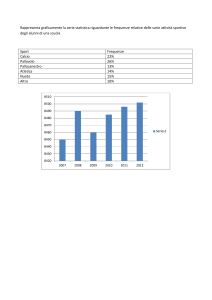
laboratorio evoluzione
DERIVA
[Eigen M. – Winkler R. (1975),
Il gioco, Adelphi Milano 1986
Luigi Luca Cavalli Sforza,
L’evoluzione della cultura,
Codice Torino 2004]
Marcello Sala
Anemia falciforme
malattia ereditaria
dovuta a una mutazione
in un gene per l’emoglobina
eritrociti con vita dimezzata
Soggetti omozigoti senza mutazione: sani
soggetti omozigoti con mutazione: gravemente malati
soggetti eterozigoti: forme più lievi di malattia
Soggetti con a.f. sono più frequenti in zone malariche:
perché?
Il Plasmodium falciparum agente della malaria
non riesce a riprodursi negli eritrociti falciformi
l’anemia protegge dalla malaria
Esempio di processo di selezione naturale
nonostante il carattere in sé sia svantaggioso
Negli USA
lo 0,04%
della popolazione
soffre di acromatopsia
malattia ereditaria
Nell’atollo di Pigelap (Micronesia)
la percentuale è del 5%
A cosa è dovuta
questa anomala frequenza?
L’isola nel 1775 venne devastata da un tifone
la popolazione fu ridotta a 20 persone
tra di esse una era portatrice della malattia (1/20 = 5%)
“… sono convinto che la selezione naturale
è stata la causa principale, ma non l’unica,
delle modificazioni.”
C. Darwin L’origine delle specie
per mezzo della selezione naturale, 1859
GIOCO A
Regole:
• scacchiera 6x6
• pedine di due tipi diversi
• si riempie la scacchiera di un numero uguale
di pedine dei due tipi messe a caso
• a ciascuna faccia di una moneta si assegna uno dei due tipi
• si lancia la moneta sorteggiando così uno dei due tipi
• una pedina qualsiasi di quel tipo viene sostituita
con una dell’altro tipo
• il gioco finisce quando non ci sono più pedine di un tipo
perché non ci può essere più “sostituzione”
Le regole riguardano i cambi di stato delle singole caselle
non prevedono lo sviluppo della situazione sull’intera scacchiera
Versione più astratta del gioco:
le frequenze dei due tipi di pedine
non dipendono dalla collocazione sulla scacchiera
si può eliminare la rappresentazione spaziale
A ogni mossa
- contare le pedine dei due tipi
- estrarre a sorte un tipo (O - X)
- effettuare la “sostituzione” ovvero
sottrarre una unità al tipo estratto e
aggiungere una unità all’altro tipo
- contare le pedine dei due tipi
IL SOFTWARE
si assegna un numero progressivo a ogni pedina di un tipo
poi, esaurite le n pedine di questo, a quelle dell’altro
i numeri ≤ n corrispondono al primo tipo, > n al secondo
si creano così una “zona rossa” e una “zona verde”
Lancio dei dadi sostituito dall’estrazione di un numero
estraendo a caso tra 1 e il totale, il numero può
cadere ella zona rossa o nella zona verde
Se cade nella zona rossa
si aumenta di 1 il numero di pedine rosse e
si diminuisce di 1 quello delle pedine verdi
Viene ricalcolata la frequenza dei due tipi
Alla fine il software produce una striscia con
zona rossa e verde proporzionali alle frequenze
Le strisce una sotto l’altra riproducono le “mosse” successive
Esempio tipico
numero pedine
0
5
10
15
20
25
1
6
11
16
21
Che cosa prevedete
che succederà
alle frequenze dei due tipi?
31
36
41
46
mosse
Dinamica: le frequenze
“oscillano” fortemente
Non apparenti tendenze:
sviluppo casuale
26
51
56
61
66
71
76
81
Perché succede così?
86
91
96
L'aumento/diminuzione
del numero di pedine di un tipo in ogni fase
è indipendente dalla sua frequenza
E se all’inizio le frequenze dei due tipi sono diverse?
30
35
Esempio tipico
E se all’inizio le frequenze
dei due tipi sono diverse?
Stessa dinamica:
le frequenze
“oscillano” molto,
Tanto che a volte
uno dei due tipo
“si estingue”
Perché succede così?
Se si è più vicini a un estremo più facilmente
un’oscillazione forte può arrivare alla frequenza 0
6
5
GIOCO B
4
Regole:
3
• scacchiera 6x6 e pedine di due tipi diversi 2
• si riempie la scacchiera di un numero uguale 1
(18) di pedine dei due tipi messe a caso
1 2 3 4 5 6
• si lanciano 2 dadi di due colori: uno determina l’ascissa
l’altro l’ordinata della casella
• la pedina in quelle coordinate “si riproduce”
cioè una pedina dello stesso tipo, presa dalla riserva,
dovrà essere messa sulla scacchiera
• per liberare un posto si lancia la moneta:
una pedina qualsiasi del tipo corrispondente viene tolta
•il gioco finisce se uno dei due tipi scompare dalla scacchiera
Le regole riguardano i cambi di stato della singola casella
non prevedono lo sviluppo della situazione sull’intera scacchiera
Su computer versione più astratta
eliminando la rappresentazione spaziale
Il software esegue le estrazioni a caso e, a ogni mossa,
calcola le frequenze dei due tipi di pedine
che non dipendono dalla collocazione sulla scacchiera
Che cosa prevedete che succederà
alle frequenze dei due tipi?
Esempio tipico
Oscillazioni meno forti
ma tendenze progressive
fino all’estinzione
Perché succede così?
Pertinente la frequenza:
più pedine di un tipo
più probabile che
vengano sorteggiate
il "tasso di riproduzione"
aumenta con la frequenza
una differenza iniziale
si amplifica
L’eliminazione per far posto alla nuova pedina non bilancia
questa dinamica, perché è del tipo “moneta” e quindi colpisce
in modo uguale i due tipi indipendentemente dalla loro frequenza
“deriva” (Sewall Wright 1932)
Scacchiera più grande (30x30): previsioni?
Sviluppo più “lento” ma ugualmente “deriva”
Esempio tipico
numero pedine
0
50 100 150 200 250 300 350 400 450 500 550 600 650 700 750 800 850 900
1
6
11
16
21
26
31
36
41
mosse
46
51
56
61
66
71
76
81
86
91
La deriva è legata
alla disparità
delle frequenze
di partenza
o al numero
di elementi?
È sensibile al
numero di elementi
96
Esempi tipici
Scacchiera più piccola (4x4): previsioni?
Meno elementi dinamica più veloce fino all’estinzione
la deriva dipende in modo sensibile dalla scarsità di elementi
Esempio tipico
Deriva: dinamica
puramente statistica
dipende solo dalle frequenze
un tipo si estingue o
diventa dominante sull’altro
senza alcuna causa selettiva
[Ferraguti – Castellacci, Evoluzione. modelli e processi, Pearson 2011]
alleli fissati
alleli persi
Variazione delle
frequenze di
coppie di alleli
a partire da
un valore 0,5
in 50 simulazioni
di popolazioni di
diversa dimensione
Cosa succede se c’è anche
una pressione selettiva?
Selezione 2 a 1 a favore
del verde: previsioni?
Esempi tipici
Aumenta il favorito
dalla selezione ma
a volte anche lo sfavorito
effetto di deriva
Selezione 5 a 1 a favore del verde: previsioni?
Esempi tipici
Selezione 5 a 1 a favore del verde
con meno frequenza iniziale
Sotto una certa frequenza la pressione selettiva non riesce
a invertire la deriva sfavorevole
Tra gli indiani del SudAmerica il gruppo sanguigno 0 è dominante
mentre nella popolazione dell’Asia, Europa e Africa
i gruppi A, B, 0
sono in proporzioni
poco diverse tra loro
perché?
America colonizzata
da Homo sapiens
da (25.000) 18.000 anni fa
popolazioni poco numerose
dall’Asia orientale
(stretto di Bering
Isole Aleutine collegate
dai ghiacci, navigazione
attraverso l’Oceano)
passaggi stretti e difficili “collo di bottiglia”
Migrazione in territorio “vuoto”
popolazione abbastanza piccola
(gruppi di famiglie persone imparentate
corredi genetici più omogenei)
distribuzione delle diverse forme
di un tratto ereditario (alleli di geni)
lontana dall’equilibrio della popolazione intera:
“effetto del fondatore”
Deriva il disequilibrio tende ad accentuarsi
(se la popolazione rimane isolata,
non viene contrastato dall’ibridazione)
Ayala, Scientific American, sett. 1978
VV
VL
LL
Gel elettroforetico: malatodeidrogenasi (enzima ossidante)
da campioni di 22 individui di Drosophila equinoxiails
perché tre tipi di polipeptidi?
L’enzima consiste di due catene che si combinano
spontaneamente dopo essere state sintetizzate
Due forme varianti prodotti di due alleli:
uno che migra velocemente (V) uno lentamente (L)
gli omozigoti producono solo una forma (o VV o LL)
gli eterozigoti ne producono tre: VV, LL e VL
Perché esistono tre varianti dello stesso enzima
nella stessa popolazione?
Zuckerkandl, Scientific American, 213, p 110, 1965
Inizio delle sequenze delle emoglobine e in alcune specie
evidenziate le posizioni occupate dallo stesso AmminoAcido
Come sono possibili così estese variazioni (mutazioni)
in uno stesso enzima (= stessa funzione)?
Zuckerkandl, Scientific American, 213, p 110, 1965
26 siti di AA sono uguali in tutte le
emoglobine note (a sinistra)
11 di essi sono uguali anche nelle mioglobine
(a destra con numeri diversi)
Che cosa ci dicono questi dati?
I siti invarianti sono importanti
per la struttura e la funzione del polipeptide
Le mutazioni colpiscono parti della proteina poco funzionali
“La recente scoperta della “degenerazione”
del codice, e cioè l’esistenza di due o più triplette
che codificano per lo stesso aminoacido,
sembra suggerire che le mutazioni neutrali
potrebbero essere non così rare come si pensava.
In aggiunta, alcune sostituzioni di aminoacidi
in una catena polipeptidica possono avere
un effetto assai scarso sull’attività biologica della proteina,
rafforzando così l’idea di mutazioni neutrali o quasi.”
M. Kimura, 1968
“… per la maggior parte i geni mutanti,
individuati solo con i procedimenti chimici
della genetica molecolare
sono neutrali di fronte alla selezione,
cioè, sotto l’aspetto adattativo,
non sono né più né meno vantaggiosi
dei geni che sostituiscono; a livello molecolare
la maggior parte dei cambiamenti evolutivi è provocata dalla
deriva casuale di geni mutanti
che sono equivalenti di fronte alla selezione.”
M. Kimura 1980
le mutazioni “neutrali” per la variazione fenotipica
sfuggono al processo di selezione naturale
si accumulano per deriva
Ridley, Evolution, 2004, p. 165-67
I tassi di evoluzione
delle emoglobine
sembrano essere costanti
Cosa suggerisce
questo dato?
Le mutazioni neutrali
costituiscono un
“orologio molecolare”
(Zuckerkandl e Pauling 1962)
Come si possono con l’orologio molecolare
misurare i tempi dell’evoluzione?
Se in uno stesso punto della catena di una proteina
in specie diverse ci sono AA diversi
una mutazione è avvenuta su una delle due linee
che provengono dall’antenato comune
Se le differenze si sono accumulate a ritmo costante
il numero di differenze AmminoAcidiche fra le due specie
deve essere proporzionale al tempo della loro divergenza
Perché devono essere mutazioni neutrali?
Il ritmo del loro accumulo non dipende dalla selezione
naturale (e quindi dal mutare delle condizioni ambientali)
Futuyma, Evolutionary Biology, 1986, p. 452
.
Ricostruzione della
filogenesi delle
mioglobine (Mb) e
delle emoglobine (Hb)
di un mollusco (Aplysia)
e di alcuni vertebrati
basata su una analisi
delle sequenze
di AmminoAcidi
I numeri lungo i rami
si riferiscono alle
sostituzioni di nucleotidi
stimate dalle differenze
di amminoacidi
I tempi di divergenza
dei taxa sono basati
su reperti fossili
)
Diverse visioni del valore selettivo delle mutazioni da parte
dei selezionisti (sinistra) e dei neutralisti (destra)
Ridley, Evolution,
2006, p. 147
Coefficiente di selezione = 0: mutazioni neutrali
Le due teorie sono conflittuali e alternative?
Si tratta piuttosto di valutare caso per caso
quanto la situazione è effetto della selezione naturale
e quanto della deriva (molecolare o popolazionale)
A
Una parte
della
popolazione
attraversa
una barriera
geografica
preesistente
B
Una nuova
barriera
geografica
si forma nel
territorio della
popolazione
e la divide
Quali differenze nel processo di speciazione?
Diversa velocità
Selezione o deriva?
Nel caso A
filtro selettivo
più forte
ma anche deriva
Perché il “neutralismo”
è una critica (interna) e una integrazione
al darwinismo?
“Le condizioni ambientali svolgono sicuramente
un ruolo decisivo nel determinare
quali fenotipi vengono selezionati;
Ma la selezione darwiniana, o positiva,
non tiene in gran conto in che modo
questi fenotipi siano determinati dai genotipi.
Le leggi che governano l’evoluzione molecolare sono chiaramente
diverse da quelle che governano l’evoluzione fenotipica.
Anche se il principio darwiniano della selezione naturale prevale,
a livello fenotipico, nel determinare l’evoluzione,
al livello più basso della struttura interna del materiale genetico,
una grande parte dei cambiamenti evolutivi
è promossa dalla deriva casuale.
M. Kimura, 1980
Un gruppo di 4 ♂ di cognomi A, B, C, D
e 4 ♀ di cognomi E, F, G, H
migra e fonda un nuovo villaggio
Si formano coppie: ♂A-♀E ♂B-♀F ♂C-♀G ♂D-♀H
Se ogni coppia genera un ♂ e una ♀
quanti cognomi sono presenti nella nuova generazione?
4 (da 8): A, B, C, D, rappresentati ciascuno da un ♂ e una ♀
I giovani formano coppie con partner di cognome diverso:
♂A-♀B
♂B-♀C ♂C-♀D ♂D- ♀A
Se ogni coppia genera un ♂ e una ♀
quanti cognomi sono presenti nella nuova generazione?
4: A, B, C, D, rappresentati ciascuno da un ♂ e una ♀
…
I giovani formano coppie con partner di cognome diverso:
♂A-♀B
♂B-♀C ♂C-♀D ♂D- ♀A
Se la prime due coppie generano 2 ♂
e le altre generano 2 ♀
quanti cognomi sono presenti nella nuova generazione?
4: A e B rappresentati da 2 ♂, C e D da 2 ♀
I giovani formano coppie con partner di cognome diverso:
♂A-♀C
♂A-♀D ♂B-♀C ♂B- ♀D
Se ogni coppia genera un ♂ e una ♀
quanti cognomi sono presenti nella nuova generazione?
2 (da 4): A e B rappresentati ciascuno da 2 ♂ e 2 ♀
Perché nei paesi piccoli senza immigrazione pochi cognomi?
“Estinzione” dei cognomi per effetto di deriva
I mitocondri contengono alcuni anelli DNA
per un totale di solo poche decine di migliaia di nucleotidi
residuo di simbiosi
Nella fecondazione i mitocondri provengono tutti
dalla cellula uovo materna
il DNA mitocondriale è ereditato per linea materna
Perché il DNA mitocondriale è un buon tracciante
per l’evoluzione nella popolazione umana?
se la popolazione iniziale è poco numerosa, stessa dinamica
dei cognomi (estinzione per deriva), ma a favore della ♀
tendenza all’estinzione, nessuna “immigrazione”,
nessuna ricombinazione riduzione della variazione
bassa esposizione a selezione accumulo di mutazioni neutrali
Colori = varianti
genetiche nelle
popolazioni umane
dati da analisi del
DNA mitocondriale
1999 Kenneth K. Kidd, Yale University
Oggi la diversità genetica tra due boscimani
è 5 volte superiore a quella tra
un europeo e un aborigeno australiano:
come si spiega?
1999 Kenneth K. Kidd, Yale University
Con la migrazione
la variazione diminuisce
Se a migrare è
un piccolo gruppo di individui,
parte della variazione
della popolazione di partenza
va persa per deriva genetica