Le reazioni della fotosintesi
1. Reazioni della fase luminosa:
Producono NADPH e ATP
2. Reazioni di fissazione del carbonio
Producono triosi fosfati
Fissazione e riduzione della CO2
L’ATP e il NADPH sono utilizzate per la riduzione della
CO2 in carboidrato
Questa
reazione si
realizza
nello stroma
del
cloroplasto
Ciclo di Calvin o
Ciclo riduttivo dei pentosi
fosfati
Tre fasi
1) Carbossilazione
(PGA)
RUBISCO
Km(CO2)= 12 µM, Km(RUBP)=60 µM, Km(O2)=250 µM
2) Riduzione
Il 3-PGA e’ ridotto ad un carboidrato a 3 atomi
di carbonio nella forma di gliceraldeide 3-P
(GAP)
3) Rigenerazione
A partire dalla gliceraldeide 3-P si
rigenera il ribulosio 1,5-bisfosfato
(l’accettore di CO2)
5/6 dei trioso fosfati rigenerarano il
Ribulosio 1,5-bisP;
1/6 e’ utilizzato
saccarosio o amido
per
la
sintesi
di
Luce, pianta
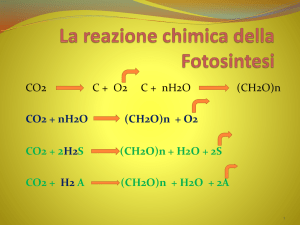
CO 2 + H 2 O (CH 2 O) + O 2
6CO2 + 6H2 O C6H12 O6 + 6O2
n CO2 + 2n H2O + ATP + NADPH → (CH2O)n + n O2 + n H2O
n dipende dallo zucchero considerato, se è il glucosio:
6 CO2 + 12 H2O + 18 ATP + 12 NADPH
C6H12O6 + 6 O2 + 6 H2O + 18 ADP + 18 Pi +12 NADP
Regolazione del Ciclo di Calvin
5 enzimi regolati alla luce
1. Rubisco;
2. NADP:gligeraldeide-3-P deidrogenasi;
3. Fruttosio 1,6-difosfato fosfatasi;
4. Sedeptulosio-1,7-difosfato fosfatasi;
5. Ribulosio-5-fosfato chinasi
Questi enzimi tranne la Rubisco sono attivati alla luce
tramite il sistema ferredossina-tioredossina
il sistema ferredossina-tioredossina
Richiede la formazione di un complesso carbammato-Mg2+
Durante la fotosintesi:
>pH
>conc. Mg2+
pH
Mg2+
Alcune peculiarità della
prima reazione della fase
oscura
Carbossilazione
Rubisco
(PGA)
RUBISCO
Km(CO2)= 12 µM, Km(RUBP)=60 µM, Km(O2)=250 µM
La RUBISCO catalizza anche la reazione di ossigenazione
al ciclo
di Calvin
Fotorespirazione
o
Il ciclo (C2) per l’ossidazione
fotorespiratoria del carbonio
Il ciclo C2
coinvolge tre
differenti
organelli
cellulari
1. Rubisco lega O2
2. 2-fosfoglicolato> glicolato
6. 3-fosfoglicerato
3. (l’attività dei perossisomi è di
rimuovere H dai composti organici
consumando O2 e rimuove H2O2)
•glicolato> gliossilato
•Gliossilato+glutammato>
glicina + a-chetoglutarato
4. 2 molecole di glicina> serina
5. Serina+ achetoglutarato>
idrossipiruvato
>glicerato
Il ciclo per l’ossidazione fotorespiratoria del
carbonio
(o C2) agisce come un’operazione di
salvataggio per recuperare (in parte) la CO2 persa
durante la reazione di ossigenazione della rubisco.
In totale, 4 C sono persi dal cloroplasto (2 molecole di
fosfoglicolato convertite in 2 di glicina) ma 3C sono
recuperati come 3 fosfoglicerato (mentre 1 è perso
come CO2). Il ciclo C2 recupera il 75% del carbonio perso dal
ciclo di Calvin sotto forma di acido 2-fosfoglicolico.
Perché esiste il processo fotorespiratorio?
1. Quando le piante si sono evolute (400 mil di anni fa)
la CO2 dell’atmosfera era cosi alta che l’attività
ossigenasica della Rubisco non aveva alcuna
importanza. Quando la CO2 atmosferica è scesa fino
ai livelli attuali, la pressione selettiva ha selezionato
il processo fotorespiratorio
2. E’ possibile che la fotorespirazione sia necessaria
per dissipare l’eccesso di ATP e di potere
riducente generati dalle reazioni luminose, in
condizioni di alta intensita’ luminosa e di bassa
concentrazione di CO2 prevenendo cosi’ eventuali
danni all’apparato fotosintetico (esperimenti su
mutanti di Arabidopsis).
Molte piante non fotorespirano o lo fanno
in modo limitato…
Meccanismi di concentrazione della CO2!!!
Piante C4
mais
Come funzionano le piante C4?
Il primo prodotto di fissazione della CO2 non era
un composto C3 (PGA) ma acido ossalacetico, un
composto C4.
Anatomia della foglia
Es: Frumento
Arabidopsis
Pomodoro
Pesco
Es: Mais
Canna da zucchero
Sorgo
Piante C4
Varianti del ciclo fotosintetico C4
3 varianti che prendono il nome dall’enzima che
catalizza la decarbossilazione
enzima malico dipendente dall’ NADP e’ tipico
del mais, canna da zucchero e sorgo.
enzima malico dipendente dall’ NAD e’ tipico del
panico, amaranto, portulaca.
fosfoenolpiruvato carbossichinasi e’ tipico del
miglio (Panicum maximum).
Meccanismi di concentrazione della CO2:
Il Metabolismo Acido delle Crassulacee
(CAM)
Caratteristico delle piante che vivono in ambienti
desertici.
Separazione temporale dell’apertura
e chiusura degli stomi.
NADPH NADP+
Buio
Luce
Il metabolismo CAM è regolato dalla fosforilazione della
PEP carbossilasi
Considerazioni ecofisiologiche
Metabolismo C4
Adattamento alle alte temperature
In climi caldi e secchi, il ciclo C4 riduce la fotorespirazione e la
perdita di acqua
Due caratteristiche del ciclo C4 evitano gli effetti deleteri di un
aumento della temperatura sulla fotosintesi
1. l’affinità della fosofenolpiruvato carbossilasi (verso la CO2 nella
forma HCO3-) è cosi alta che è satura a conc. atmosferiche. Inoltre,
l’O2 non è un competitore della reazione (ciò consente di ridurre
l’apertura stomatica e quindi la traspirazione).
2. La fotorespirazione viene soppressa dall’accumulo di CO2
nella cellule della guaina del fascio
Attivita’ fotosintetica a luce saturante
in funzione della temperatura
La percentuale di specie C4 tende ad aumentare dalle
aree piu’ fresche e umide fino alla preponderanza nelle
regioni piu’ calde.
In termini di produttivita’ le C4 sono in genere piu’
efficienti a illuminazioni e a temperature alte.
La richiesta energetica delle C4 e’ superiore a quella del
ciclo C3.
Sono le condizioni ecologiche degli ambienti caldi a
rendere competitive le piante C4.
Rapporto traspirazione
Rapporto traspirazione
L'efficacia delle piante nel limitare la perdita d'acqua e
nello stesso tempo permettere l'assorbimento di CO2
per la fotosintesi viene espressa tramite il rapporto di
traspirazione:
Rapp. trasp.= moli H2O trasp/moli CO2 fissata
per le piante C3, Rapp. trasp.= circa 500!
dovuto principalmente:
1. il gradiente di concentrazione del vapor d'acqua è
circa 50 volte più elevato di quello per CO2; CO2 è solo
0,03%
2. CO2 più grande dell'H2O, Ds minore.
3. CO2 deve percorrere una via di diffusione più lunga e
complessa prima di essere assimilata dal cloroplasto
(membrana plasmatica, citoplasma, membrana del
cloroplasto)
Le piante C3, Rapp. trasp.= circa 500
Le piante C4, Rapp. trasp.= circa 250
Le CAM, Rapp. trasp.= circa 50
Sintesi di amido e saccarosio
Sia l’amido che il saccarosio sono sintetizzati
dal trioso fosfato generato dal ciclo di Calvin.
Il saccarosio e’ traslocato dal floema in
tutta la pianta.
L’amido e’ un carboidrato stabile ed insolubile
di riserva presente in tutte le piante.
La sintesi del saccarosio e dell’amido sono
reazioni competitive
• Il traslocatore di fosfato catalizza lo spostamento di Pi
e del trioso fosfato nelle direzioni opposte fra il
cloroplasto e il citosol
• Una bassa concentrazione citosolica di Pi limita
l’esportazione nel citosol del trioso fosfato e quindi
favorisce la sintesi di amido (che si accumula nei
cloroplasti –amido primario)
• Un’alta concentrazione citosolica di Pi promuove
l’esportazione nel citosol del trioso fosfato favorendo la
sintesi di saccarosio e l’inibizione di amido
Su chi agisce il Pi o il trioso fosfato per regolare la
biosintesi di amido e saccarosio?
Regolando l’attività di alcuni enzimi coinvolti nella loro
biosintesi.
Es: ADP glucosio pirofosforilasi (enzima chiave nella
biosintesi di amido) è inibito dal Pi.
Glucosio 1-fosfato + ATP -> ADP-glucosio + H2P2O7-2
La sintesi del saccarosio e dell’amido sono
reazioni competitive
ADP glucosio pirofosforilasi
Componenti dell’amido