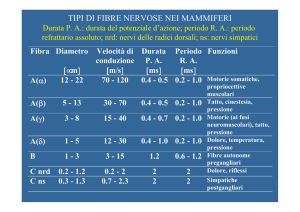
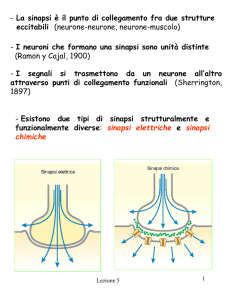
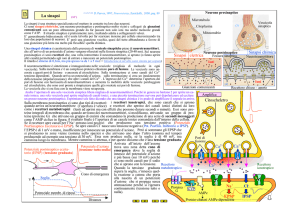
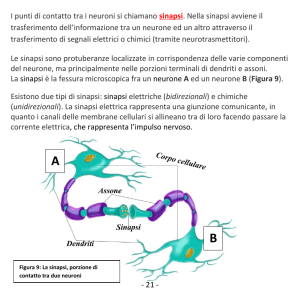
LE SINAPSI
Al fine di rendere più efficiente la comunicazione chimica di tipo paracrina l'evoluzione ha
"prodotto" le sinapsi, costituite da due microaree di membrana, appartenenti rispettivamente a due
cellule diverse, dette elemento presinaptico ed elemento postsinaptico. Le due aree sono separate da
uno spazio intersinaptico, dello spessore di circa 30 nm o 300 Å (un ångstrom è pari a 0.1 nm).
L'elemento presinaptico è più frequentemente la parte terminale di un assone, leggermente espansa
(viene detto bottone sinaptico), l'elemento postsinaptico è più frequentemente il corpo, i dendriti o il
terminale di un assone di un neurone ( si parla dunque sinapsi interneurali asso-somatica, assodendritica ed asso-assonica) (Fig. 7.1). Esistono però sinapsi cito-neurali, dove l'elemento
presinaptico è una cellula sensoriale (quale la cellula ciliata dell'organo dell'udito) e sinapsi neuromuscolari, dove l'elemento postsinaptico è una cellula muscolare. L'informazione passa
dall'elemento presinaptico a quello postsinaptico, utilizzando un messaggero chimico detto
neurotrasmettitore. Questo è sintetizzato nel corpo neuronale e viene trasportato lungo l'assone sino
al terminale presinaptico, dove è concentrato in vescicole. Viene quindi liberato per esocitosi nello
spazio sinaptico, dove diffonde ed interagisce con recettori posti sulla membrana dell'elemento
postsinaptico (Fig. 7.1, inserto). Le gap junction sono spesso considerate sinapsi elettriche. I due
meccanismi non hanno nulla di comune e sono perciò probabilmente evoluti in modo separato.
Terminali
presinaptici
C
A
Dendriti
Assone
PdA e
trasporto del
neurotrasmettitore
Sinapsi assosomatica
B
Vescicole
Bottone
sinaptico
Sinapsi assodendritica
Recettore
Fig. 7.1. Il neurone A prende contatto, tramite l'assone, con il corpo cellulare ed i dendriti del
neurone B; si costiutiscono così sinapsi asso-somatiche ed asso-dendritiche. Il neurone A è a sua
volta contattato dall'assone C. Nell'inserto è schematizzata l'esocitosi e l'interazione
neurotrasmettitore-recettore.
CENNI SULLA STRUTTURA DEL SISTEMA NERVOSO CENTRALE
Il sistema nervoso centrale (SNC) è costituito dall'encefalo, racchiuso e protetto dal cranio, e dal
midollo spinale, situato all'interno dei corpi delle vertebre (Fig. 6.2); dal SNC si dipartono i nervi
che si distribuiscono a tutto il corpo; questi fanno parte del Sistema Nervoso Periferico.
Cervelletto
Encefalo
Cervello
Tronco
cerebrale
Midollo
spinale
A
Midollo
spinale
Nervi
periferici
B
C
Fig. 7.2. Struttura del sistema nervoso. A, il sistema nervoso, visto posteriormente. B, il sistema
nervoso, visto posteriormente (ingrandimento dell'area tratteggiata in A). C, sezione trasversa del
sistema nervoso centrale, sino alle prime vertebre toraciche.
L'encefalo comprende il cervello e, inferiormente, il tronco cerebrale e il cervelletto. Il tronco
cerebrale è costituito dal mesencefalo, il ponte ed il bulbo, cui segue il midollo spinale. Il
cervelletto è situato inferiormente al cervello, dietro il tronco encefalico (Fig.7.2).
Nel SNC umano abbiamo circa 1011 neuroni, ciascuno dei quali ha migliaia di connessioni
sinaptiche.
MECCANISMI PRESINAPTICI
L'esocitosi del neurotrasmettitore è fortemente favorita dall'arrivo, al terminale presinaptico, di un
potenziale d'azione. Il terminale non è fornito di canali al Na+(V), cosicchè il potenziale d'azione
non insorge nel terminale ma lo depolarizza, con lo stesso meccanismo con cui depolarizza le aree
vicine nel propagarsi lungo la fibra nervosa. Il terminale possiede un tipo di canale al Ca2+
voltaggio-dipendente (Ca2+(V)), che si apre in seguito alla depolarizzazione evocata dal potenziale
d'azione. L'ingresso di Ca2+ scatena l'esocitosi, come meglio vedremo successivamente. Il terminale
presinaptico ha canali al K+ attivati dal Ca2+ intracellulare (K+(Ca2+)), cosicchè l'aumento della
[Ca2+]i attiva questi canali, che ripolarizzano il terminale e chiudono i canali al Ca2+(V).
Contemporaneamente, lo scambiatore Na+/Ca2+ e la Ca2+ATPasi espellono il Ca2+: tutti questi
meccanismi rendono molto breve (dell'ordine di qualche ms) l'aumento della [Ca2+]i e la durata
dell'esocitosi.
Esocitosi ed endocitose delle vescicole sinaptiche
Il processo dell'esocitosi è controllato da alcune proteine, inserite sia nella membrana della
vescicola sia nella membrana del terminale presinaptico. La Sintaxina ( associata a MUNC18-1) e
la SNAP-25 sono inserite nella membrana plasmatica, mentre la Sinapto-brevina è inserita nella
membrana vescicolare. Queste proteine sono complessivamente denominate proteine SNAREs (Fig.
6.3). L'esocitosi è preceduta dalla fase di Docking, nella quale la vescicola si avvicina alla
membrana plasmatica. Questa fase, poco nota nel suo meccanismo molecolare, è reversibile. La
fase successiva è detta Priming: la Sinapto-brevina interagisce con la Sintaxina e la SNAP-25,
avvicinando la vescicola alla membrana plasmatica. La fusione avviene con la cooperazione della
Sinapto-tagmina, una molecola Ca2+-sensibile inserita nalla membrana della vescicola. L'ingresso di
Ca2+ attiva la Sinaptotagmina (non schematizzata in figura), che insieme alle proteine SNAREs
induce la fusione della membrana e l'apertura della vescicola verso l'ambiente extracellulare (Fig.
7.3).
Ca2++ e
sinaptotagmina
Fig. 7.3. Alcune proteine connesse con l’esocitosi Ca2+-dipendente.
L’esocitosi deve essere seguita dalla rimozione della membrana vescicolare, che avviene tramite un
processo mediato da clatrina. Le vescicole accumulano poi nuovamente il mediatore, mediante un
meccanismo di trasporto attivo, ed entrano a far parte del pool di riserva, che abbandoneranno con il
processo di docking (Fig. 7.4).
Accumulo del
neurotasmettitore
Rimozione
della Clatrina
Pool di deposito
Docking
Priming
Fusione
Liberazione del
neurotrasmettitore
Fig. 7.4. Ciclo delle vescicole del terminale presinaptico.
Endocitosi
Clatrinadipendente
MECCANISMI POSTSINAPTICI
La risposta postsinaptica è funzione del tipo di recettore di membrana che viene attivato dal
neurotrasmettitore. Questo può attivare un sistema di messaggeri intracellulari, o aprire dei canali
ionici attivabili da molecole chimiche.
Esamineremo ora brevemente la risposta postsinaptica nella sinapsi neuromuscolare e nelle sinapsi
del sistema nervoso centrale.
LA SINAPSI NEUROMUSCOLARE
La contrazione della fibra muscolare scheletrica è controllata da un neurone detto motoneurone, il
cui assoone prende contatto sinaptico con la fibra muscolare scheletrica. Quando un PdA raggiunge
il terminale presinaptico, viene liberata Ach che interagisce con i recettori colinergici nicotinici
presenti sulla membrana plasmatica della fibra muscolare (Fig. 7.5).
Guaina mielinica
PdA della fibra nervosa
Terminale
presinaptico
+
Na (V): questi canali
sono presenti sino
all’inizio del terminale
presinaptico
Ach
Na+(V)
K+(V)
PdA della fibra
muscolare
Membrana
del terminale
+
Ca2+(V)
+
Riassorbimento
di colina
+
PdA della fibra
muscolare
Fibra muscolare
scheletrica
++
++ Ach-esterasi
Canali nicotinici: entra Na+ ed
esce K+; si genera un flusso netto
di cariche positive in ingresso.
Fig. 7.5. Processi connessi con la trasmissione sinaptica nella sinapsi neuro-muscolare. L'arrivo di un PdA al
terminale presinaptico genera sempre un PdA nella fibra muscolare scheletrica; questo innesca la contrazione
della fibra.
Nella sinapsi neuromuscolare il PdA libera circa 300 vescicole, che depolarizzano sino a circa -20
mV la membrana plasmatica della fibra muscolare situata di fronte all’elemento pre-sinaptico, priva
di Na+(V). Questa depolarizzazione, con l'usuale meccanismo, depolarizza le zone della membrana
plasmatica vicine all’area postsinaptica, generando due PdA, che si propagano in direzione opposta
lungo tutta la fibra.
L’Ach viene rapidamente idrolizzata dall’Ach-esterasi, cosicchè il mediatore resta nella fessura
sinaptica per circa 5 ms; la colina viene riassorbita attivamente, ed utilizzata per sintetizzare, nel
citosol, Ach. Questa viene poi riassorbita attivamente dalle vescicole.
LE SINAPSI NEL SNC
La sinapsi neuromuscolare è particolarmente semplice: la fibra muscolare scheletrica riceve una
sola afferenza sinaptica ed ogni volta che arriva un PdA al terminale sinaptico la liberazione di
mediatore è talmente elevata che insorge sempre un PdA nell’elemento postsinaptico.
Nel SNC l’organizzazione sinaptica è molto più complessa. Un neurone riceve migliaia di afferenze
sinaptiche, che liberano differenti tipi di neurotrasmettitori, in grado di attivare messaggeri
intracellulari o aprire canali ionici. Se l’Ach e l’acido Glutammico interagiscono con recettoricanale, il flusso ionico depolarizzerà la cellula, favorendo l’insorgenza di un PdA e la sinapsi viene
detta eccitatoria. Se il mediatore è il GABA, l’apertura dei canali al Cl- tende ad inibire
l’insorgenza del PdA e la sinapsi viene detta inibitoria.
L’elemento postsinaptico riceve sia afferenze eccitatorie che inibitorie ed effettua un processo di
sommazione algebrica delle afferenze: se prevale l’eccitazione potrà insorgere un PdA
nell’elemento postsinaptico. Si noti che nelle sinapsi del SNC un singolo PdA presinaptico non è
mai in grado di generare un PdA nell’elemento postsinaptico.
LA SOMMAZIONE POSTSINAPTICA
In Fig. 7.6, A, è rappresentato un neurone che riceve due afferenze eccitatorie (giallo) ed
un’afferenza inibitoria (rossa). L’arrivo di un PdA presinaptico in una sinapsi eccitatoria libera il
neurotrasmettitore che apre dei canali ionici nell’elemento postsinaptico. Sebbene il mediatore
venga rimosso od inattivato nel giro di pochi ms, la depolarizzazione evocata dall'apertura dei
canali ionici, denominata potenziale post-sinaptico eccitatorio (PPSE) ha una durata ben maggiore
(circa 20-30 ms; Fig. 7.6, B).
Se al terminale presinaptico arriva un secondo PdA prima che la depolarizzazione evocata dal primo
PdA sia estinta, si avrà una sommazione delle due depolarizzazioni. Lo stesso processo può
intervenire nel caso di sinapsi inibitorie, che tendono ad iperpolarizzare la cellula (in questo caso si
ha un potenziale postsinaptico inibitorio, o PPSI). Questo tipo di sommazione, dovuta all’azione di
un singolo terminale presinaptico al quale arrivano in rapida successione più PdA, è detta
sommazione temporale.
La sommazione può avvenire anche in seguito all’attivazione di due terminali distinti. Se un
terminale viene attivato ed un altro terminale viene attivato prima che sia terminato l’effetto del
primo, avremo la sommazione, detta in questo caso spaziale. La sommazione spaziale consente la
sommazione di effetti eccitatori (depolarizzanti) ed inibitori (iperpolarizzanti).
In Fig. 7C viene rappresentato il processo di sommazione. La soglia è –40 mV. Un singolo PPSE
non raggiunge la soglia, mentre 2 PPSE la superano, generando un PdA. La traccia rossa
rappresenta un PPSI; la traccia arancione è la somma tra un PPSE ed un PPSI. La figura, disponibile
in rete, presenta errori ed imprecisioni. I processi di sommazione sono meglio rappresentati nelle
figure 7.7, 7.8 e 7.9.
Il neurone postsinaptico effettua dunque la somma algebrica delle afferenze eccitatorie ed
inibitorie, generando un PdA se viene superata la soglia: è, di fatto, la struttura "decisionale" del
SN.
A
B
(mV)
Microelettrodo - 85
Vm
per la misura
di Vm
Corpo
cellulare
- 90
Dendriti
0
40 (ms)
Assone
C
soglia
Fig. 7.6. A, viene schematizzato un neurone che riceve due afferenze eccitatorie (gialle) ed una
afferenza inibitoria (rossa). B, potenziale postsinaptico eccitatorio. C, viene illustrato il processo di
sommazione, con notevole imprecisione (la curva arancione rappresenta la somma di un'afferenza
eccitatoria e di un'afferenza inibitoria). Il pannello C è disponibile in rete.
Potenziale di m em brana
-20
-30
-40
Soglia= - 50 mV
Vm
-50
-60
-70
-80
-90
0
25
50
75
100
125
150
175
200
Tempo (ms)
Fig. 7.7. Due PPSE distinti. Le freccie indicano l'arrivo di un PdA nell'elemento presinaptico. Da questo
grafico non è possibile capire se trattasi di sommazione spaziale o temporale.
Potenziale di m em brana
-20
-30
-40
Soglia= - 50 mV
Vm
-50
-60
-70
-80
-90
0
50
100
150
200
250
300
350
400
Tem po (m s)
Fig. 7.8 Due PPSE che si sommano. Le freccie indicano l'arrivo di un PdA nell'elemento presinaptico.
L'arrivo di un terzo PPSE può far superare la soglia.Da questo grafico non è possibile capire se trattasi di
sommazione spaziale o temporale.
Potenziale di m em brana
-20
-30
Vm
-40
Soglia= - 50 mV
-50
-60
-70
-80
-90
0
50
100
150
200
Tempo (ms)
7.9 Un PPSI si somma ad un PPSE. Le freccie indicano l'arrivo di un PdA nell'elemento presinaptico.
Trattasi ovviamente di sommazione spaziale.
Si ricordi che il canale aperto dal Glutammato di tipo NMDA è permeabile solo se l'elemento
postsinaptico è depolarizzato. Se questo è dotato sia di recettori-canale NMDA che di recettoricanali AMPA, l'apertura dei canali AMPA depolarizzerà la cellula, rimuovendo il blocco da Mg2+
dei canali NMDA. A depolarizzare l'elemento postsinaptico può però provvedere qualsiasi sinapsi
eccitatoria, quale una sinapsi colinergica nicotinica. Il canale NMDA sarà quindi pervio se viene
liberato sia Acido Gluttammico sia, da un'altro terminale, Ach. Il canale NMDA può essere
comparato all'operatore booleano AND, che genera 1 (vero) se le condizioni in ingresso sono tutte 1
(vere): il canale NMDA genera 1 (canale aperto) se è attivato dall'acido Glutammico (prima
condizione) AND se la cellula è depolarizzata (seconda condizione).
L'INIBIZIONE PRESINAPTICA
Nell'inibizione presinaptica il terminale sinaptico di una fibra nervosa prende contatto con il
terminale sinaptico di un'altra fibra nervosa ed è in grado di inibirne la liberazione di mediatore
(Fig. 7.10). Facendo riferimento a questa figura, supponendo che B depolarizzi C, l'attivazione di A
inibisce la depolarizzazione di C. Si può osservare che se A inibisse direttamente C, tenderebbe a
limitare l'effetto eccitatorio di tutte le afferenze depolarizzanti che convergono su C; nell'inibizione
presinaptica A inibisce unicamente l'effetto depolarizzante esercitato dal terminale B. Il
meccanismo dell'inibizione presinaptica non è oggetto di trattazione.
B
A
C
Fig. 7.10. Inibizione presinaptica. L'attivazione del terminale presinaptico A inibisce la liberazione di
mediatore da parte del terminale presinaptico B.
LA GENESI DEL PdA NELL'ELEMENTO POSTSINAPTICO
Come precedentemente accennato, la depolarizzazione evocata dalle afferenze sinaptiche si
trasmette al cono assonico, dove vi è una forte densita di Na+(V) e K+(V) delayed. Il PdA insorge
dapprima nel cono assonico perchè l'elevata densita di Na+(V) rende la soglia molto inferiore a
quella presente nel soma. il PdA, che si propaga in direzione ortodromica, allontanandosi dal corpo
cellulare (Fig. 7.11). ma, come già spiegato, non può tornare indietro.
La forte depolarizzazione del PdA è poi in grado di depolarizzare oltre la soglia anche il soma del
neurone e può invadere il corpo cellulare, muovendosi in direzione antidromica. Come già
sottolineato, non può però tornare indietro.
+++
++
Terminali
presinaptici
++
i
Cono assonico
Dendriti
PdA
i
A
++
+
Assone
PdA e
trasporto del
neurotrasmettitore
Vescicole
Terminali
presiptici
Terminale
presinaptico
Fig. 7.11. La depolarizzazione provocata nella cellula A dalle afferenze sinaptiche depolarizza il suo cono
assonico, evocando un PdA. Le 2 frecce affiancate da una "i" indicano una corrente (movimento di ioni) che
depolarizza il cono assonico. Il PdA si propaga lungo l'assone attivando la liberazione di neurotrasmettitore
dai terminali presinaptici in contatto con la cellula B. Si noti come il neurite effettua anche il trasporto del
neurotrasmettitore.