ideale a T e P standard
(V0 =22.4 L mol-1)
La solubilità di un gas diminuisce man mano che la
temperatura aumenta
Le concentrazioni dei due gas CO2 e O2 variano con la
temperatura in quanto la solubilità di un gas diminuisce
con l’aumentare della temperatura.
Il rapporto fra carbossilazione e ossigenazione catalizzate
dalla rubisco diminuisce e aumenta il rapporto fra
fotorespirazione e la fotosintesi