Francesco Fiume
Fisiologia generale
delle piante da orto
Fiume Francesco
Fisiologia generale delle piante da orto
Autore: Francesco Fiume
Titolo: Fisiologia generale delle piante da orto
Impostazione e grafica: Francesco Fiume
Elena Ciscognetti
Impaginazione: Francesco Fiume
Finito di stampare: 15 gennaio 2010
Stampato a Pontecagnano (Salerno)
Presso la Sezione di Biologia, Fisiologia e Difesa
dell’Istituto Sperimentale per l’Orticoltura
Via Cavalleggeri, 25 – 84098 Pontecagnano (Salerno)
Direttore della Sezione: dott. Francesco Fiume
-2-
Fiume Francesco
Fisiologia generale delle piante da orto
INTRODUZIONE
La fisiologia vegetale è la scienza che si occupa delle manifestazioni della vita delle
piante, allo scopo di comprendere come queste nascono, si nutrono, crescono, si moltiplicano e
si riproducono, reagiscono agli stimoli ambientali attraverso alcune proprietà come la sensibilità
e la motilità, invecchiano e muoiono.
Non è noto come e quando la vita comparve sul nostro pianeta. Le prime impronte della
vita sono state riconosciute in rocce vecchie di circa 3,8 miliardi di anni e furono lasciate da
microscopici organismi viventi che si possono considerare batteri che ricavavano l’energia
necessaria ai loro processi vitali dalla trasformazione di composti dello zolfo, prodotti
dall’attività vulcanica presente sul fondo degli oceani. Questi microrganismi sono di circa un
miliardo di anni più giovani della Terra e non sono stati trovati altri reperti rocciosi più antichi
nei quali è stato possibile individuare tracce di vita in epoca anteriore.
Più tardi, circa 3 miliardi di anni fa, tali organismi acquistarono la capacità di procurarsi
energia direttamente dalla luce solare e di accumulare azoto. Gli organismi più complessi dotati
di organizzazione cellulare eucariotica non si sono evoluti fino a circa un miliardo e mezzo di
anni fa. Per circa 2 miliardi di anni, pertanto, gli organismi unicellulari procariotici, i batteri,
furono le sole forme di vita presenti sul nostro pianeta. Nella figura 1 è possibile osservare i più
antichi fossili costituiti da batteri provvisti di una struttura molto semplice.
Fig. 1 – Sezione ultrasottile al microscopio elettronico dove si osservano le pareti trasversali
delimitanti le singole cellule di batteri uniti in catenelle.
-3-
Fiume Francesco
Fisiologia generale delle piante da orto
Un momento fondamentale nella storia della vita è rappresentato dalla comparsa dei
cianobatteri, o alghe azzurre, che si accrebbero formando sul fondo strutture a strati cavoliformi
analoghe a quelle tuttora visibili al largo delle coste australiane.
I cianobatteri si servirono dell’energia della luce solare per ricavare carboidrati dalla
combinazione chimica dell’acqua con l’anidride carbonica. Tale processo, noto come
fotosintesi, diede come prodotto di scarto l’ossigeno che, a partire da 2,5 e sino a 1,75 miliardi
di anni fa, si legò chimicamente col ferro ossidandolo e formando sul fondo degli oceani
depositi a strati di rocce ferrose.
Il tappeto di alghe azzurre era anche in grado di intrappolare e creare strati di sostanze
quali i fanghi di carbonato di calcio, che formarono le strutture note come stromatoliti. Non
appena tutto il ferro fu legato chimicamente, l’ossigeno resosi disponibile cominciò ad
accumularsi nell’acqua degli oceani e a diffondersi nell’atmosfera; qui costituì uno strato di
ozono, che fece da scudo contro le dannosissime radiazioni ultraviolette provenienti dal sole
che, altrimenti, avrebbero reso impossibile lo sviluppo della vita. Solo pochissimi fossili di
organismi provengono dal Precambriano, anche dal più recente, e per la maggior parte sono
costituiti da piante. Alghe calcaree si diffusero ampiamente nei mari dell’America (Montana,
Alberta) e della Rhodesia; nella selce nera precambriana dell’Ontario e nelle rocce del
Michigan, Minnesota, Inghilterra e Scozia, sono stati trovati primitivi funghi acquatici e alghe.
Gli animali fossili sono rari; è stata scoperta una medusa nel Gran Cañion ed alcune tracce
rinvenute nelle rocce del Montana testimoniano una possibile esistenza di esseri viventi. Un
maggior numero di fossili sono stati ritrovati in Australia, in depositi recentemente scoperti.
Sembra probabile che gli animali del Precambriano avessero il corpo privo di parti di una certa
consistenza e perciò scarsamente conservabili sotto forma di fossile; solo in seguito, una grande
quantità di differenti specie svilupparono parti ossee ed i loro fossili diventarono più comuni. Il
paesaggio dell’epoca precambriana risulta al principio piuttosto desolato: uno sterile deserto di
nuda roccia attorniato da mari poco profondi, da dove hanno inizio le prime forme di vita.
Origine della vita
La comprensione della vita e della sua origine sul nostro pianeta è un problema che
l’uomo ha sempre, invano, cercato di risolvere ed il notevole sviluppo delle diverse discipline
biologiche che si è verificato negli ultimi cento anni ha permesso di affrontare, in modo
sperimentale, l’approccio alla problematica.
Anticamente, in assenza di conoscenze scientifiche e tecnologiche, la comprensione
dell’origine della vita si è dimostrata di una tale difficoltà ed incertezza da ingenerare le più
disparate interpretazioni metafisiche e filosofiche.
Dalla mitologia classica si deduce questa grande confusione, per cui numerosi personaggi
prendevano origine da piante ed animali o addirittura si supponevano vivificati dalla materia
inanimata. Inoltre, la complessità della vita è giudicata tale che, come logica conseguenza,
soltanto un Essere superiore può averla creata e molti scienziati ancora ammettono, in modo
esplicito o implicito, l’esistenza di un creatore intelligente e soprannaturale la cui esistenza non
è razionalmente descrivibile e dimostrabile attraverso una ricerca sperimentale.
La corrente di pensiero che suppone che le funzioni connesse alla vita siano legate ad una
invisibile ed intangibile forza vitale o spirituale è indicata con il termine di vitalismo. Questi
punti di vista, che erano sostenuti fino a circa tre secoli addietro, si concretizzano, per esempio,
nella teoria della generazione spontanea per la quale piante ed animali potevano prendere
-4-
Fiume Francesco
Fisiologia generale delle piante da orto
origine da sostanze in decomposizione e, comunque, da un mondo inanimato. Si credette che le
anguille e molti altri vermi prendessero origine dal fango o dalla terra umida, che i parassiti
derivassero dal sudiciume e le larve della Sarcophaga carnaria, fossero prodotte dalla carne in
via di decomposizione, scambiando, così, l’ambiente favorevole con la causa efficiente.
L’ipotesi della generazione spontanea si esaurì con Francesco Redi, Lazzaro Spallanzani e
Luigi Pasteur, i quali dimostrarono, con metodo sperimentale, rispettivamente per gli insetti, i
protozoi ed i batteri, che la nascita degli organismi viventi deriva da altri viventi. L’approccio
che consente l’applicazione del metodo scientifico attraverso la sperimentazione e la verifica dei
risultati ottenuti è denominato meccanicismo.
Va ancora detto, tuttavia, che i principi della teoria della generazione spontanea vengono
ancora oggi chiamati in causa, sia pure su basi differenti, per spiegare e sostenere l’origine della
vita sul pianeta terra. E’ opinione comune di molti studiosi del problema che, una volta che si è
verificata l’evoluzione polimerica dei composti del carbonio, in determinate condizioni
ambientali di estremo dinamismo è scoccata, dopo infiniti tentativi, la scintilla dell’evoluzione
prebiotica che ha condotto alla comparsa delle prime forme viventi in un mondo senza vita. A
tal proposito, numerose sono le ricerche che hanno tentato e cercano di stabilire non tanto i passi
dell’evoluzione, dal momento che è impossibile la formulazione di qualunque ricostruzione
storica dell’epoca primordiale, ma di stabilire quali possono essere state le caratteristiche
fondamentali per la nascita della vita e di ottenere dati sperimentali sui principi fisici che hanno
spianato la strada al processo della nascita e della prima evoluzione della vita. La conoscenza
della fisica assume un’importanza fondamentale per lo studio dell’energia ed il suo
trasferimento in strutture molecolari da cui si sono poi evolute le prime forme viventi.
D’altra parte, numerosi scienziati, tra i quali S. Arrhenius, a cavallo dei due secoli
precedenti, e F.H Crick, uno degli ideatori, nel 1953, del modello spaziale della doppia elica del
DNA, ammettono che la vita non abbia mai avuto una propria origine e che essa sia sempre
esistita come proprietà della materia. Particolari molecole e macromolecole provenienti dallo
spazio possono moltiplicarsi e colonizzare i corpi celesti soltanto se su questi esistono
condizioni ambientali compatibili con la vita intesa come macchina funzionante.
L’evoluzione prebiotica
I tentativi fatti per imitare la vita sono ancora poca cosa rispetto al tempo di tre miliardi e
mezzo di anni, trascorso dalla sua origine.
Nessuno può dire di sicuro quali fossero le condizioni atmosferiche o terrestri in quel
momento o quale fosse la molecola che oltrepassò la soglia critica tra la chimica organica e la
biologia.
Una delle prime interessanti ricerche che ha avuto come obiettivo l’ottenimento di dati
sperimentali sulle condizioni ambientali che possono aver determinato l’insorgere della vita è
stata quella di S. L. Miller nel 1953, un neolaureato ventitreenne dell’Università di Chicago.
Tra i numerosi e fondamentali problemi che bisogna prendere in considerazione nello
studio dell’origine della vita, vi è il modo in cui possono essersi formate le prime proteine in un
mondo privo di vita. In un apposito pallone, con all’interno due elettrodi, fu introdotta una
miscela di metano, ammoniaca, idrogeno ed acqua, prodotti che probabilmente potevano
costituire l’atmosfera terrestre primitiva (brodo primordiale).
L’acqua del pallone fu portata all’ebollizione e la miscela di gas fu sottoposta per una
settimana a continue scariche elettriche. Il vapore acqueo trascina con sé i prodotti che si sono
-5-
Fiume Francesco
Fisiologia generale delle piante da orto
formati dalla reazione dei tre gas e sono raccolti in un apposito tubo. Dopo 24 ore dall’inizio
dell’esperimento, circa la metà del carbonio originariamente presente nel gas metano è
trasformato in aminoacidi ed in altre molecole organiche. Nella figura 2 è riportato
schematicamente l’esperimento.
Fig. 2 – Esperienza di Stanley Miller. Apparecchio originale in cui fu compiuto l’esperimento
(A) e corrispondente schema dell’apparecchio (B): fatto il vuoto nell’apparecchiatura,
una miscela di metano, ammoniaca ed idrogeno viene introdotta nel pallone (2);
l’acqua del pallone (1) viene portata all’ebollizione ed il vapore viene spinto nel senso
delle frecce; nel pallone (2), contenente la miscela di gas che simula l’atmosfera
primitiva, viene fatta passare una scarica elettrica per una settimana; il vapore d’acqua
trascina con se i prodotti della reazione che subiscono un raffreddamento (3); i
composti che si formano sono raccolti nel tubo a U (4), che può essere paragonato ad
un oceano primordiale.
In questo modo fu ottenuta la sintesi di parecchi aminoacidi (i mattoni costitutivi delle
principali macromolecole biologiche) e non può essere una coincidenza non significativa il fatto
che proprio alcuni di questi sono quelli più importanti presenti negli organismi viventi.
Numerosi altri studiosi hanno confermato questi esperimenti, con la scoperta che è
possibile ottenere aminoacidi anche a seguito della modificazione della miscela iniziale,
sostituendo ad esempio il metano con l’ossido di carbonio o l’anidride carbonica. E’ stata
-6-
Fiume Francesco
Fisiologia generale delle piante da orto
ottenuta la sintesi degli aminoacidi in condizioni abiologiche artificiali attraverso l’impiego di
diversi tipi di radiazioni come quelle α, β, γ ed ultraviolette o con l’impiego del calore. Dal
grande successo ottenuto con la sintesi artificiale degli aminoacidi con metodi diversi, si deduce
che questi composti organici possono essersi formati in molti ambienti terrestri prima della
nascita della vita.
Se è stato relativamente semplice la dimostrazione che gli aminoacidi possono prodursi
secondo procedimenti abiologici, molto più limitato è risultato l’ottenimento delle proteine per
sintesi prebiologica. E’ stato dimostrato che un poli-α-aminoacido, costituito da residui di
glicina, poteva ottenersi per condensazione di aminoacetonitrile senza formazione intermedia di
aminoacidi ed è stato proposto che questa preproteina possa essere servita come precursore per
la sintesi di polimeri molto più complessi, come le proteine. Tuttavia, nonostante sia stato
possibile introdurre in questa proteina primordiale altre forme aminoacidiche, i prodotti ottenuti
non si sono mai avvicinati alla complessità delle proteine oggi conosciute.
Una convinzione che viene condivisa dalla maggior parte dei ricercatori è che la materia
deve aver preso vita da una serie di tappe, tutte molto probabili. Questa interpretazione risale a
Darwin il quale ipotizzò che la vita abbia avuto inizio quando alcune sostanze attivate dal
calore, dalla luce o da scariche elettriche cominciarono a reagire con altre, generando composti
organici di complessità via via crescente e suggerì una spiegazione del perché oggi non si possa
veder balzare la vita fuori da sostanze inanimate. Un qualsiasi organismo primitivo, scrisse,
verrebbe istantaneamente distrutto o assorbito da quelli più evoluti.
La versione sostenuta da Miller è anch’essa esprimibile secondo i concetti darwiniani. La
vita ebbe inizio quando alcuni composti, oppure una categoria di sostanze fu capace di replicarsi
secondo un processo tale da dare origine, di tanto in tanto, a degli errori ereditabili, in
conseguenza dei quali sono state prodotte nuove generazioni di molecole capaci di copiare se
stesse con maggiore efficienza rispetto a quelle parentali.
Negli anni che seguirono il classico esperimento di Miller, le proteine sembrarono essere i
migliori candidati al ruolo di prime molecole in grado di autoreplicarsi, poiché erano in grado di
riprodursi ed anche di organizzarsi. Un grande sostegno a questa ipotesi prese corpo verso la
fine degli anni cinquanta quando S.W. Fox e K. Harada, scaldando a secco miscugli di
aminoacidi con acido aspartico ed acido glutammico in sufficiente quantità, riuscirono ad
ottenere delle aggregazioni sferiche costituite da polimeri contenenti 18 aminoacidi che si
trovano comunemente nelle proteine. Tali polimeri, detti proteinoidi, hanno un elevato peso
molecolare, sono digeribili dagli enzimi proteolitici e possiedono deboli proprietà catalitiche.
Per tali caratteristiche e per altre ancora, i proteinoidi sono assimilabili qualitativamente alle
attuali proteine. I proteinoidi , tuttavia, rimangono tali nel tempo e non riescono ad evolvere o a
riprodursi. Diversi altri ricercatori, in particolare C.A. Ponnamperuma dell’Università del
Maryland, ripartendo dagli studi di Fox e Harada hanno tentato di ottenere proteine in grado di
replicarsi senza gli interventi di acidi nucleici.
E’ opinione comune che gli acidi nucleici, come il DNA, siano da ritenersi, attualmente, i
più idonei al ruolo di prime molecole capaci di autoreplicazione. La difficoltà è rappresentata
dal fatto che il DNA non può svolgere i propri compiti, compresa la formazione di ulteriore
DNA, senza la presenza di proteine catalitiche quali gli enzimi. In altre parole, le proteine non
possono formarsi senza DNA, ma neppure il DNA può prendere origine senza la presenza di
proteine.
-7-
Fiume Francesco
Fisiologia generale delle piante da orto
A questo dilemma dettero una plausibile risposta gli esperimenti condotti, all’inizio degli
anno ottanta, dai biologi molecolari T.R. Cech dell’Università del Colorado e S. Altman della
Yale University. Era stato ipotizzato che la prima ad autoreplicarsi possa essere stata una
molecola di RNA, ma nessuno aveva dimostrato in che modo essa possa efficacemente dar
luogo a delle copie di se stessa senza la presenza di enzimi. I due ricercatori scoprirono che certi
tipi di RNA potevano comportarsi come enzimi nei confronti di se stessi, tagliandosi in due e
ricucendosi nuovamente.
Una tale scoperta valse ai due studiosi, nel 1989, il premio Nobel e fu subito colta dai
ricercatori interessati al problema dell’origine della vita. Se le molecole di RNA possono agire
come enzima, potrebbero anche replicarsi senza l’aiuto delle proteine. Fu introdotto, da W.
Gilbert dell’Università di Harvard, il termine “mondo a RNA”, per cui i primi organismi
consistevano di semplici molecole di RNA autoreplicantisi. La loro evoluzione indusse la
capacità a sintetizzare le proteine, che favorivano una replicazione più rapida, ed i lipidi che
potevano formare una membrana cellulare. Successivamente questi prebiotici ad RNA
formarono le prime molecole di DNA che avrebbe costituito un più sicuro deposito di
informazione genetica.
In laboratorio sono stati riprodotti con successo i diversi momenti di questa evoluzione.
J.W. Szostak ed altri ricercatori hanno costruito speciali molecole di RNA capaci di agire come
enzimi di restrizione, ossia di tagliare e poi saldare tra loro le diverse molecole, loro stesse
incluse, per molte volte di seguito. Altri ricercatori, in particolare M. Eigen del Max Planck
Institut di Gottinga, dimostrano sperimentalmente come la molecola di RNA, protagonista in
questa vicenda evolutiva dell’origine della vita sul nostro pianeta, sia capace di adattarsi e di
evolversi e sia potenzialmente capace di generare nuovi elementi biologici.
Fig. 3 – Possibile schema di evoluzione prebiotica attraverso un mondo a RNA.
Questo effetto è stato chiamato “evoluzione diretta” ed è schematicamente rappresentato
nella figura 3, dove, attraverso un mondo a RNA, lo stesso acido ribonucleico si forma dal
ribosio e da altri composti organici (a), si evolve apprendendo al capacità della replicazione (b),
inizia a sintetizzare proteine dotate di capacità catalitiche (c), le quali aiutano il nucleotide a
-8-
Fiume Francesco
Fisiologia generale delle piante da orto
replicarsi ed a sintetizzare nuove proteine caratterizzate da una maggiore efficienza ed a
costruire versioni a doppio filamento di se stesso che poi evolveranno in DNA (d), il quale
assumerà una posizione preminente nell’utilizzare le molecole di RNA per costruire ulteriori
proteine che, a loro volta, aiutano lo stesso DNA a produrre copie di se stesso ed a trasferire
l’informazione genetica allo stesso RNA (e).
Questo antico mondo prebiotico, formato da molecole di RNA poteva aver rappresentato
un ponte tra la chimica dei composti semplici ed i prototipi di quelle cellule complesse, la cui
sopravvivenza nel tempo è assicurata dal DNA che costituisce il patrimonio ereditario degli
attuali organismi. Queste cellule, secondo alcune testimonianze fossili, sarebbero comparse
durante il primo miliardo di anni che seguì la formazione del nostro pianeta, databile intorno a
4,5 miliardi di anni.
L’attento esame della teoria del “mondo a RNA” pone il quesito principale di come possa
essersi formato il primo RNA.
La molecola di RNA ed i suoi componenti sono difficili da sintetizzare in laboratorio,
anche nelle migliori condizioni sperimentali e tali difficoltà saranno state ampiamente
amplificate in un mondo primordiale, prima dell’origine della vita.
In particolare, la sintesi dello zucchero chiave dell’acido ribonucleico, il ribosio, produce
tutta una serie di altri zuccheri che potrebbero inibire la replicazione dello stesso RNA. Inoltre,
non è chiaro perché il fosforo, un elemento relativamente raro in natura, sia così fortemente
rappresentato nella molecola di RNA e DNA. La sintesi in laboratorio di RNA può ottenersi, ma
la sua replicazione può avvenire soltanto in determinate condizioni che non è detto che si siano
verificate all’epoca dell’origine della vita.
Alcuni ricercatori e tra questi L.E. Orgel ritengono che qualche molecola più semplice e
forse del tutto diversa possa aver spianato la strada per la sintesi di RNA. L’identificazione di
questo composto non è facile ed un gruppo di ricercatori, coordinato da J. Rebek, lavorando
intorno a questo problema ha condotto un’ampia sperimentazione che ha portato, intorno alla
metà degli anni novanta, alla sintesi di nuove molecole organiche in grado di autoreplicarsi.
Molecole autoreplicanti
Le molecole, naturali o sintetiche, riescono a replicarsi quando le loro forme e le loro
proprietà chimiche sono dotate di complementarietà.
Una molecola, in virtù di come occupa lo spazio e di come i suoi atomi o gruppi atomici
sono distribuiti al suo interno, può posizionarsi nei recessi e negli angoli reconditi di un’altra
molecola. L’adattamento ottimale tra due molecole complementari dipende così non soltanto
dalla struttura spaziale delle molecole, ma anche dai differenti tipi di legami chimici che le
tengono unite in gruppi. Questi gruppi o complessi si formano e si dissociano rapidamente, nello
spazio di frazioni infinitesime di secondo, in tempi molto brevi ma sufficientemente lunghi da
permettere lo svolgimento di reazioni chimiche.
Le forze che tengono uniti i complessi, spesse volte più deboli dei legami covalenti che si
stabiliscono tra gli atomi nelle molecole, sono costituiti dai legami idrogeno, dalle forze di van
der Waals e dall’impilamento aromatico.
Il legame a idrogeno o legame polare si forma quando un atomo di idrogeno dotato di una
carica parziale positiva è attratto da una atomo di ossigeno che ha una carica parziale negativa.
-9-
Fiume Francesco
Fisiologia generale delle piante da orto
Le forze di van der Waals sono quelle che si stabiliscono tra due molecole, quando gli
elettroni di una riescono a spostare quelli dell’altra, creando uno squilibrio di carica e stabilendo
tra i due complessi delle forze di attrazione.
Il terzo tipo di attrazione, quello dell’impilamento aromatico, si stabilisce tra le superfici
piane di molecole cicliche aromatiche le quali si avvicinano le une alle altre, ponendo a contatto
diretto le loro facce piatte e venendo a stabilire una configurazione più stabile. Si viene a
stabilire una repulsione per il solvente in cui tali molecole si trovano, per cui lo stesso solvente
non può interporsi tra le superfici molecolari combacianti che risultano così protette, poiché
solventi in grado di danneggiarle, acidi, basi, ossidanti in soluzione, non sono in grado di
raggiungerle. Allora legami covalenti forti hanno il tempo di congiungere le parti complementari
e, talvolta, soltanto due su tre molecole di un complesso formano un legame, mentre la terza
serve semplicemente a facilitare il processo. Il ricorso a forme concave e convesse può essere
utile per rappresentare schematicamente il tipo di accoppiamento descritto. Una superficie
molecolare concava può riconoscere e circondare il proprio complemento convesso e può
fungere da stampo per assemblare la molecola convessa a partire dalle sue parti componenti.
Reciprocamente la molecola convessa può costituire uno stampo per riunire e fondere insieme le
parti componenti della molecola concava. Si ottiene, in pratica, che ciascuna molecola forma
l’altra e questi due tipi di replicazione costituiscono un bi-ciclo.
Nella figura 4 è indicata la rappresentazione schematica del riconoscimento molecolare di
due frammenti, con proprietà geometriche e chimiche complementari, fra i quali il solvente che
s’interpone viene eliminato, con conseguente stabilizzazione del complesso neoformato. Si
realizza un accoppiamento di due molecole complementari con attrazioni elettrostatiche dei
segni opposti.
Fig. 4 – Semplice accoppiamento di due molecole complementari con attrazioni elettrostatiche
indicate con i segni + e -. Questo tipo di accoppiamento riproduce uno schema
abbastanza diffuso di replicazione che è quello preferito dal DNA.
- 10 -
Fiume Francesco
Fisiologia generale delle piante da orto
Nella figura 5 è schematicamente rappresentato un bi-ciclo di replicazione coinvolgente
due molecole che hanno forme complementari rappresentate da due elementi in cui il primo si
inserisce nel secondo.
Fig. 5 – Bi-ciclo di replicazione in cui l’elemento A raccoglie le due parti di B attorno a sé per
formare un complesso; le parti reagiscono tra loro formando un elemento B intero per
poi dissociarsi velocemente (ciclo a sinistra). L’elemento B riunisce i frammenti
dell’elemento A in modo che le due molecole complementari catalizzano ciascuna la
formazione dell’altro (ciclo a destra).
Un modello alternativo di replicazione è rappresentato da due molecole complementari
all’interno di un complesso che si uniscono in qualche punto che non si trova sulla superficie di
riconoscimento, con la formazione di un’unica molecola di cui un’estremità è complementare
dell’altra e l’insieme è complementare di se stesso.
Fig. 6 – Se due molecole complementari A e B, anziché inserirsi l’una nell’altra come avviene
in un bi-ciclo, si uniscono in corrispondenza di un altro sito di reazione formando una
molecola autocomplementare, ha luogo un’autoreplicazione. La molecola assembla
copie delle forme originarie e le trattiene in modo che anch’esse possano reagire per
dare origine ad una copia di sé stessa.
- 11 -
Fiume Francesco
Fisiologia generale delle piante da orto
Le superfici di riconoscimento alle estremità di questa nuova molecola
autocomplementare sono ancora accessibili ad altre molecole e possono catturare, ciascuna, un
frammento identico a quello che si trova all’altra estremità. Una volta catturate, queste due
nuove componenti non possono più muoversi liberamente e si spostano insieme nello spazio e le
probabilità che si uniscano l’una all’altra sono molto potenziate. In tal modo, l’entità
autocomplementare produce una copia e, identicamente, molte copie di sé. Non sono necessari
enzimi poiché è la stessa molecola che catalizza la propria formazione (figura 6).
Fig. 7 – Una molecola autocomplementare (ARNI) riunisce i componenti di cui ha bisogno per
replicarsi: una molecola di adeninribosio ed una molecola di naftalenimmide. Gli aloni
colorati indicano gli atomi coinvolti nei legami a idrogeno.
- 12 -
Fiume Francesco
Fisiologia generale delle piante da orto
E’ stato proprio questo il metodo che J. Rebek, nel 1994 ha utilizzato in laboratorio per
produrre molecole che fossero in grado di reagire l’una con l’altra in modo da richiamare alla
mente il mondo vivente. Quando sono associate insieme in un complesso, l’adenina e l’immide
si uniscono mediante legame covalente e formano una molecola autocomplementare.
L’inserimento tra l’adenina e l’immide di una molecola di naftalene (più voluminosa e più
rigida) al posto della catena a legami singoli e di un gruppo di ribosio ciclico, quale unità di
collegamento meno flessibile, ha lo scopo di impedire che la lunga catena di atomi di carbonio,
quale risultante struttura autocomplementare, si ripiegasse su se stessa senza associarsi ad altre
molecole e senza replicarsi. La nuova molecola ottenuta è stata l’adeninribosonaftalenimmide
(ARNI) che offrì uno dei primi esempi di autoreplicazione (figura 7).
Tuttavia, l’ARNI non andava incontro ad un accrescimento secondo una curva sigmoide,
tipico degli acidi nucleici. Per ottenere ciò si è dovuto inserire una superficie per l’impilamento
leggermente più lunga, cioè un bifenile al posto del naftalene per avere così una nuova molecola
sintetica che si autoreplica autenticamente, cioè l’adeninribosobifenlimmide (ARBI). L’ARBI,
però, pur essendo in grado di replicarsi, è una molecola capace di fare copie soltanto di se stessa,
mentre un prodotto ereditabile tipico degli organismi viventi, affinché possa dar luogo ad un
processo evolutivo, deve poter sintetizzare di tanto in tanto altre molecole in grado di svolgere
una migliore attività duplicativa e dar luogo a variabilità.
E’ stato necessario ottenere, per impostare uno schema che potesse dare un’idea di come
la vita ha avuto origine sul nostro pianeta, una molecola che catalizzasse non soltanto la propria
formazione, ma anche quella di una molecola di forma simile, tuttavia dotata di una maggiore
capacità di replicazione.
Rebek e collaboratori hanno progettato molecole in grado di fare “errori”, di subire, in
altre parole, delle mutazioni, cioè delle modificazioni strutturali e biochimiche sotto l’azione di
cause ambientali, come, per esempio, le radiazioni ultraviolette. Si trattava di simulare in
laboratorio quello che probabilmente accadde sul nostro pianeta qualche miliardo di anni fa,
quando si verificarono le condizioni che determinarono la nascita della vita.
In chimica organica, si verifica un “errore” quando i reagenti non possiedono selettività.
Per produrre un’ipotesi più o meno valida sulla nascita della vita bisognava disporre di una
molecola che catalizzasse non soltanto la propria formazione, ma anche quella di una molecola
di forma simile. Inoltre, almeno una di queste due molecole doveva essere in grado di
trasformarsi in un’altra dotata di maggiore capacità di replicazione.
Gli studi condotti sul DNA da Watson e Crick dimostrarono l’esistenza di due siti
dell’adenina dove si formano i legami a idrogeno: un sito lungo il cosiddetto spigolo di Watson
e Crick, che è interessato nella replicazione del DNA, ed un sito presente lungo lo spigolo di
Hoogsteen, una parte del DNA che rimane normalmente esposta, anche se a volte è congiunta in
eliche triple. Orbene, le immidi di Rebek possono attaccarsi in corrispondenza dell’uno o
dell’altro spigolo e se uno degli idrogeni del gruppo amminico (–NH2) dell’adenina, per
l’appunto interessato nel legame a idrogeno, viene sostituito da un gruppo di maggiori
dimensioni, quest’ultimo blocca l’accesso allo spigolo di Watson e Crick, mentre lo spigolo di
Hoogsteen rimane ampiamente accessibile. Infatti, quando un gruppo metilico (–CH3) si unisce
all’adenina, si rileva che oltre 85% dei recettori delle immidi si lega lungo lo spigolo di
Hoogsteen.
Utilizzando il cambiamento della velocità di replicazione, conseguenza del blocco dello
spigolo di Watson e Crick, sono state ottenute due diverse molecole di adenina: una con un
- 13 -
Fiume Francesco
Fisiologia generale delle piante da orto
gruppo benzilossicarbonilico (Z), bloccante nella sintesi delle proteine, ed un’altra con lo stesso
gruppo al quale era stato aggiunto un radicale –NO2 (Z–NO2). Ciò allo scopo di assemblare
sullo stampo una molecola alterata di adenina ed un’immide, con gruppi bloccanti penzolanti in
siti lontani dal punto dove si forma il legame covalente, così che la sintesi potesse svolgersi
indipendentemente dall’identità dei gruppi, poiché Z ad un’estremità non sapeva se il gruppo
bloccante all’altra estremità poteva essere Z oppure Z-NO2. Prendendo ancora in considerazione
la molecola autoreplicante ARBI, questa è stata modificata in ZARBI (con molecole di adenina
contenenti il gruppo Z) e ZNARBI (con molecole di adenina provviste del gruppo Z-NO2), due
molecole in grado di autoreplicarsi lentamente e di compiere “errori”. Infatti, una catalizza la
propria formazione e funge da stampo per l’assemblaggio dell’altra, mentre quest’ultima
catalizza la propria formazione e quella della prima.
Va anche detto che l’addizione di molti gruppi può portare alla formazione di molecole
autoreplicanti in grado di compiere “errori”, tuttavia –NO2 si comporta in modo particolare,
poiché è facile rimuoverlo con irraggiamento mediante particolari lunghezze d’onda
nell’ultravioletto.
Una volta staccato il gruppo Z-NO2, lo spigolo di Watson e Crick diventa accessibile, la
nuova molecola diventa più leggera ed è possibile trovare una corrispondenza oltre che lungo lo
stesso spigolo anche verso quello di Hoogsteen con un raddoppiamento dell’efficienza di
autoreplicazione.
Le ricerche di Rebek e collaboratori hanno così portato alla realizzazione di una versione
chimica della mutazione, con un cambiamento strutturale permanente, ereditabile, che influisce
sulla capacità di sopravvivenza di un organismo o del suo analogo, come in questo caso, la
molecola autoreplicante.
Inoltre, come forse avvenne nelle condizioni ambientali all’origine della vita, le
modificazioni nella struttura di una molecola autoreplicante possono essere causati da variazioni
termiche, acidità, salinità, radiazioni e molti altri fattori.
Facendo competere i derivati adeninici contenenti i gruppi Z e Z-NO2 per una quantità
limitata del recettore bifenilico complementare, una volta avvenuto il consumo completo del
recettore, il recipiente di reazione è stato irradiato con luce ultravioletta della lunghezza d’onda
di 350 nm.
Dopo alcune ore di irradiazione i gruppi bloccanti Z-NO2 sono stati tutti rimossi, sia dalle
molecole autoreplicanti ZNARBI, sia dai loro progenitori adeninici. In altre parole, le molecole
ZNARBI sono state tutte trasformate in molecole ARBI e le molecole di adenina contenenti i
gruppi Z-NO2 sono diventate semplicemente adenina.
Era stata ottenuta una vera mutazione, sollecitata da un cambiamento dell’ambiente. Le
molecole ZARBI e la Z-adenina sono rimaste inalterate, ma a seguito dell’aggiunta di altro
recettore bifenilico, la molecola ARBI, prodotto dell’irradiazione, diventa loro concorrente e
prende il completo e rapido sopravvento nell’utilizzazione delle risorse del sistema, sia perché di
forma più affusolata, sia per il vantaggio di replicarsi lungo lo spigolo di Watson e Crick e
quello di Hoogsteen, ambedue privi di gruppi bloccanti.
Nella figura 8 viene riportato lo schema di ottenimento di molecole autoreplicanti e
mutanti, quando una molecola di adenina, provvista di un qualunque gruppo supplementare (ad
esempio un gruppo Z o Z-NO2) si unisce con una molecola di bifenilimmide, formando una
molecola autocomplementare.
- 14 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 8 – Ottenimento di molecole mutanti autocomplementari a partire da adenina, provvista di un
gruppo R, e da bifenilimmide. R può essere un gruppo Z o Z-NO2, il che dà luogo ad una
molecola ZARBI o NZARBI, rispettivamente. Quest’ultima può fissare una molecola di
adenina soltanto lungo lo spigolo di Hoogsteen e non con lo spigolo di Watson e Crick,
poiché questo è bloccato dal gruppo R, e fonderla con una molecola di bifenilimmide. La
molecola NZARBI realizza così la catalisi della propria formazione, oltre a quella della
molecola competitrice.
- 15 -
Fiume Francesco
Fisiologia generale delle piante da orto
Diventa ora possibile proporre una semplice spiegazione evoluzionistica. Supponendo che
la molecola ZARBI sia la molecola origine, la sua replicazione comporta necessariamente la
presenza di una Z-adenina e del recettore bifenilico. Addizionando acido nitrico, alcune
molecole di Z-adenina acquistano il gruppo Z-NO2 e vanno a costituire molecole ZNARBI le
quali si autoreplicano con maggiore efficienza del predecessore ZARBI. Con l’irraggiamento di
luce ultravioletta si verifica un’altra modificazione per la quale ZNARBI si trasforma nella
molecola ARBI più semplice ed efficiente, poiché dimostra la migliore autoreplicazione.
La mutazione è il fenomeno più efficiente in quasi tutti i processi evolutivi, ma anche la
ricombinazione assume importanza fondamentale nell’evoluzione. Due cromosomi possono
dividersi, scambiarsi filamenti di DNA e ricongiungersi, combinando in tal modo anche i loro
caratteri.
La mutazione induce piccoli singoli cambiamenti, mentre la ricombinazione permette
l’ottenimento di ibridi che sono molto diversi dai loro genitori. Orbene, gli studi di Rebek e
collaboratori, allo scopo di dimostrare la ricombinazione a livello molecolare, hanno condotto
alla produzione di un insieme di molecole autoreplicanti completamente nuovo. Due molecole
complementari vengono unite da un legame covalente per dare origine ad una struttura
autocomplementare capace di agevolare la propria sintesi. Specificamente, sono state ottenute
molecole autoreplicanti basate sull’adenina e sulla timina le quali, introdotte nello stesso
reattore, sono state in grado di rimescolare le proprie componenti in nuove combinazioni ed
alcuni dei prodotti della ricombinazione, la diamminotriazinxantentimina (DIXT) e
l’adeninribosotimina (ART) hanno dimostrato di essere molecole dotate di autoreplicazione, in
particolare la seconda è stata la molecola autoreplicante più prolifica mai incontrata, mentre
un’altra, la diamminotriazinxantenbifenilimmide (DIXBI) non riusciva affatto a replicarsi,
mostrando caratteristiche, per così dire, di sterilità.
L’efficienza autoreplicante di ART si può spiegare col fatto che questa molecola
assomiglia molto ad un segmento di DNA che, probabilmente, è la molecola che si autoreplica
nel migliore dei modi fra tutte quelle esistenti. Il ribosio di cui è provvista dà luogo ad una
configurazione molto utile allo scopo, poiché rende le superfici di riconoscimento parallele le
une alle altre. L’elevata affinità dell’adenina per la sua complementare timina permette, inoltre,
la formazione di un complesso che si assembla senza difficoltà.
L’inefficienza nell’autoreplicazione di DIXBI è legata alla sua conformazione molecolare
globale, poiché questa è costituita da due molecole a forma di U, unite da un distanziatore
bifenilico rigido che induce una struttura complessiva a forma di C o a S.
Nella prima forma le superfici di riconoscimento sono affacciate all’interno, dove non c’è
spazio sufficiente per la formazione di un complesso autoreplicante.
Nella seconda forma le superfici di riconoscimento sono molto distanziate, per cui, quando
si forma un complesso, le parti reattive sono troppo distanti l’una dall’altra e non riescono ad
unirsi tramite un legame covalente.
Ciò spiega perché la DIXBI, pur essendo una molecola autocomplementare non è capace
di autoreplicazione.
Un insieme relativamente piccolo di molecole può, pertanto, originare un vero albero
genealogico di molecole dotate della peculiarità dell’autoreplicazione. Nella figura 9 viene
rappresentato lo schema di un albero genealogico di molecole autoreplicanti, per cui una
soluzione con frammenti molecolari complementari può generare alcune molecole che si
autoreplicano diverse tra loro.
- 16 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 9 – Rappresentazione di un albero genealogico di molecole capaci di autoreplicazione.
L’adenin ribosio e la bifenilimmide producono l’ARBI e se la soluzione, una sorta di
brodo primordiale, contiene anche diamminotriazinxantene e timina avviene la sintesi
anche di ART, DIXT e DIXBI. La prima molecola è la più prolifica delle quattro,
mentre l’ultima è assolutamente sterile.
- 17 -
Fiume Francesco
Fisiologia generale delle piante da orto
Alcune di queste molecole si autoreplicano con buona efficienza, mentre un ramo
dell’albero si estingue per assenza della capacità autoreplicante.
Molto interessante sarebbe la reale possibilità che le molecole sterili, frantumate in parti,
potessero dar luogo a raggruppamenti chimici che le molecole con elevata capacità di
autoreplicazione sarebbero in grado di utilizzare a loro vantaggio.
Dalle molecole autoreplicanti ad un modello di cellula primordiale
Un altro evento importante per la comparsa della vita è la costruzione di una membrana
capace di operare una separazione spaziale tra gli aggregati di molecole e l’ambiente. In altri
termini, le conquiste evolutive, raggiunte da un sistema attraverso apposite sintesi biochimiche,
sono vanificate se vi è la libera circolazione delle macromolecole e dei loro prodotti che
altrimenti verrebbero condivisi dai loro competitori. La membrana impedisce, inoltre, la
penetrazione dall’esterno di molecole in grado di disorganizzare il programma abbozzato,
operando da barriera grazie alla sua continuità spaziale ed alla sua particolare struttura fisicochimica. La natura lipoproteica, infatti, rende la membrana altamente isolante ed impermeabile
alle molecole idrofile ed a quelle di grosse dimensioni, anche se tale funzione di
impermeabilizzazione non è assoluta, nel senso che la stessa membrana,con una certa selettività,
può far passare o trasportare molecole appartenenti alle più diverse categorie, presupposto
essenziale al mantenimento della vita della cellula.
Gli eventi che hanno condotto alla formazione della membrana sono sconosciuti e tra le
tante teorie ce n’è una che chiama in causa complessi che sono detti coacervati (figura 10).
I coacervati, secondo A.I. Oparin possono ottenersi in laboratorio dissolvendo una
proteina in acqua. Se si aggiunge un acido si osserva l’intorbidamento del liquido dovuto alla
comparsa di migliaia di piccole goccioline, per l’appunto i coacervati, visibili al microscopio e
del diametro compreso tra 0,5 e 2.000 nm, che si formano per l’esistenza sulle macromolecole di
cariche elettriche che attirano l’acqua.
I coacervati sono delimitati all’esterno da un evidente strato con carattere di membrana,
attraverso il quale possono verificarsi scambi in maniera selettiva.
Fig. 10 – Gocce di coacervato, ottenuto mescolando gelatina e gomma arabica (a sinistra) ed
addizionando RNA ad un istone.
- 18 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 11 – Lo schema di formazione delle microsfere distinto in quattro tappe. La prima comporta
la presenza di gas costituenti l’atmosfera primordiale (anidride carbonica o metano,
ammoniaca, acqua ed idrogeno); questi subiscono una condensazione e danno luogo
alla seconda tappa con formazione di aminoacidi; un successivo addensamento degli
amminoacidi genera la terza tappa con formazione dei proteinoidi; infine la quarta
tappa che produce microsfere dotate di membrane limitanti e di gemme. Nelle due
microfotografie è possibile osservare delle microsfere (A), ottenute da S.W. Fox, nelle
quali è ben visibile la membrana, le quali stanno producendo un processo formativo
delle gemme (B).
- 19 -
Fiume Francesco
Fisiologia generale delle piante da orto
Questi modelli di cellule sono molto interessanti, poiché è possibile che simili aggregati
selettivi di macromolecole si siano formati negli oceani primordiali. S.W. Fox e S. Yuyama
sono riusciti ad ottenere coacervati a partire da proteinoidi, preparati per via termica per
immersione in soluzioni saline calde.
Fenomeni analoghi, svoltisi in un cosiddetto brodo primordiale, avrebbero permesso la
formazione di aggregati colloidali complessi e coacervati, le cui dimensioni avrebbero superato
da 200 a 1.000 volte quelle delle singole macromolecole. Queste ultime si sarebbero concentrate
in punti localizzati nella massa fluida, fino alla comparsa dell’individualità. Ciascun coacervato
o microsfera o micella, gradualmente, avrebbe avuto a disposizione un proprio ambiente interno
in cui sarebbero avvenute le principali reazioni chimiche attraverso l’assorbimento, regolato da
una membrana rudimentale, di materiale organico dalla soluzione acquosa esterna. Ciascuna
goccia, da questo momento, avrebbe seguito un differente cammino: avrebbe potuto evolversi o
estinguersi (figura 11).
La successiva evoluzione delle microsfere avrebbe portato ad intravedere i primi passi di
un rudimentale metabolismo. Esse si sarebbero orientate verso una graduale organizzazione in
semplici sistemi aperti, capaci di scambiare energia e materia mediante processi di diffusione ed
osmosi attraverso le membrana, con passaggio preferenziale di alcune molecole come l’acqua,
gli zuccheri semplici, gli aminoacidi, prefigurando così i primi processi nutritivi, probabilmente
di tipo eterotrofo.
Uno degli attributi chiave della vita è una delimitazione rappresentata dalla parete di un
contenitore e, specificamente, di una cellula che separi l’interno dall’esterno ed impedisca alle
molecole utili di essere portate via, mentre le molecole che non interessano vengono tenute sotto
controllo.
E’ noto che i virus utilizzano come contenitore un involucro proteico di molte copie
identiche di una sola unità proteica.
Le unità sono autocomplementari, ma le loro superfici di riconoscimento sono orientate in
modo che esse si assemblano in un involucro chiuso.
Il capside virale è costituito da molte copie identiche di una proteina, poiché il genoma
virale non possiede una quantità di informazioni adeguata per il coinvolgimento di diverse
molecole.
Rebek e collaboratori hanno pensato ad un progetto molto semplice basato sulla struttura
di una palla da tennis.
Tagliata lungo la cucitura, essa dà luogo a due metà identiche, le cui estremità convesse
sono complementari come forma alle parti centrali concave.
E’ stata sintetizzata, in laboratorio, una struttura che imita le forme dei pezzi della palla da
tennis e possiede una complementarietà chimica, con unità che si adattano bene tra loro con
legami a idrogeno lungo la cucitura (figura 12).
Una molecola sintetica è stata creata per adattarsi bene all’interno, tuttavia, altri ostacoli si
interpongono affinché possa dimostrarsi sino in fondo che un simile progetto possa aver
realmente dato luogo all’origine di una struttura vivente. In particolare, è molto difficile
dimostrare come un simile pseudorganismo possa trarre energia e da dove, cioè dalla luce o da
altre molecole, ed ancora ci si chiede come possono essere reintegrate le parti componenti delle
molecole autoreplicanti ed i loro contenitori.
- 20 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 12 – Ipotesi della palla da tennis sull’origine della membrana cellulare, vale a dire di un
contenitore autoreplicante di molecole autoreplicanti in grado di separare l’interno
dall’esterno. Sulla destra viene indicata la rappresentazione di una molecola che,
assemblandosi con la propria gemella, dà origine ad una sfera cava in grado di
contenere molecole autoreplicanti.
Questi esperimenti starebbero a significare che le conoscenze dei concetti intimi che
stanno alla base della vita e della creazione siano prossimi ad essere svelati, mentre le cose non
stanno proprio in questi termini e si può sostenere logicamente che la vita è così complessa che
potrebbe essere stata creata da un inventore intelligente e la spiegazione possa essere ancora
correlata all’esistenza di un Creatore.
Anche se si riuscisse a creare in laboratorio qualcosa dotato di proprietà simili alla vita,
rimarrebbe sempre in sospeso la domanda se le cose andarono effettivamente in quel modo. La
risposta è molto difficile dal momento che la tettonica delle zolle, il vulcanismo e l’erosione
hanno cancellato la maggior parte delle tracce risalenti al primo miliardo di anni del nostro
pianeta. Basti pensare che per stimare l’età della Terra bisogna rivolgersi alle meteoriti che si
presume siano relitti dell’era in cui il sistema solare si condensò da una nube di gas e polvere e
che il tasso di decadimento radioattivo osservato nelle stesse meteoriti indica che esse, e così la
Terra, hanno approssimativamente un’età di 4,5 miliardi di anni.
La vita è il prodotto più straordinario scaturito dall’interazione tra sistemi complessi. Lo
sviluppo di un organismo vivente è il frutto di una serie articolata di rapporti reciproci che
coinvolgono un gran numero di componenti diverse. Queste componenti, o sottosistemi, sono a
loro volta costituiti da elementi molecolari più piccoli, ciascuno dei quali, indipendentemente
dagli altri, mostra un comportamento dinamico tipico, come, per esempio, la capacità di
catalizzare reazioni chimiche. Quando questi elementi sono combinati insieme in unità
funzionali più grandi ne scaturiscono nuove ed imprevedibili proprietà come la capacità di
accrescersi, di nutrirsi, di moltiplicarsi e di reagire agli stimoli. L’unità funzionale dotata di
queste caratteristiche e che rappresenta la chiave per capire come funziona la vita è la cellula ed
il fenomeno per il quale diverse componenti si uniscono per formare strutture stabili più grandi,
dotate di proprietà inedite rispetto a quelle delle singole componenti, è noto come
autoassemblaggio. La cellula è l’unità fondamentale della vita, poiché tutti gli organismi viventi,
- 21 -
Fiume Francesco
Fisiologia generale delle piante da orto
siano essi batteri, animali o vegetali, sono costituiti da cellule e la loro conoscenza rappresenta
la chiave per capire la struttura e la funzione delle piante e degli animali.
La cellula
La cellula fu osservata fin dal 1674 quando l’olandese A. van Leeuwenhoek gettò lo
sguardo in uno dei microscopi a lente singola che egli stesso aveva costruito, scoprendo un
universo del tutto nuovo ed affascinante. Lo studioso olandese inconsapevolmente inaugurò
l’era della microscopia, in un’epoca nella quale l’ottica non si era ancora sviluppata in una vera
e propria scienza. Egli costruì da solo oltre 500 microscopi e per mezzo di questi strumenti
ancora rudimentali riuscì ad osservare molte strutture cellulari, scoprì l’esistenza dei globuli
rossi del sangue e degli spermatozoi e descrisse anche batteri, protozoi, cellule vegetali e funghi.
I primi organismi cellulari che apparvero sul nostro pianeta, all’incirca 3,7 miliardi di anni
fa, erano di dimensioni microscopiche e formati da una sola cellula. Essi erano molto simili ad
alcuni batteri attuali. Le cellule di questo tipo sono classificate come procarioti perché prive di
un nucleo ben delimitato, contenente il meccanismo genetico responsabile dell’ereditarietà dei
caratteri. I procarioti ebbero un grande successo e, grazie alla loro straordinaria capacità di
evoluzione e di adattamento, diedero origine ad una grande varietà di specie ed invasero ogni
forma di habitat che il mondo aveva da offrire.
Le forme viventi del nostro pianeta sarebbero ancora costituite totalmente da procarioti se
non fosse intervenuto un processo straordinario che dette origine ad un tipo di cellula molto
diverso, la cellula eucariote, dal greco eu che significa bene e karyon che vuol dire nucleo,
provvista di un nucleo vero e proprio. Le conseguenze di questo avvenimento furono
straordinarie ed oggi tutti gli organismi pluricellulari sono formati da cellule eucarioti, molto più
complesse e con più elevate possibilità evolutive dei procarioti.
In un mondo senza cellule eucarioti non sarebbe stata possibile l’esistenza di un’intera
variegata schiera di organismi vegetali ed animali e non sarebbero esistiti neppure gli esseri
umani a godere di questa diversità ed a penetrarne i segreti.
La cellula procariote ed eucariote
L’evoluzione della cellula fino al tipo eucariote si realizzò, con ogni probabilità, a partire
da antenati procarioti. E’ difficile dire come questo avvenne poiché nessun intermedio di questa
transizione è sopravvissuto oppure ha lasciato fossili in grado di fornire indizi diretti. L’unica
cosa che è possibile osservare è il prodotto finale di questo processo evolutivo, cioè qualcosa di
straordinariamente diverso da qualsiasi cellula procariote.
Per valutare un simile percorso di evoluzione bisogna comprendere le diversità tra i due
tipi fondamentali di cellule.
Le cellule procarioti sono molto più piccole di quelle eucarioti, poiché hanno un volume
10.000 volte minore. L’aspetto più evidente che distingue la cellula procariote da quella
eucariote è la presenza, in quest’ultima, del nucleo nel quale gran parte del DNA è contenuto nei
cromosomi dalla struttura molto complessa. Nei procarioti tutto il materiale genetico è contenuto
nell’unico cromosoma, costituito semplicemente da un filamento circolare di DNA, immerso
nella massa citoplasmatica.
Negli eucarioti sono presenti, nella regione che circonda il nucleo e che costituisce il
citoplasma, una serie elaborata di compartimenti, con funzioni molto diversificate, delimitati da
membrane. Nello stesso citoplasma vi sono gli elementi scheletrici, che conferiscono alla cellula
- 22 -
Fiume Francesco
Fisiologia generale delle piante da orto
eucariote un sostegno strutturale, mentre alcuni piccoli motori molecolari permettono il
rimescolamento del proprio contenuto ed anche movimenti di spostamento.
Le membrane che inducono la compartimentazione delle cellule eucarioti, osservate al
microscopio elettronico, appaiono molto simili nei vari organismi animali e vegetali. Esse sono
costituite da tre strati, cioè da due strati scuri, ciascuno dello spessore di circa 250 nm, separati
da uno strato chiaro dello spessore di circa 35 nm.
Gli eucarioti si distinguono ulteriormente dai procarioti per la presenza di migliaia di
strutture specializzate subcellulari, gli organelli, aventi dimensioni all’incirca di una cellula
procariote.
Anche gli eucarioti unicellulari, come per esempio i lieviti e le amebe, sono provvisti di
numerosi organelli e di complicate strutture interne che fanno di essi, a differenza dei procarioti,
organismi dotati di un’enorme complessità.
Una caratteristica molto importante della cellula eucariote è la proprietà della
differenziazione per la quale, nello stesso organismo, cellule geneticamente uguali possono
assumere forme e funzioni molto diverse e dotate di alta specificità. Possono, in tal modo,
formarsi i diversi tessuti ed organi con un’esaltazione delle proprietà tipiche delle diverse specie
animali e vegetali.
Queste sono soltanto alcune delle differenze tra le cellule procariote e quelle eucariote,
poiché esse sono ancora più numerose e concettuali.
Le cellule eucarioti vegetali presentano delle loro particolari caratteristiche ed una loro
trattazione, come ripetizione ed anteprima, viene ora effettuata allo scopo di conoscere le loro
principali strutture ed alcune delle loro funzioni meglio conosciute.
LA CELLULA VEGETALE
La cellula vegetale è costituita da una parete più o meno rigida e da un protoplasto.
Quest’ultimo è formato dal citoplasma e dal nucleo.
Il citoplasma comprende un certo numero di entità definite, dette organelli, che
comprendono i ribosomi, i microtubuli, i plastidi ed i mitocondri, oltre a sistemi di membrana
quali il reticolo endoplasmatico ed i corpi del Golgi.
Nella figura 13 è possibile osservare lo schema ed una microfotografia di una cellula
vegetale.
Guardando la microfoto è evidente la forma regolare e rigida della cellula vegetale, dovuta
alla presenza di una struttura esterna alla membrana, la parete cellulare.
Sempre nella stessa foto sono evidenti i plasmodesmi che collegano tra loro i protoplasti
di cellule contigue. Dallo schema della figura si può osservare come, in una cellula vegetale
adulta, il citoplasma, contenente il nucleo ed i vari organelli, sia una sottile lamina tutta attorno
al vacuolo.
La cellula vegetale possiede molte strutture che sono proprie peculiarità come i
cloroplasti, il vacuolo, la parete ed i microcorpi. In particolare, la presenza del cloroplasto,
propria del sistema plasmidiale della cellula vegetale, rende possibili quei processi di crescita
delle piante e quel tipo di nutrizione e di metabolismo cellulare (assimilazione del carbonio e
dell’azoto ed emissione di ossigeno nell’atmosfera) che permettono il mantenimento della vita
sul nostro pianeta.
- 23 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 13 – Microfotografia di una tipica cellula vegetale adulta, con schema tridimensionale e
bidimensionale nei quali cercano di identificarsi alcuni degli organelli cellulari.
- 24 -
Fiume Francesco
Fisiologia generale delle piante da orto
Nello studio della fisiologia della cellula vegetale sono ora esaminate le principali strutture
cellulari rappresentate dal contenitore, cioè la parete cellulare, dalle membrane, dal citoplasma,
dai mitocondri, dai plastidi, dai microcorpi, dal vacuolo.
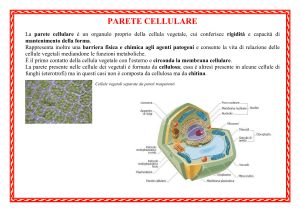
PARETE CELLULARE
La parete cellulare è una struttura tipica delle sole cellule vegetali delle quali ne avvolge il
protoplasto. Di quest’ultimo ne determina la forma e lo protegge dagli eventuali traumi e
conseguenti danni.
Per comprendere una delle più importanti funzioni della parete basta ricordare che le
cellule vegetali potrebbero subire, a seguito dell’assorbimento e della perdita d’acqua, forti
variazioni di volume.
La parete cellulare costituisce un involucro sufficientemente resistente e tale da difendere
la stessa cellula dai danni che altrimenti deriverebbero dalla variazione della pressione di
turgore. In particolare, con la plasmolisi, che si verifica quando la cellula vegetale viene
immersa in una soluzione sufficientemente concentrata di saccarosio o di qualsiasi altro soluto
(ad esempio cloruro di sodio), si ha fuoriuscita di acqua dal protoplasto, con conseguente
contrazione dello stesso. Ma la cellula continua a mantenere la propria forma per un certo tempo
ed il rapido ripristino delle normali condizioni di tonicità del liquido extracellulare permette la
riassunzione dell’acqua ed il ritorno al normale turgore.
Fig. 14 – La parete cellulare esercita un’azione di sostegno e di stabilità della forma della cellula
(A). Ad esempio, quando il contenuto si contrae per plasmolisi (B), la parete consente
alla cellula il mantenimento della propria forma ed impedisce che possa afflosciarsi,
almeno in tempi brevi, anche quando il protoplasto si contrae vistosamente per una
grande differenza di concentrazione tra i liquidi intra ed extracellulari (C).
- 25 -
Fiume Francesco
Fisiologia generale delle piante da orto
La figura 14 riporta lo schema della plasmolisi ed evidenzia il mantenimento della forma
originaria della cellula, nonostante la contrazione del contenuto.
Analogamente, grazie alla presenza della parete, le cellule vegetali possono sopportare le
continue variazioni di concentrazione dei liquidi esterni, come i cambiamenti di potenziale
osmotico del terreno (in cui le piante alloggiano le loro radici), come accade quando un terreno
secco viene bagnato dalla pioggia o da un intervento di irrigazione.
La parete cellulare difende la cellula dalle azioni traumatiche causate dagli eventi
meteorici (una pioggia battente, la grandine), dagli animali (particolarmente gli erbivori) e dai
parassiti che, per utilizzare le sostanze nutritive della cellula, debbono prima lesionarla e
distruggerla.
La parete svolge una funzione di sostegno della cellula e dell’intera pianta, poiché
permette il mantenimento di una certa pressione di turgore che consente alla pianta di
mantenersi eretta.
La mancanza d’acqua produce appassimento ed avvizzimento della pianta e conseguente
afflosciamento.
La parete cellulare può subire ispessimenti parziali (ad esempio soltanto in corrispondenza
degli angoli), come avviene nelle cellule dei tessuti collenchimatici e ciò conferisce grande
flessibilità e resistenza all’organo che di tali tessuti è dotato (ad esempio il picciolo delle foglie).
Ma può anche completamente lignificarsi (le cellule morte dei tessuti sclerenchimatici e del
legno delle piante arboree), conferendo alla struttura tissutale un’enorme resistenza e
consentendo alle piante di crescere fino a raggiungere dimensioni colossali (si pensi all’enorme
sviluppo di certe querce e dei baobab).
Un tempo era opinione diffusa che la parete cellulare fosse soltanto un prodotto esterno ed
inattivo del protoplasto, un semplice contenitore, mentre oggi sono state accertate delle funzioni
specifiche, essenziali non solo alla cellula ed al tessuto di cui essa fa parte, ma anche all’intero
organismo vegetale.
Nelle piante, infatti, le pareti cellulari intervengono in molti processi fisiologici e fra
questi l’assorbimento ed il trasporto dell’acqua e dei soluti, la traspirazione, il differenziamento,
le secrezione e l’escrezione di alcune sostanze e rappresentano siti di attività dei lisosomi e
dell’attività di degradazione e di decomposizione di alcune molecole.
Composizione chimica della parete cellulare
La composizione chimica della parete cellulare è complessa e variabile in rapporto all’età
di sviluppo della cellula ed al tipo di tessuto cui appartiene.
La parete è costituita da sostanze ternarie (formate da carbonio, idrogeno ed ossigeno),
quali la cellulosa, le emicellulose e le sostanze pectiche, e da sostanze quaternarie, cioè le
proteine.
La cellulosa rappresenta il polisaccaride della parete cellulare e della struttura
extracellulare delle piante più abbondante nel mondo vegetale.
Essa è costituita da molecole formate da lunghi polimeri di glucosio destrogiro (Dglucosio, che ruota a destra la luce polarizzata) con legami β(1→4).
Si ricorda, a tal proposito, che le forme α e β del D-glucosio non sono strutture a catena
aperta, ma strutture ad anello a sei atomi di carbonio, ottenute dalla reazione del gruppo alcolico
secondario dell’atomo di carbonio 5 con il gruppo aldeidico del carbonio 1.
- 26 -
Fiume Francesco
Fisiologia generale delle piante da orto
Queste strutture ad anello sono chiamate piranosiche perché derivate dal composto
eterociclico del pirano.
La forma α del D-glucosio presenta l’ossidrile del carbonio 1 a destra nella formula di
proiezione, mentre la forma β lo presenta a sinistra:
HCOH
⏐
HCOH
⏐
O
HOCH
⏐
HCOH
⏐
HC
⏐
CH2OH
HOCH
⏐
HCOH
⏐
O
HOCH
⏐
HCOH
⏐
HC
⏐
CH2OH
α-D-glucosio o
α-D-glucopiranosio
β-D-glucosio o
β-D-glucopiranosio
Le forme isomere di monosaccaridi, che differiscono tra loro soltanto per la
configurazione intorno all’atomo di carbonio carbonilico, sono dette anomeri e l’atomo di
carbonio è chiamato carbonio anomerico.
Naturalmente, le forme α e β presentano proprietà fisiche molto diverse.
La forma α devia il piano della luce polarizzata con un angolo di + 112,2°, ha un punto di
fusione di 146 °C, una solubilità, in 100 mL d’acqua, di 82,5 g, una velocità relativa
d’ossidazione da parte della glucosio-ossidasi pari a 100.
La seconda, la forma β ha una rotazione specifica di + 18,7°, fonde a 150 °C, ha una
solubilità di 178 g, ha una velocità di ossidazione da parte dello stesso enzima inferiore all’unità.
Queste differenze si ripercuotono profondamente e determinano le diversità tra l’amido e
la cellulosa.
Il peso molecolare minimo della cellulosa proveniente da fonti diverse è stato stimato tra
50.000 e 2.500.000 in specie diverse, equivalente (considerato che il peso molecolare del
glucosio è 180) a 278-14.000 residui di glucosio.
L’analisi della diffrazione ai raggi X indica una struttura fisica variabile, con catene
elementari riunite in fibrille in cui si distinguono le maglie cristalline separate le une dalle altre
da zone amorfe (figura 15).
In corrispondenza delle maglie, le catene sono saldamente unite, probabilmente, dai ponti
idrogeno e dalle forze di Van der Waals, mentre nelle zone amorfe i legami sono meno forti.
- 27 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 15 – Le pareti cellulari sono costituite da fili lunghi ed interconnessi, le macrofibrille, tanto
grandi (diametro di 0,5 μm e lunghezza di 4 μm) da poter essere viste al microscopio
ottico. Queste sono costituite da fasci di fibre più piccole, le microfibrille, visibili solo
al microscopio elettronico e larghe da 10 a 25 nm. Alcune porzioni delle microfibrille,
le micelle, sono disposte in maniera ordinata conferendo alla cellulosa delle proprietà
cristalline. Si osserva un frammento di micella con tratti di molecole di cellulosa con
una disposizione a maglia. Le micelle, rappresentate da catene di cellulosa, sono
chimicamente strutturate secondo la formula riportata in basso.
- 28 -
Fiume Francesco
Fisiologia generale delle piante da orto
Ogni microfibrilla risulta formata da un fascio di 40-300 lunghi polimeri (ogni polimero è
una molecola di cellulosa). La struttura particolarmente robusta e resistente delle microfibrille
dipende dai legami idrogeno che si stabiliscono tra i singoli polimeri di cellulosa (figura 16).
Questa particolare struttura conferisce alla cellulosa una resistenza pari a quella di una lamina di
acciaio di uguale spessore.
Fig. 16 – Due lunghi polimeri di cellulosa saldamente uniti dai legami idrogeno.
Tali caratteristiche fisiche, chimiche, strutturali e la presenza d’altri componenti
determinano anche il comportamento biologico della cellulosa.
Nelle pareti cellulari delle piante, le fibrille di cellulosa sono stipate fittamente in fasci
paralleli regolari intorno alla cellula e spesso sono sistemati in strati incrociati (figura 17).
Fig. 17 – Superficie della parete cellulare di una cellula vegetale nella quale è possibile
osservare le fibrille di cellulosa che si intersecano tra loro conferendo grande resistenza.
- 29 -
Fiume Francesco
Fisiologia generale delle piante da orto
Queste fibrille sono cementate tra loro da una matrice, che occupa gli spazi interfibrillari
ed è costituita da tre materiali polimerici: l’emicellulosa, la pectina e l’estensina. Il legno
contiene un’altra sostanza polimerica, la lignina, che costituisce quasi il 25% del suo peso secco.
Le emicellulose non sono simili strutturalmente alla cellulosa, ma sono così chiamate
perché, come questa, hanno, all’incirca, le stesse difficoltà di estrazione. Le emicellulose sono
polisaccaridi costituiti da xilani, β-glucani, xiloglucani, arabinogalattani, glucuronoarabinoxilani, ramnogalatturonani ed altri.
Gli xilani delle piante contengono legami β(1→4) tra residui di D-xilosio (un
monosaccaride aldeidico pentoso) e si formano da UDP-D-xilosio (uridina difosfato D-xilosio),
secondo il seguente schema:
UDP-D-xilosio + (D-xilosio)n → UDP + (D-xilosio)n + 1
catena preesistente
catena allungata
Si ricorda che l’uridina difosfato D-xilosio è un ribonucleoside 5’-fosfato, indicato nello schema
seguente, in cui la base azotata è l’uracile:
Nello schema successivo è rappresentato il legame β(1→4) che si stabilisce tra due molecole di
D-xilosio:
- 30 -
Fiume Francesco
Fisiologia generale delle piante da orto
ed è proposta la struttura di una porzione di una molecola di xiloglucano di una parete cellulare:
Il numero di residui di glucosio in una molecola completa di xiloglucano è compreso tra 15 e 30.
I legami glucosidici sono glucosio-β(1→4)-glucosio; xilosio-α(1→6)-glucosio; galattosioβ(1→2)-xilosio e fucosio-α(1→2)-galattosio.
- 31 -
Fiume Francesco
Fisiologia generale delle piante da orto
Va ricordato che, in generale, i polisaccaridi sono detti glicani e consistono in
monosaccaridi legati insieme da legami glucosidici. Essi sono classificati in omopolisaccaridi ed
eteropolisaccaridi, a seconda che siano costituiti da un solo tipo di monosaccaridi o da più tipi.
Anche se le sequenze di monosaccaridi e polisaccaridi possono in linea di principio variare
come quelle delle proteine, essi sono in effetti composti soltanto da pochi tipi di monosaccaridi
che si alternano in sequenze ripetitive. Orbene, i β-glucani sono polimeri del β-D-glucosio con
legami sia β(1→3), sia β(1→4) glucosidici. Il β-D-glucosio differisce dall’α-D-glucosio per la
posizione nello spazio, nella forma piranosica, dei gruppi ossidrili, come è evidenziato nel
seguente schema:
α-D-glucosio
D-glucosio
β-D-glucosio
Analogamente i galattani sono polimeri del galattosio. Nella composizione chimica della
parete cellulare sono presenti gli arabinogalattani, polimeri di L-galattosio con frequenti residui
laterali di D-arabinosio, singoli o in brevi catene. Nello schema di seguito riportato si ricordano
le formule di L-galattosio e di D-arabinosio, nelle forme α e β e si propone la struttura
dell’arabinogalattano:
α-L-galattosio
L-galattosio
- 32 -
β-L-galattosio
Fiume Francesco
α-D-arabinosio
Fisiologia generale delle piante da orto
D-arabinosio
- 33 -
β-D-arabinosio
Fiume Francesco
Fisiologia generale delle piante da orto
L’arabinogalattano può collegarsi, per un’estremità, con le proteine della parete cellulare ed in
particolare con residui di serina di glicoproteine ricche in idrossiprolina. Inoltre, questo
polimero plisaccaridico è caratterizzato dalla presenza di un residuo terminale di ramnosio di cui
si riportano le forme piranosiche di α-L-ramnosio e di β-L-ramnosio, con al centro la catena
lineare:
α-L-ramnosio
β-L-ramnosio
L-ramnosio
I glucurono-arabinoxilani sono costituiti da acido glucuronico, xilosio e arabinosio. Dei
primi due si riportano le strutture nello schema seguente:
acido β-D-glucuronico
β-D-xilosio
Le emicellulose, che insieme alla cellulosa vanno a costituire l’impalcatura della parete
cellulare, sono abbastanza eterogenee poiché, come si è visto, contengono un gran numero di
monosaccaridi polimerizzati in modo più o meno complesso con catene principali e secondarie
di varia lunghezza. Tuttavia, le emicellulose, al pari della cellulosa, sono ordinatamente
distribuite e si trovano a stretto contatto con le fibrille cellulosiche, costituendo un manicotto
amorfo e spingendosi negli spazi sottostanti, dove, in particolare gli arabinogalattani, si
collegano con le proteine.
Le sostanze pectiche sono delle macromolecole che derivano dalla polimerizzazione
dell’acido α-D-galatturonico, i cui gruppi carbossilici possono essere liberi o bloccati da gruppi
metossilici o acetilici. Nel primo caso si hanno gli acidi pectici che rappresentano una riserva di
cariche elettriche negative le quali possono essere neutralizzate da cationi calcio, magnesio e
potassio, per formare pectati di calcio, di magnesio e di potassio, rispettivamente, mentre nel
secondo caso si hanno le pectine in senso stretto. Queste catene sono abbastanza lineari e
ciascuna forma un omogalatturonano, in cui le molecole di acido α-D-galatturonico sono
- 34 -
Fiume Francesco
Fisiologia generale delle piante da orto
connesse da legami acido galatturonico-α(1→4)-acido galatturonico e presentano i gruppi
carbossilici bloccati da gruppi –COO- o –COCH3.
Le sostanze pectiche della parete cellulare in molti casi possono contenere molti altri
monosaccaridi, come il ramnosio, il galattosio, l’arabinosio, che vanno a costituire strutture più
complesse le quali possono essere esaminate mediante un’idrolisi parziale e successiva
separazione cromatografica. I ramnogalatturonani sono tra le sostanze pectiche più comuni. In
seguito si ricorda la formula di struttura dell’acido α-D-galatturonico e si riporta la struttura
polimerica di un omogalatturonano e del ramnogalatturonano (quest’ultima proposta da
Talmadge e collaboratori) della parete di una cellula vegetale:
acido α-D-galatturonico
omogalatturonano
ramnogalatturonano
- 35 -
Fiume Francesco
Fisiologia generale delle piante da orto
In particolare, i ramnogalatturonani formano una struttura a zig-zag nella quale è possibile
individuare 3 bracci che vanno a costituire una forma ad Y. Il primo è formato da una sequenza
di ramnosio, cui seguono approssimativamente 8 molecole di acido galatturonico, quindi una
molecola di ramnosio; i legami sono ramnosio-α(1→4)-acido galatturonico, acido galatturonicoα(1→4)-acido galatturonico che costituisce le regioni omogalatturoniche, acido galatturonicoα(1→2)-ramnosio. Il secondo braccio è dato dalla sequenza ramnosio-α(1→4)-acido
galatturonico-α(1→2)-ramnosio. Il terzo braccio è un galattano oppure un arabinano, formato
ciascuno da sequenze di galattosio ed arabinosio, ognuno dei quali si lega al carbonio 4 del
ramnosio che chiude la regione omogalatturonica.
Le sostanze pectiche possono facilmente essere estratte dalle pareti cellulari, per bollitura
o a seguito di un trattamento con sostanze chelanti, sotto forma di acidi pectici o pectati di
calcio, poco solubili e con tendenza a precipitare. E’ quello che capita quando si immerge un
frutto in acqua bollente, assistendo al suo rammollimento, per perdita di pectina dalle pareti
cellulari insieme alla trasformazione di zuccheri complessi in zuccheri semplici.
Le molecole di acido pectico possono aggregarsi in macromolecole di grandi dimensioni
per costituire una protopectina nella porzione di parete cellulare che separa due cellule figlie
ottenute da una divisione cellulare. La resistenza di queste pectine può variare con la formazione
o meno di ponti covalenti fra residui fenolici, spesso presenti nelle sostanze pectiche. In
particolare, tra i residui aromatici prevale l’acido ferulico che è noto perché agisce da coenzima
per l’IAA-ossidasi, un enzima che degrada e disattiva l’acido indolacetico (IAA), un vero
ormone auxinico delle piante capace di indurre crescita dei tessuti vegetali. Quindi, i legami
delle pectine possono influenzare la capacità di dilatazione ed estensione delle pareti cellulari e
regolare e controllare l’accrescimento delle cellule. Un aumento di acido ferulico incrementa
l’attività della IAA-ossidasi che inattiva e rimuove l’IAA, con conseguente riduzione
dell’accrescimento cellulare; al contrario, l’assenza o la diminuzione di acido ferulico riduce
l’attività e la sintesi della IAA-ossidasi e, quindi, per l’incremento di IAA si ottiene un
incremento di crescita cellulare con dilatazione e distensione della parete della cellula.
Un aspetto pratico della conoscenza della fisiologia delle sostanze pectiche della parete
cellulare riguarda la conservazione dei frutti. La maturazione dei frutti carnosi implica numerosi
cambiamenti, come la diminuzione del contenuto in clorofilla e la formazione di pigmenti che
ne cambiano il colore (invaiatura), ma soprattutto la diminuzione della consistenza, per cui il
frutto si ammorbidisce, fino al rammollimento ed al disfacimento. E’ questa una conseguenza
della scissione enzimatica della pectina, uno dei componenti della parete cellulare del frutto
carnoso, che causa lo scivolamento delle cellule l’una sull’altra. Uno dei principali problemi
relativo alla commerciabilità dei frutti carnosi (ad esempio le bacche di pomodoro) è il
mantenimento della consistenza per un periodo il più lungo possibile affinché il prodotto possa
giungere sulla tavola del consumatore in condizioni ottimali. In numerose esperienze,
trattamenti alle bacche di pomodoro con ascorbato di calcio per tempi brevi dell’ordine di
grandezza di 2-4 minuti ed alla dose di 0,4% possono prolungare e migliorare la conservazione,
riducendo a valori trascurabili gli scarti in magazzino. L’acido ascorbico, infatti, può abbassare
il pH del succo del frutto di pomodoro ed inibire, così, l’attività catalitica della poligalatturonasi
endogena, responsabile della degradazione delle pectine della parete cellulare durante il
processo di maturazione, mentre lo ione calcio può salificare, in seguito alla penetrazione nei
tessuti del frutto, gli acidi pectici che si formano durante i processi idrolitici correlati alla
- 36 -
Fiume Francesco
Fisiologia generale delle piante da orto
maturazione. Ne consegue un prolungamento della consistenza di una bacca (figura 18). A tal
proposito va anche detto che l’acido ascorbico, insieme ai cationi di ferro trivalente, è un
importante cofattore nella biosintesi dell’idrossiprolina, che è un aminoacido fondamentale di
quelle glicoproteine strutturali, le estensine, che formano tra loro legami intermolecolari in
grado di rafforzare la parete cellulare.
Fig. 18 – Stato di conservabilità di bacche di pomodoro trattate con due livelli quantitativi di
ascorbato di calcio, in confronto con il testimone non trattato (da Monopoli e
Scaramucci, 1982).
Tab. 1 – Aminoacidi standard. Denominazione e simboli.
Denominazione
dell’aminoacido
Alanina
Arginina
Asparagina
Acido aspartico
Cisteina
Glutamina
Acido glutamico
Glicina
Istidina
Isoleucina
Leucina
Lisina
Metionina
Fenilalanina
Prolina
Serina
Treonina
Triptofano
Tirosina
Valina
Simbolo a tre lettere
Simbolo a una lettera
Ala
Arg
Asn
Asp
Cys
Gln
Glu
Gly
His
Ile
Leu
Lys
Met
Phe
Pro
Ser
Thr
Trp
Tyr
Val
A
R
N
D
C
Q
E
G
H
I
L
K
M
F
P
S
T
W
Y
V
- 37 -
Fiume Francesco
Fisiologia generale delle piante da orto
Le proteine della parete sono principalmente costituite da polipeptidi ricchi di alcuni
aminoacidi, quali l’idrossiprolina, la serina, l’alanina e la glicina. In particolare, una diffusa
classe di proteine è rappresentatadall’estensina, una glicoproteina ricca in idrossiprolina che si
lega, come già visto, al galattosio terminale dell’arabinogalattano tramite gruppi di
idrossiprolina dei residui di serina.
L’idrossiprolina è un aminoacido cosiddetto raro o inconsueto o non standard, derivato
dalla prolina, uno dei 20 aminoacidi standard (tabella 1).
L’idrossiprolina, formata da una catena laterale idrofobica (un idrocarburo alifatico, indicato come gruppo
R) non polare, viene indicata nel seguente schema:
4-idrossiprolina
La sintesi dell’idrossiprolina avviene all’interno della cellula a partire da residui di
prolina.
La prolina viene sintetizzata da acido glutamico il cui gruppo γ-carbossilico viene
fosforilato dall’ATP o dal NADPH, analogamente a quanto avviene nella riduzione del 3fosfoglicerato a gliceraldeide 3-fosfato nell’inverso della glicolisi.
Si forma L-prolina che è anche un inibitore allosterico della reazione di trasformazione
del’acido glutamico in semialdeide dell’acido glutamico.
I residui di prolina che si formano, solo dopo la loro incorporazione nella catena
polipeptidica, per azione della prolina-4-monossigenasi danno luogo ai residui di 4idrossiprolina.
Questa ossigenasi a funzione mista (poiché richiede un primo substrato per catalizzare
l’inserzione di un atomo di ossigeno e di un secondo substrato che ceda gli elettroni per la
- 38 -
Fiume Francesco
Fisiologia generale delle piante da orto
riduzione ad acqua del secondo atomo della molecola di ossigeno) utilizza acido αchetoglutarico come coriducente:
Residuo di prolina + O2 + Acido α-chetoglutarico + CoA → Residuo di 4-idrossiprolina +
Succinil CoA + CO2 + H2O
Il Fe3+ e l’acido ascorbico sono necessari cofattori nella trasformazione dei residui di
prolina in residui di idrossiprolina. La somministrazione di ascorbato di calcio per via esogena a
frutti carnosi in via di maturazione, oltre a fornire ioni Ca2+, che vanno a costituire ponti tra le
molecole di pectina acida rafforzandone i collegamenti (figura 19), rende disponibile acido
ascorbico che agisce favorevolmente sulla sintesi di 4-idrossiprolina e, di conseguenza,
sull’arrichimento delle estensine nella parete cellulare. Ne deriva una più duratura stabilità della
stessa parete, con preservazione del frutto dai fenomeni di rammollimento e miglioramento della
consistenza nel tempo.
Le glicoproteine di parete, ricche in idrossiprolina, insieme alle pectine ed alle
emicellulose, si modificano ulteriormente nelle cisterne dei dittiosomi del reticolo
endoplasmico. Lo scheletro delle glicoproteine è costituito dal polipeptide che si forma dentro le
cisterne del reticolo endoplasmatico ruvido. Sui bordi delle cisterne dei dittiosomi, per
gemmazione, si formano numerose vescicole di secrezione, le quali sono incaricate del trasporto
delle proteine che sono finalmente scaricate nella parete cellulare in fase di crescita.
- 39 -
Fiume Francesco
Fisiologia generale delle piante da orto
Nella figura 19 è riportato lo schema di trasporto del polipeptide dai ribosomi del reticolo
endoplasmatico ruvido alle cisterne del dittiosoma. Precisamente, dal reticolo endoplasmatico
ruvido, provvisto di dittiosomi (a differenza del reticolo endoplasmatico liscio che ne è privo), si
staccano piccole vescicole di transizione che via via si accrescono e vanno a formare le cisterne
giovani che costituiscono la faccia in formazione del dittiosoma. Queste cisterne trasformano il
loro contenuto, diventano mature e formano, sulla faccia opposta del dittiosoma, la faccia di
maturazione formata da cisterne vecchie. Dalla cisterna vecchia prendono origine per
gemmazione delle vescicole più grandi, le vescicole di secrezione, nelle quali sono ormai
sintetizzate le glicoproteine di parete, che migrano fino alla superficie interna della parete
cellulare, nella quale versano il proprio contenuto.
Fig. 19 – Schema di trasporto del materiale della parete cellulare a partire dal sito di formazione:
lo scheletro del polipeptide si è formato nel reticolo endoplasmatico ruvido e viene
trasportato - prima mediante le vescicole giovani e poi attraverso le vescicole vecchie
secretorie, che non sono altro che le giovani vescicole che sono maturate e nelle quali
è avvenuta la modificazione secondaria delle glicoproteine di parete insieme alle
pectine ed emicellulose - fino alla membrana plasmatica, alla quale si fondono, per
versare il loro contenuto nella parete cellulare in accrescimento.
- 40 -
Fiume Francesco
Fisiologia generale delle piante da orto
L’assemblaggio dei diversi composti costituenti la parete cellulare è schematicamente
mostrato nella figura 20, in cui è evidente che le molecole di emicellulosa sono legate alla
superficie delle microfibrille di cellulosa da legami idrogeno, che alcune di queste molecole di
emicellulosa sono a loro volta legate trasversalmente a molecole acide di pectina da molecole
neutre di pectina, che le estensine circondano, collegano e bloccano le microfibrille irrobustendo
la parete.
Fig. 20 – Schema di interconnesione fra le microfibrille cellulosiche ed i diversi componenti
della parete cellulare.
Infine, si ritiene utile sintetizzare la composizione polimerica centesimale delle pareti di una
sospensione culturale di cellule vegetali che viene riportata nella tabella 2.
Tab. 2 - Composizione polimerica centesimale media delle pareti di cellule vegetali.
Componente della parete
Arabinano
3,6-arabinogalattano
4-galattano
Cellulosa
Proteina
Ramnogalatturonano
Tetra-arabinosidi attaccati all’idrossiprolina
Xiloglucano
Totale
- 41 -
Pareti cellulari (%)
10
2
8
23
10
16
9
21
99
Fiume Francesco
Fisiologia generale delle piante da orto
Stratificazione della parete cellulare
La parete cellulare non è una struttura statica ed inerte e neppure un semplice recipiente
contenente il protoplasma, avente la modesta funzione e tuttavia di vitale importanza, di indurre
consistenza e protezione, ma, al contrario, essa mostra un’intensa attività metabolica, interagisce
con gli altri comparti e componenti della cellula durante i processi di accrescimento e di
differenziamento, mostra un notevole dinamismo per i cambiamenti della composizione chimica
a seguito delle stimolazioni esterne ed ambientali.
Lo sviluppo delle cellule vegetali evidenzia che la parete è costituita da due strati: la
lamella mediana, più esterna alla cellula, che costituisce la sostanza intercellulare che
s’interpone tra due cellule contigue e la parete primaria, più interna. Molte cellule vegetali, in
particolare quelle che hanno completato il loro accrescimento, depositano, all’interno della
parete primaria, verso il citoplasma, un altro strato che va a costituire la parete secondaria.
La lamella mediana è formata da sostanze pectiche e da proteine e prende origine dalla
fusione delle vescicole della piastra cellulare durante la citodieresi, vale a dire quando si ha la
divisione del citoplasma durante la mitosi. Durante la formazione della piastra cellulare,
vescicole piene di pectine si staccano dai dittiosomi dell’apparato del Golgi e vanno a fondersi
lungo l’equatore per formare la nuova lamella mediana.
La parete primaria è lo strato della parete che si è formato durante la crescita cellulare,
per coinvolgimento di polisaccaridi non cellulosici secreti dalla cellula durante la formazione
della piastra cellulare per mezzo di vescicole dittiosomiche, all’interno e sulla lamella mediana.
Oltre alla cellulosa, alle emicellulose ed alle sostanze pectiche, le pareti primarie contengono
glicoproteine come le estensine. Le pectine conferiscono qualità plastiche e ciò consente alla
parete primaria di distendersi durante l’accrescimento della cellula.
Le cellule giovani in proliferazione, come quelle dei tessuti meristematici e quelle che
vanno a riparare lesioni e ferite con la formazione del callo di cicatrizzazione, e le cellule adulte
in attività metabolica, come quelle nelle quali avviene la fotosintesi, la respirazione e la
secrezione, sono provviste della sola parete primaria. Queste cellule, normalmente, sono capaci
di modificarsi morfologicamente, perdere la loro forma cellulare per la quale fanno parte di
tessuti specializzati e moltiplicarsi e differenziarsi in nuovi tipi cellulari. Le pareti primarie di
queste cellule hanno uno spessore non uniforme, poiché in alcune zone, denominate campi di
punteggiature primarie, diventano molto sottili. I plasmodesmi, filamenti di citoplasma che
collegano i protoplasti vivi di cellule contigue stabilendo tra loro un rapporto continuo di
interscambio, attraversano i campi di punteggiature primarie.
La parete secondaria viene prodotta dal protoplasto all’interno della parete primaria
quando la cellula vegetale ha terminato il proprio accrescimento e la stessa parete primaria non
ha più bisogno di estendersi in superficie. La presenza della parete secondaria si può considerare
un sintomo di invecchiamento della cellula, poiché il protoplasto, dopo la deposizione, spesso
muore. Con questo non significa che queste cellule hanno perduto le loro funzioni, anzi, al
contrario, vanno a costituire importanti tessuti specializzati nella conduzione della linfa e nel
sostegno della pianta e dei suoi organi.
La parete secondaria impartisce alla parete cellulare un notevole aumento della resistenza
meccanica e ciò impedisce l’eventuale schiacciamento delle cellule per effetto delle condizioni
di elevata tensione che spesso si verificano al loro interno a seguito del flusso di acqua che le
attraversa (cellule dei vasi dei fasci vascolari).
- 42 -
Fiume Francesco
Fisiologia generale delle piante da orto
La parete secondaria è ricca di cellulosa ed emicellulose e spesso contiene lignina ed altre
sostanze, mentre è priva di pectine e manca di glicoproteine (estensine). Le caratteristiche
funzionali della parete secondaria sono rappresentate dall’orientamento ordinato delle
microfibrille di cellulosa, che così possono costituire pareti multistratificate laminate, che si
osservano, ad esempio, nel legno compensato, o strutture geometriche come le tracheidi
spiralate dei tessuti xilematici; dalla presenza di lignina, che impartisce grande consistenza e
robustezza ai tessuti che possiedono cellule con pareti secondarie; dalla mancanza di sostanze
pectiche che induce rigidezza e limitata possibilità di distensione; dall’assenza delle estensine,
che sarebbero inutili qui localizzate, poiché la robustezza della parete è più che sufficientemente
impartita dalla lignina. Nella parete secondaria, le fibrille di cellulosa si orientano
differentemente tanto che diventa possibile distinguere tre strati (esterno, mediano ed interno)
ognuno dei quali è caratterizzato dal fatto che l’angolo, costituito dalla fibrilla e dall’asse
principale della fibra di cellulosa, ha un valore costante e specifico e viene usato in merceologia
per la valutazione delle qualità meccaniche del materiale. Le catene di cellulosa possono
integrarsi ed associarsi con lignina, cere, resine, tannini e pigmenti, in rapporto al tessuto ed alla
specie della pianta (figura 21).
Fig. 21 – Catene di cellulosa in strati successivi, in una tipica fibra con elevati depositi di parete
secondaria (tre strati). Gli strati sono costituiti da molecole di cellulosa che avvolgono
la cellula secondo anelli a spirale.
La parete secondaria non è mai depositata in corrispondenza dei campi di punteggiature
primarie della parete primaria, dove la parete cellulare permane molto sottile e consente d’essere
attraversata dai plasmodesmi. La parete secondaria si presenta, pertanto, caratterizzata da
depressioni ed assottigliamenti e punteggiature possono formarsi anche dove non si trovano i
campi di punteggiature primarie, in corrispondenza di una punteggiatura della cellula contigua.
- 43 -
Fiume Francesco
Fisiologia generale delle piante da orto
La lamella mediana e le due pareti primarie di due cellule vicine sono dette membrana della
punteggiatura, mentre una coppia di punteggiatura comprende le due punteggiature delle due
cellule contigue più la membrana.
Le cellule provviste di pareti secondarie possono avere una punteggiatura semplice ed una
punteggiatura areolata. In queste ultime, e sole in queste, la parete secondaria crea la cavità
della punteggiatura (figura 22).
Fig. 22 – Cellula del parenchima, fotosinteticamente attiva, con plasmodesmi in corrispondenza
dei campi delle punteggiature primarie, pareti primarie e lamelle mediane (A); cellule
provviste di pareti secondarie e punteggiature semplici (B); punteggiature semplici (C)
ed areolate (D) in cellule provviste di pareti secondarie.
Le punteggiature sono, pertanto, delle piccole aree della parete cellulare, il più delle volte
di forma circolare, dove non ha avuto luogo la formazione della parete secondaria (molto meno
permeabile all’acqua della parete primaria) e le cellule sono quindi separate soltanto dalla parete
primaria. Ciò consente il passaggio dell’acqua tra le trachee o le tracheidi e le cellule circostanti.
In particolare, le punteggiature areolate permettono il passaggio dell’acqua sia attraverso
le estremità, di solito inclinate, delle tracheidi, sia attraverso le pareti laterali di esse e dei vasi.
Le punteggiature areolate possono modulare il passaggio dell’acqua, nel senso che, nel caso di
un eccesso di pressione su uno dei due lati del tracheide, il toro ostruirà l’apertura
interrompendo il flusso.
Il toro è una specie di valvola che si forma per rigonfiamento della parete primaria al
centro della punteggiatura, laddove la parete secondaria si è fortemente assottigliata, che si
chiude quando la pressione da un lato della parete è maggiore che dall’altro.
Nella figura 23 è riportato il particolare schematico di una punteggiatura areolata di un
tracheide di conifera.
- 44 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 23 – A: Punteggiatura areolata di un tracheide; il toro consente l’apertura quando la
pressione sui due lati del tracheide è di uguale valore (in basso a sinistra) e si chiude,
interrompendo il flusso idrico, quando su uno dei due lati si determina un eccesso di
pressione (in basso a destra). B: Fotografia al microscopio elettronico a scansione dei
vasi xilematici del caule di cetriolo, caratterizzati da molte punteggiature (da
Troughton e Donaldson, 1972).
Fig. 24 – Fotografia, al microscopio elettronico, di plasmodesmi in una sezione longitudinale
(A) e trasversale (B), in un campo di punteggiature primarie di una cellula di una
foglia di granoturco (Raven et al., 2000).
- 45 -
Fiume Francesco
Fisiologia generale delle piante da orto
I plasmodesmi (figura 24), come già accennato, rappresentano l’unica soluzione di
continuità della parete cellulare e la sola via di collegamento intercellulare diretto, per il
trasporto di materiali da una cellula all’altra. Essi sono sottili canali contenenti filamenti di
citoplasma e si trovano sparsi nella parete cellulare o raggruppati in corrispondenza delle
punteggiature primarie. Hanno un diametro di 30-60 nm, sono delimitati dalla membrana
plasmatica ed attraversati da un tubulo del reticolo endoplasmico, detto desmotubulo, rimasto
intrappolato nella piastra cellulare durante la sua formazione, in corso di divisione cellulare. Va
anche puntualizzato, però, che i plasmodesmi si formano anche nelle cellule che non subiscono
la divisione.
Il trasporto di sostanze da una cellula all’altra attraverso i plasmodesmi è detto trasporto
simplastico e simplasto è l’insieme dei protoplasti interconnessi ed i loro plasmodesmi. Al
contrario, il trasporto apoplastico è il movimento dei fluidi (acqua e sostanze in soluzione)
lungo le pareti cellulari. Il continuum delle pareti cellulari va a costituire l’apoplasto che,
pertanto, circonda il simplasto.
I plasmodesmi permettono una comunicazione più efficiente e rapida di quella che avviene
con un passaggio indiretto attraverso la membrana plasmatica e la parete della cellula. Il
rifornimento nutritivo delle cellule e dei tessuti distanti dalle fonti di approvigionamento può
realizzarsi per semplice diffusione, o per flusso di massa attraverso i plasmodesmi, come pure
l’entrata e l’uscita di alcune sostanze dai vasi, che trasportano la linfa a lunghe distanze (xilema
e floema), può attuarsi attraverso i plasmodesmi.
La dimostrazione che i plasmodesmi collegano cellule adiacenti e consentono il passaggio
di sostanze da una cellula all’altra è data da una semplice esperienza. Se s’inietta nel citoplasma
di una cellula (ad esempio di uno stame) un colorante fluorescente (la fluoresceina sodica, verso
cui la membrana plasmatica è impermeabile), dopo due minuti diventano fluorescenti anche le
cellule adiacenti, segno, questo, che il colorante, non potendo attraversare la membrana
plasmatica, è passato soltanto attraverso le uniche soluzioni di continuità della membrana, i
plasmodesmi. Va tuttavia anche aggiunto che non è ancora noto se i plasmodesmi possono
regolare il trasporto intercellulare di sostanze dinamogene e plastiche, anche se essi potrebbero
disporre di apposite valvole in grado di farlo.
Attraverso i plasmodesmi avviene anche la diffusione di agenti infettivi delle piante come,
per esempio, i virus. Le particelle virali penetrano nelle cellule delle piante ospiti, si replicano, si
diffondono nelle cellule vicine ed attraverso il sistema vascolare infettano l’intera pianta
(infezione sistemica). Il passaggio dei virus da una cellula all’altra avviene attraverso i
plasmodesmi ed i movimenti delle particelle infettive sono resi possibili ed agevolati dalle
proteine di movimento (Movement Proteins) che sono codificate dagli stessi virus. Ancora poco
conosciute sono le modalità di diffusione dei virus a lunga distanza; tale diffusione,
probabilmente, si realizza attraverso l’interazione delle proteine del capside virale con prodotti
del floema dotati di elevata specificità.
Modificazioni e particolarità funzionali della parete della cellula vegetale
In relazione alle numerose funzioni della parete cellulare, questa può modificarsi nella sua
struttura in rapporto all’assorbimento ed alla circolazione dell’acqua e delle sostanze nutritive
nella pianta. Queste modificazioni possono avvenire per infiltrazione ed incrostazione di
sostanze polimerizzanti tra le fibrille di cellulosa, oppure per apposizione di materiali
impermeabili sul lato esterno della stessa membrana.
- 46 -
Fiume Francesco
Fisiologia generale delle piante da orto
L’arricchimento della parete cellulare in lignina avviene per incrostazione e ciò induce
alla cellula vegetale ed all’intera pianta un’enorme resistenza meccanica, di sostegno e
biologica.
Per questo processo di incrostazione di lignina, le piante possono assumere dimensioni
gigantesche e possono creare un efficiente sistema di conduzione della linfa che è lo xilema.
La lignina, dopo la cellulosa, è il più abbondante polimero che si trova nelle piante
superiori ed è presente in maggior quantità nelle piante legnose, dove si accumula nella lamella
mediana, nella parete primaria e nella parete secondaria dei vasi legnosi che, nel loro complesso,
vanno per l’appunto a costituire lo xilema.
La lignina s’infiltra fra le microfibrille di cellulosa rendendo la parete cellulare resistente
alle forze di compressione, mentre la cellulosa assolve la funzione di resistenza alle forze di
tensione durante l’accrescimento della cellula e sotto l’azione della pressione di turgore.
La lignina si presenta come una sostanza bruna ed amorfa e si evidenzia per la colorazione
rossa che assume quando è trattata con una miscela di fluoroglucinolo ed acido cloridrico.
Per il suo elevato peso molecolare (probabilmente oltre 10.000) e per i legami idrogeno
dei suoi gruppi ossidrili ai diversi carboidrati e soprattutto alla cellulosa in vari punti, la lignina
è difficilmente solubile.
Utilizzando il processo al solfito, è possibile estrarla dal legno a seguito di trattamenti con
soluzioni di bisolfito di sodio (NaHSO3) che degradano la lignina in sostanze più semplici
solubili in acqua.
Un altro metodo di estrazione della lignina è il processo kraft che consiste nella bollitura
della segatura di legno in una miscela di solfuro di sodio (Na2S), carbonato di sodio (Na2CO3)
ed idrossido di sodio (NaOH).
Con questo processo vengono liberati sostanze organiche dello zolfo, di odore penetrante,
mentre la lignina subisce un profondo processo di demolizione.
Sono questi due processi che vengono impiegati nell’industria cartaria per rimuovere la
lignina, la cui presenza darebbe luogo a carta di cattiva qualità per la colorazione giallastra. La
lignina rappresenta, così, un importante prodotto di scarto dell’industria della carta e della
cellulosa, la cui quantità annua da smaltire nel mondo ammonta a svariati milioni di tonnellate.
Come si è visto, è difficile estrarre la lignina senza scomporla, fino alla degradazione delle
sue unità di base e ciò ha reso finora impossibile definire esattamente la sua struttura chimica.
Le conoscenze che si hanno in proposito derivano dall’analisi di alcuni composti intermedi che
si formano nel ciclo biosintetico della lignina, da cui si è accertata l’esistenza strutturale di
alcuni alcoli aromatici, quali l’alcool coniferilico, l’alcool sinapilico e l’alcool p-cumarilico, in
particolare nelle piante legnose.
Le lignine di piante erbacee contengono grandi quantità di acidi fenolici, come l’acido pcumarico e l’acido ferulico i quali si legano fra loro con i rispettivi gruppi alcolici e con l’alcool
sinapilico, l’alcool p-cumarilico ed altri.
La via biosintetica che porta alla formazione di lignina è la biosintesi della fenilalanina
che dà luogo prima all’acido cinnamico, poi all’acido cumarico e quindi alla lignina.
Nello schema che segue sono indicati i vari passaggi che portano prima alla biosintesi
dell’acido corismico, poi della fenilalanina, dell’acido cinnamico, dell’acido cumarico,
dell’alcool cumarilico ed infine, per dimerizzazione e polimerizzazione di quest’ultimo, a
lignolo e lignina, rispettivamente; di quest’ultima si propone una struttura parziale:
- 47 -
Fiume Francesco
Fisiologia generale delle piante da orto
- 48 -
Fiume Francesco
Fisiologia generale delle piante da orto
L’acido corismico rappresenta il crocevia di diverse vie biosintetiche: quella del triptofano
e della tirosina. Dà, inoltre, origine a folati e ubichinoni. L’acido cumarico è il composto di
partenza nel processo di polimerizzazione del lignolo e della lignina, ma anche dell’acido
caffeico e delle cumarine.
La cutina e la suberina sono polimeri insolubili dei lipidi che fanno parte della struttura
della parete cellulare di molti vegetali. Essi costituiscono la matrice di composti lipidici a lunga
catena, rappresentati dalle cere le quali conferiscono alla parete cellulare caratteristiche
idrofobiche.
La cutina è un polimero eterogeneo costituito in prevalenza da diverse combinazioni di
composti di due gruppi di acidi grassi, con 16 e 18 atomi di carbonio, rispettivamente e con uno
o più gruppi ossidrilici. Sono, inoltre, presenti piccole quantità di composti fenolici.
- 49 -
Fiume Francesco
Fisiologia generale delle piante da orto
Chimicamente, per circa il 50%, la suberina è costituita da una complessa miscela di acidi
grassi a molti atomi di carbonio, di acidi grassi idrossilati, di acidi dicarbossilici e di alcoli a
lunga catena. Tutti questi composti presentano quasi sempre più di 16 atomi di carbonio. Il
restante 25-50% della suberina totale contiene composti fenolici, fra i quali l’acido ferulico è
quello più rappresentato. Per la consistenza della frazione lipidica esiste una certa somiglianza
fra cutina e suberina e le differenze sono dovute al tipo di acidi grassi presenti ed al fatto che
quest’ultima è provvista di una maggiore quantità della frazione fenolica.
Le cere sono costituite da diversi idrocarburi a lunga catena contenenti anche piccole
quantità di ossigeno. Molte cere contengono acidi grassi a molti atomi di carbonio, esterificati
con alcoli monoidrici a catena lunga, ma anche alcoli liberi, aldeidi e chetoni con 22-32 atomi di
carbonio ed idrocarburi con non oltre 37 atomi di carbonio.
La cutina e le cere o la suberina e le cere, ma particolarmente le cere, vanno a costituire
una barriera protettiva che ha lo scopo di prevenire e ridurre la perdita di acqua per
evaporazione e di isolare efficacemente organi e tessuti della pianta.
La cutina si deposita, per esempio, in piccoli strati nelle pareti cellulari delle cellule
epidermiche delle foglie e dei frutti per formare un tessuto protettivo, la cuticola, che limita
notevolmente la perdita d’acqua per traspirazione. In particolare, in corrispondenza delle aree
stomatiche esistono adeguati ispessimenti cuticolari che consentono il meccanismo di apertura e
chiusura degli stomi, correlato alle variazioni di turgore delle cellule di guardia. Senza la
cuticola, la traspirazione nella maggior parte delle specie vegetali sarebbe talmente rapida da
provocare la morte delle piante. La cuticola rappresenta, inoltre, un’importante barriera per la
protezione delle piante contro agenti patogeni ed eventi traumatici.
Normalmente la cutina e le cere vengono sintetizzate dall’epidermide e vengono poi
secrete sulla superficie. Le cere si trovano soprattutto all’esterno della cuticola, dove si
accumulano secondo vari modelli, ma possono anche con essa formare un miscuglio.
La cuticola assume un particolare interesse in agricoltura, nella pratica dei trattamenti
antiparassitari ed erbicidi, poiché, per la sua idrofobicità, respinge l’acqua di nebulizzazione del
fitofarmaco. Per permettere all’acqua del trattamento di bagnare le foglie è necessario
aggiungere al fitofarmaco un tensioattivo, detto bagnante, allo scopo di ridurre la tensione
superficiale del liquido.
La suberina partecipa alla struttura della parete delle cellule dei tessuti secondari di
rivestimento. Tali cellule, a seguito del deposito di suberina, vanno incontro ad un vero e
proprio isolamento, con la conseguenza che alla fine esse muoiono, costituendo un tessuto detto
sughero. Questo, formato di cellule morte e piene d’aria, ha una funzione d’isolamento e di
protezione molto efficace.
La suberina va ad impregnare la parete primaria di un particolare tipo di cellule che
costituiscono la banda del Caspary che si trova nell’endoderma, lo strato più interno della
corteccia primaria della radice, formato da cellule a stretto contatto fra loro, quindi prive di spazi
intercellulari. La banda del Caspary è un ispessimento nastriforme che si trova sulle pareti
anticlinali, cioè sulle pareti orientate perpendicolarmente alla superficie delle radici (pareti
radiali) e riveste un’importanza fisiologica fondamentale per l’intera pianta. In relazione al fatto
che l’endoderma è privo di spazi intercellulari e la banda del Caspary è impermeabile all’acqua,
tutte le sostanze del cilindro centrale della radice debbono obbligatoriamente entrare nei
protoplasti delle cellule dell’endoderma. Il passaggio dei liquidi e dei soluti può verificarsi
- 50 -
Fiume Francesco
Fisiologia generale delle piante da orto
attraverso le membrane plasmatiche o per mezzo dei plasmodesmi che mettono in
comunicazione le cellule dell’endoderma con le cellule della corteccia e del cilindro centrale.
La banda del Caspary impedisce, pertanto, il passaggio delle sostanze attraverso le pareti
cellulari e ciò è stato dimostrato nelle radici di granoturco alle quali era stato fatto assorbire
lantanio, il cui catione non è in grado di attraversare la membrana plasmatica. L’osservazione al
microscopio elettronico mostrava presenza di lantanio soltanto nelle pareti delle cellule della
corteccia, segno, questo, che le bande del Caspary dell’endoderma ne avevano impedito il
passaggio nel cilindro centrale. Nella figura 25 è visibile il tratto della parete cellulare, più
denso, dove è presente la banda del Caspary.
Fig. 25 – Sezione trasversale radiale della parete cellulare di due cellule contigue
dell’endoderma. E’ evidente la banda del Caspary in cui la parete è più densa e le
membrane plasmatiche vi aderiscono tenacemente.
Come è stato già accennato, il trasporto dell’acqua si realizza attraverso l’apoplasto o
attraverso il simplasto. Nel primo caso il movimento dell’acqua avviene attraverso le pareti
cellulari, mentre nel secondo caso il movimento si verifica da protoplasto a protoplasto
attraverso i plasmodesmi. Il trasporto idrico può anche essere misto, cioè via apoplasticasimplastica. L’acqua, pertanto, penetra dal terreno nel pelo radicale, attraversa, via apoplastica
prevalentemente, l’epidermide, la rizodermide e la corteccia per giungere alla banda del Caspary
dove il trasporto apoplastico è bloccato. La banda del Caspary obbliga l’acqua, che si muove
verso lo xilema, ad attraversare la membrana plasmatica ed il protoplasto delle cellule
endodermiche (trasporto simplastico) seguendo il passaggio plasmodesmico. Una volta
attraversata la banda del Caspary, l’acqua può ancora seguire la via apoplastica nel suo
movimento verso il cilindro vascolare (le trachee dello xilema). A livello delle cellule
dell’epidermide avviene l’assorbimento attivo degli ioni inorganici che, via simplastica, sono
trasportati attraverso la corteccia fino alle cellule parenchimatiche delle trachee nelle quali sono
immessi. La figura 26 mostra schematicamente la via di trasporto apoplastico e simplastico
dell’acqua e degli ioni inorganici dal terreno fino alle trachee.
- 51 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 26 – Trasporto apoplastico e simplastico dell’acqua e dei soluti attraverso
l’epidermide, la corteccia, l’endoderma e la banda del Caspary, il periciclo, fino ai
vasi xilematici (trachee). Notare le cellule della banda del Caspary con le pareti
ispessite ed impregnate di polimeri idrofobici (suberina) che hanno la funzione di
impedire il passaggio apoplastico dell’acqua e dei soluti dalla corteccia al cilindro
centrale della radice e viceversa.
Una particolare modificazione della parete cellulare si trova nelle cellule di trasferimento
(figura 27), comunemente indicate con il termine anglosassone di transfer cells. Osservate al
microscopio elettronico, queste cellule presentano una parete a superficie molto sinuosa e con
numerose introflessioni che aumentano enormemente la superficie della stessa parete e della
membrana plasmatica, consentendo capacità di trasporto molto elevate. Le cellule transfer,
infatti, attuano il trasferimento dei soluti a brevi distanze, sono molto diffuse nelle piante dove
svolgono la stessa funzione in ogni parte. In particolare, cellule parenchimatiche transfer hanno
la funzione di mediare, in parte, i processi di scambio con il floema (linfa “elaborata”) degli
elementi minerali e dei soluti organici presenti nel succo xilematico (linfa “grezza”). Si calcola
che nelle cellule di trasferimento la superficie di scambio possa decuplicarsi, a parità di volume
cellulare, con la conseguenza di un proporzionale aumento della velocità di trasporto. Le
transfer cells si trovano in associazione con lo xilema ed il floema di piccole nervature (ad
esempio negli ultimi rami di una nervatura fortemente ramificata) fogliari e cotiledonari oppure
delle tracce fogliari a livello dei nodi, nelle monocotiledoni e nelle dicotiledoni. Si trovano
altresì nei tessuti di alcuni elementi riproduttivi (placenta, sacco embrionale, endosperma), in
diverse strutture di secrezione (ghiandole, nettari), nella stele radicale e nei noduli delle radici
delle leguminose.
- 52 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 27 – Cellule di trasferimento nella nervatura minore della foglia di una pianta della famiglia
delle Asteracee. Le cellule, che si presentano inscurite per il loro citoplasma molto
denso, hanno una parete caratterizzata da sinuosità e numerosissime introflessioni che,
aumentando la superficie, velocizzano il trasporto dell’acqua e dei soluti (da Gunning e
Steer, 1996).
La parete cellulare può subire interessanti modificazioni per opporsi all’ingresso di
microrganismi dannosi alle piante che possono causare l’insorgenza di patologie fino a causarne
la morte. Le piante sono esposte all’aggressione di innumerevoli e differenti specie di agenti,
causa di infezione ed infestazioni, rappresentati da virus, batteri, funghi, nematodi, acari, insetti.
In particolare, un microrganismo per infettare una pianta deve superare le sue barriere difensive
ed una volta che ciò è avvenuto la stessa pianta può dar luogo a delle risposte fisiologiche che
hanno lo scopo di bloccare la diffusione del patogeno ed evitare di soccombere. Il patogeno può,
a sua volta, mettere in atto nuovi meccanismi di virulenza per superare le barriere difensive
frapposte dalla pianta. Viene, in tal modo, ad instaurarsi tra pianta e patogeno un complesso
rapporto di azioni e reazioni incessantemente presente, con una continua evoluzione dei processi
di patogenesi e di difesa delle piante. Ciò porta ad affermare che la sensibilità delle piante alla
malattia è un’eccezione e non la norma ed il danno che l’organismo vegetale può ricevere
dipende non soltanto dal patogeno, ma anche dall’ambiente, dalla condizione fisiologica della
pianta e dall’interazione di tutti questi aspetti.
Un patogeno può penetrare nella pianta attraverso aperture naturali o ferite oppure
attraverso le pareti cellulari dopo esserne venuto a contatto ed averle degradate o indebolite. La
maggior parte dei batteri e dei funghi fitopatogeni studiati è in grado di produrre, in vitro,
enzimi che degradano le pareti cellulari, ma per dimostrare il loro ruolo nella patogenesi è
necessario accertarsi della loro presenza nella pianta ammalata. In queste piante è stata
individuata la presenza di cellulasi, emicellulasi e diversi tipi di pectinasi che sono in grado di
operare la distruzione delle pectine della lamella mediana e conseguente distruzione della parete
cellulare. I più importanti enzimi pectici delle malattie sono le endopoligalatturonasi che
- 53 -
Fiume Francesco
Fisiologia generale delle piante da orto
spezzano in modo casuale l’acido poligalatturonico invece di distaccare i singoli residui
terminali.
La domanda che sorge è il motivo per il quale questi enzimi della parete, prodotti
abbondantemente in coltura dai microrganismi patogeni, sono in grado di degradare soltanto le
pareti cellulari degli ospiti sensibili alla malattia e non quelle di tutte le piante. La risposta può
essere data considerando diversi meccanismi biochimici messi in atto dalle piante. La cellula,
per esempio, può produrre degli inibitori delle endopoligalatturonasi anche se, tuttavia, tali
inibitori non sono sufficientemente diversi nelle varie specie e varietà di piante da poter spiegare
il fatto che lo stesso patogeno è in grado di attaccare soltanto certe specie o varietà e non altre.
Anche le modificazioni nelle pareti cellulari dei tessuti più vecchi, come la formazione di pectati
di calcio, possono aumentare la resistenza agli enzimi degradanti le pectine e ciò spiega la
resistenza ad alcune malattie per l’invecchiamento della pianta. Un altro meccanismo di
resistenza delle piante è la loro capacità di produrre chitinasi che demoliscono le pareti cellulari
dei patogeni fungini.
Vi sono, inoltre, prove sperimentali su una precoce reazione di riconoscimento del
patogeno da parte della pianta, che può precedere la fase di penetrazione. Così, nella malattia del
mal del colletto del tabacco, causata dal fungo Phytophthora parasitica pv nicotiana, quando il
promicelio del conidio viene a contatto con le pareti cellulari dell’epidermide radicale delle
varietà resistenti, si ha la formazione di vescicole originate dal reticolo endoplasmico e dai
dittiosomi, il cui contenuto viene liberato attraverso la membrana plasmatica ed accumulato
contro la parete cellulare. Quando l’ifa penetra nella cellula questa muore e le sostanze
contenute sulla parete inducono l’inibizione del fungo. Ciò non accade per le cellule sensibili le
quali non reagiscono così rapidamente e consentono al fungo di propagarsi ed invadere l’intero
tessuto. Questa precoce specificità è causata dalla presenza di proteine simili nell’ospite e nel
patogeno. Nelle coppie ospite resistente-patogeno è stato accertato che le proteine dei due
individui sono diverse. Tuttavia, il meccanismo del riconoscimento o non riconoscimento delle
proteine non è stato ancora completamente chiarito, né è stata accertata la funzione intracellulare
delle stesse proteine.
Un altro modello di riconoscimento, questa volta fondato sulla chimica dei carboidrati
delle pareti cellulari, si ha, per esempio, quando due individui di sesso opposto (mating types)
del lievito Hansenula wingei si incontrano, riconoscendo i carboidrati delle rispettive pareti
cellulari. Anche i batteri azotofissatori del genere Rhizobium, che provocano la formazione dei
tubercoli radicali nelle leguminose, hanno sulla loro parete cellulare carboidrati differenti da
quelli dei ceppi che non inducono la formazione dei tubercoli. Anche i granelli di polline che
germinano ed introducono il tubetto pollinico soltanto negli stigmi della stessa specie è stata
attribuita al tipo di carboidrati presenti sulla loro parete.
In linea del tutto generale, alcuni componenti delle pareti cellulari producono messaggi
chimici capaci di modulare processi fisiologici di fondamentale importanza. Ad esempio, le
oligosaccarine, costituite da oligosaccaridi derivati da glucani e pectine, possono essere
rilasciate dalle pareti cellulari in condizioni patologiche e stimolare reazioni di difesa e regolare
alcuni fenomeni morfogenetici; gli oligogalatturonidi hanno la capacità di inibire alcune risposte
ormonali. In particolare, alcuni oligogalatturonidi derivati dalle stesse pectine delle pareti
cellulari delle piante, degradate dalle poligalatturonasi fungine, possono agire come elicitori,
sostanze, cioè, che durante l’interazione pianta-patogeno, inducono nella pianta l’attivazione di
nuovi processi biochimici, con produzione di sostanze che difendono l’organismo vegetale
- 54 -
Fiume Francesco
Fisiologia generale delle piante da orto
dall’aggressione del patogeno. Secondo la teoria del gene per gene è previsto il riconoscimento
da parte di un recettore, codificato dal gene per la resistenza della pianta, di un elicitore prodotto
dal gene per l’avirulenza del patogeno.
Il prodotto genico del patogeno funziona per evocare la resistenza da parte della pianta
ospite e, più specificamente, esso è un enzima coinvolto nella sintesi dello stesso elicitore.
Alcuni esempi di elicitori e recettori di membrana sono: l’oligosaccarina che è un recettore da
plasmalemma di soia per un β-1,3-β-1,6-eptaglucoside (con 7 monomeri di glucosio o anche
frammenti più grandi) prodotto dalle pareti del ficomicete Phytophthora megasperma, che
induce reazione di ipersensibilità; la chitina fungina ed i suoi frammenti (a base di N-acetilglucosammina) stimolano la produzione di fitoalessine (composti fenolici, o terpenoidi, o
alcaloidi, dotati di proprietà inibitorie, derivati dal metabolismo secondario, vale a dire la cui
sintesi è indotta durante il processo di infezione) in colture cellulari e danno luogo a reazione di
ipersensibilità in alcune piante, il cui recettore nel plasmalemma è stato purificato; le proteine
elicitrici di reazione ipersensibile come le harpine di diversi batteri fitopatogeni; peptidi prodotti
dei geni di avirulenza, come quelli del Deuteromicete, Ifale, Dematiaceo Cladosporium fulvum
(così, il peptide AVR9 provoca necrosi fogliare nelle cultivar di pomodoro che contengono il
sistema di resistenza nel gene Cf9, che codifica la proteina Cf9, la quale ha la struttura di una
protein-chinasi recettoriale).
Infine, molto studiato in Italia è il gene RPS2 di Arabidopsis thaliana, che conferisce
resistenza verso Pseudomonas syringae (purché questi abbia il corrispondente gene di
avirulenza), la cui proteina RPS2 è transplasmalemma, ha un dominio interno ed un dominio
recettoriale esterno per legarsi con il prodotto batterico ed è simile alla regione del ligando dei
recettori animali, mentre nella sua porzione esterna è simile all’inibitore PGIP della
poligalatturonasi.
Proteine di membrana simili, ma talvolta prive della regione extracellulare, sono
conosciute per molte altre piante.
Nello schema seguente sono riportati le formule di alcune oligosaccarine, quali un
eptaglucano (derivato dalla parete fungina di Phytophthora megasperma pv glycinea) ed un
oligogalatturonide (derivato da pectine vegetali e degradato da poligalatturonasi fungine), che
possono agire come elicitori:
Eptaglucano
- 55 -
Fiume Francesco
Fisiologia generale delle piante da orto
Oligogalatturonide
In sintesi è possibile affermare che le piante hanno evoluto la capacità di percepire la
presenza dei fitopatogeni e di trasmettere questa informazione all’interno della cellula allo scopo
di innescare una serie di risposte in grado di bloccare il processo d’infezione. Per l’attivazione di
questi meccanismi è importante il riconoscimento del binomio pianta-patogeno che viene
mediato da elicitori prodotti dal patogeno, dalla pianta o da ambedue. Queste molecole elicitrici
interagiscono a livello delle superfici di contatto tra i due simbionti antagonisti (ospite e
patogeno) per innescare la via della trasduzione del segnale che produce le risposte di difesa
della pianta. La maggior parte dei funghi fitopatogeni deve superare la barriera rappresentata
dalla parete vegetale e per questo motivo produce una serie di enzimi che degradano i
polisaccaridi della parete cellulare. L’azione delle endopoligalatturonasi (PG), enzimi che
idrolizzano l’omogalatturano della componente pectica della parete cellulare della pianta, è
sovente un prerequisito per la degradazione della parete della cellula vegetale da parte di altri
enzimi fungini. La PGIP (polygalacturonase-inhibiting protein) è un inibitore proteico delle
poligalatturonasi ed è presente nelle pareti cellulari di molte dicodiledoni. La formazione del
complesso PG-PGIP, inibendo l’attività dell’enzima fungino, promuove il rilascio di
oligogalatturonidi con grado di polimerizzazione da 10 a 14, attivi come elicitori delle risposte
di difesa. La PGIP risulta essere la prima proteina, identificata nelle piante, appartenente alla
superfamiglia di proteine caratterizzate dalla presenza di moduli ripetuti ricchi in leucina (LRR).
L’analisi della struttura primaria della PGIP ha messo in evidenza un dominio strutturale
formato dalla ripetizione in tandem di 10,5 moduli, ciascuno derivato dalla modificazione di un
peptide di 24 aminoacidi in cui è mantenuta una distribuzione regolare dei residui di leucina ,
responsabile della sua interazione con le PG fungine. Le proteine con strutture LRR, distribuite
in modo ubiquitario negli organismi viventi, pur svolgendo differenti funzioni, sono
specializzate per le interazioni proteina-proteina. La somiglianza strutturale delle regioni LRR
riflette l’evoluzione convergente di questo dominio proteico costantemente mantenuto a seguito
di pressione selettiva. L’isolamento di numerosi geni di resistenza ha rilevato che alcuni di
questi codificano proteine con strutture LRR, omologhe alla PGIP. Questa osservazione indica
che le PGIP sono membri di una famiglia di proteine specializzate nell’interazione tra pianta e
patogeni. Le PGIP sono il prodotto di geni (pgip) organizzati in famiglie multigeniche. Ad
esempio, alcuni membri della famiglia pgip di Phaseolus vulgaris sono stati clonati e
caratterizzati ed hanno mostrato che i loro prodotti differiscono soltanto di otto aminoacidi e
possiedono differenze significative nel pattern di espressione e nella capacità di inibire le
diverse PG fungine.
Sono state descritte alcune specifiche funzioni della parete cellulare e come essa è
implicata nei fenomeni di resistenza alle malattie delle piante. Ma una specifica struttura della
parete cellulare è possibile osservarla nei granuli di polline dell’antera fiorale. E’ noto che
l’antera, dopo la differenziazione, è formata da cellule uniformi ad eccezione dell’epidermide.
Poi diventa possibile la distinzione di quattro gruppi di cellule fertili (cellule sporigene),
ciascuno dei quali è circondato da diversi strati di cellule sterili che originano la parete
- 56 -
Fiume Francesco
Fisiologia generale delle piante da orto
dell’antera. Lo strato più interno della parete dell’antera si chiama tappeto ed include le cellule
nutrici che forniscono sostanze nutritive alle microspore che si sviluppano. Le cellule sporigene
diventano microsporociti e subiscono una divisione meiotica (figura 28 A). Ogni microsporocito
diploide dà luogo a quattro microspore unicellulari aploidi (tetrade). Ogni divisione meiotica del
nucleo può essere seguita dalla formazione della parete (nelle monocotiledoni), oppure ciascun
protoplasto può contemporaneamente rivestirsi della parete cellulare dopo la seconda divisione
meiotica (nelle dicodiledoni). Si sono, così, formati i granuli di polline, i quali sviluppano una
resistente parete esterna (l’esina) ed una parete interna di natura cellulosica (l’entina). L’esina è
dura e resistente, spesso è provvista di rilievi che si presentano come una scultura più o meno
elaborata, è prevalentemente costituita da sporopollenina, un polimero contenente soprattutto
carotinoidi, e deriva in parte dal tappeto ed in parte dalle microspore (figura 28 B). L’entina è
depositata dai protoplasti delle microspore ed è composta da cellulosa e pectina.
Fig. 28 – Sezione trasversale di antera immatura del genere Lilium in cui sono visibili le quattro
sacche polliniche dell’antera, contenenti i microsporociti circondati dal tappeto (A).
Caratteristica scultura, osservata al microscopio elettronico a scansione, sull’esina di
polline del genere Lilium (B).
La struttura della parete assume un ruolo di particolare importanza nelle cellule di guardia,
a proposito della mobilità degli stomi. Questi subiscono, come già accennato, continui
movimenti in rapporto alle variazioni di turgore delle cellule di guardia. Quando in queste
cellule si verifica un aumento di concentrazione, per accumulo di soluti, si determina
un’apertura degli stomi, in relazione al maggior turgore delle cellule di guardia, per richiamo di
acqua nel loro interno, rispetto alle cellule epiteliali circostanti. Il meccanismo inverso provoca
la chiusura degli stomi, per la diminuzione della pressione di turgore delle cellule di guardia,
quale conseguenza di una fuoriuscita di acqua per l’aumento di diluizione. Il turgore delle
cellule di guardia aumenta o diminuisce, pertanto, per un meccanismo endosmotico od
esosmotico, secondo un gradiente di potenziale idrico dovuto al catione potassio. Lo ione K+,
- 57 -
Fiume Francesco
Fisiologia generale delle piante da orto
misurato quantitativamente con un microelettrodo specifico per questo ione posizionato nel
vacuolo delle cellule di guardia di stomi aperti, è stato trovato sempre ad elevata concentrazione,
mentre diminuisce in concomitanza alla chiusura stomatica. L’aumento di concentrazione in
potassio nelle cellule di guardia causa un passaggio osmotico di acqua dall’esterno verso il
vacuolo, con aumento di turgore ed apertura degli stomi; la diminuzione di concentrazione di
potassio induce un passaggio osmotico di acqua dall’interno del vacuolo verso l’esterno, con
riduzione del turgore cellulare e chiusura stomatica. Il gradiente di potassio tra le cellule di
guardia e le cellule adiacenti (6 cellule sussidiarie, di cui 4 laterali e 2 terminali) si modifica
rapidamente, generando un movimento idrico, per osmosi, che induce modificazioni del turgore
cellulare. L’abbondante movimento del catione potassio, di carica positiva, necessita di anioni,
di carica negativa, onde neutralizzare elettricamente il processo ed a tale scopo provvedono gli
anioni cloridrico (Cl¯) e malico (COOH-CH2-CHOH-COO¯). Nella figura 29 sono rappresentate
le variazioni di concentrazione di K+ nelle cellule di guardia e sussidiarie di un apparato
stomatico e nelle adiacenti cellule epidermiche.
Fig. 29 – Concentrazioni di potassio nelle cellule di guardia e sussidiarie (laterali e terminali) di
uno stoma fogliare ed in quelle epidermiche adiacenti, in un apparato stomatico chiuso
(A) ed aperto (B).
Dopo questa premessa ci si domanda quale ruolo gioca la struttura della parete delle
cellule di guardia nel movimento stomatico. Due tipi di forze inducono la curvatura delle cellule
di guardia, durante l’aumento del turgore e del loro volume, costringendo lo stoma ad aprirsi. La
prima è determinata dall’orientamento radiale (detto micellazione radiale) delle microfibrille
cellulosiche della parete delle cellule di guardia e dà luogo ad una costrizione che obbliga le
cellule stesse ad allungarsi, senza espandersi lateralmente. La seconda forza è posizionata
all’estremità delle due cellule di guardia, nel punto dove esse reciprocamente si toccano e
mettono in comune la loro parete, immobilizzando questa porzione condivisa e facendole
conservare una lunghezza costante durante l’apertura e la chiusura dell’apertura stomatica.
L’aumento del turgore cellulare spinge la parete esterna ad incurvarsi verso l’esterno, proprio
perché le estremità in comune, inferiori e superiori, sono fisse, cioè non possono spostarsi.
Contemporaneamente al movimento di curvatura della parete esterna delle cellule di guardia
(mentre le estremità sono fisse), la micellazione radiale trasmette un analogo movimento alla
- 58 -
Fiume Francesco
Fisiologia generale delle piante da orto
parete che delimita la rima stomatica, ossia alla parete ventrale dell’apertura stomatica. Nella
figura 30 viene schematicamente riprodotto il meccanismo di apertura e chiusura degli stomi e,
ricorrendo all’uso di palloncini, viene dimostrato che la simulazione della micellazione radiale
permette una maggiore apertura degli stomi.
Fig. 30 – Schema di una coppia di cellule di guardia: A) disposizione radiale delle microfibrille
di cellulosa nelle cellule di guardia; B) palloncini fissati insieme alle due estremità,
tenute immobili durante il rigonfiamento; C) palloncini gonfiati sprovvisti dell’effetto
della micellazione radiale; D) palloncini gonfiati ai quali sono state applicate strisce di
carta adesiva per riprodurre l’effetto della micellazione radiale. L’apertura che si viene
a stabilire tra i due palloncini (paragonabile all’apertura stomatica) è maggiore rispetto
a quello senza strisce adesive (C).
Crescita della parete cellulare
L’accrescimento della parete cellulare avviene nella doppia direzione dello spessore e
della superficie ed ha lo scopo di consentire la crescita della cellula vegetale.
La crescita della parete richiede l’aumento della sintesi delle proteine, della respirazione e
dell’assorbimento di acqua da parte della stessa parete cellulare. E’ necessario, tuttavia,
distinguere la crescita della parete cellulare che avviene per apporto di nuovo materiale
sintetizzato, dal più semplice rilassamento strutturale della parete che ha lo scopo di aumentare
l’estensibilità della stessa parete (che diventa più plastica) allorquando la cellula aumenta,
accrescendosi, le proprie dimensioni.
Va anche chiarito che i due aspetti sono strettamente correlati, poiché prima aumenta
l’estensibilità della parete, poi si verifica la penetrazione di acqua nella cellula provocando la
distensione della parete ed, infine, si realizza la sintesi di nuovo materiale che stabilizza la
parete distesa e la ricostituisce in modo che possa distendersi ulteriormente.
Fin dal 1971 era noto, presso il laboratorio di Cleland a Seattle, il meccanismo biochimico
dell’azione auxinica sulla distensione cellulare. L’auxina non agiva direttamente sulla parete
cellulare, bensì nel citoplasma o a livello del plasmalemma e si riteneva che dovesse esistere una
relazione tra il citoplasma e la parete. In altri termini l’auxina avrebbe dovuto attivare un fattore
in grado d’indurre l’estensione della parete.
Fin dagli anni trenta l’olandese Heyn evidenziò l’effetto dell’acido 3-indolacetico sulle
pareti cellulari di tessuti sensibili che, sotto l’azione auxinica, modificava i legami tra le
macromolecole delle pareti, che si ricostituivano in funzione delle forze che venivano esercitate
sulle stesse pareti. Inoltre, un trattamento con soluzioni acide rendeva le pareti più plastiche, con
aumento della velocità di crescita, mentre, al contrario, la neutralizzazione della soluzione
induceva un blocco della crescita.
- 59 -
Fiume Francesco
Fisiologia generale delle piante da orto
Nel 1973 Cleland e Marrè, indipendentemente, scoprirono che l’acidificazione delle pareti
assume enorme importanza nella distensione della parete cellulare e che le auxine hanno proprio
la funzione di attivare la pompa protonica del plasmalemma, con estrusione di H+ ed
acidificazione dell’apoplasto.
Dal 1973 le osservazioni iniziali sulla protrusione di protoni sono state approfondite,
permettendo di accertare che il fattore della distensione plastica della parete cellulare è da
individuare certamente negli ioni idrogeno.
Più recentemente sono state scoperte alcune proteine, che prendono il nome di espansine,
aventi la funzione di rendere instabili i ponti idrogeno delle macromolecole di cellulosa,
agevolando, così, la cedevolezza plastica della parete cellulare.
L’accrescimento vero e proprio della membrana cellulare avviene con la sintesi di nuovo
materiale che viene riversato nell’apoplasto per esocitosi.
Una parte, la maggiore, delle nuove microfibrille si dispone, strato su strato, al di sopra di
quelle precedentemente formate, mentre la restante parte può essere inserita nella preesistente
struttura della parete. Le microfibrille della parete primaria formano, all’inizio, un reticolo
irregolare, con una predominante orientazione trasversale, poi, quando la superficie della parete
aumenta, le microfibrille tendono a disporsi longitudinalmente o parallelamente al diametro di
maggiore dimensione della cellula.
La sintesi della cellulosa non è ancora completamente nota. E’, tuttavia, abbastanza
evidente che le microfibrille di cellulosa sono sintetizzate sulla superficie della cellula ad opera
di un complesso enzimatico cellulosa sintasi incluso nella membrana plasmatica e che
l’orientazione delle microfibrille è controllata dai microtubuli situati subito sotto il
plasmalemma.
La cellulosa sintasi è presente fra i due strati (quello esterno, subito sotto la parete
cellulare, e quello interno, a contatto col citoplasma), di cui è costituito il plasmalemma, sotto
forma di globuli e rosette.
Il donatore di unità di β-D-glucosio per la sintesi di polimeri del tipo della cellulosa è
l’UDP-glucosio (uridina 5’-α-D-glucopiranosil-difosfato), insieme a GDP-glucosio (guanosina
difosfoglucosio). UDP-galattosio, UDP-acido galatturonico, UDP-acido glucuronico, UDPxilosio, UDP-L-ramnosio, UDP-L-arabinosio sono tutti presenti nelle cellule vegetali e sono tutti
effettivi donatori di zuccheri per la produzione, in vitro, dei corrispondenti polisaccaridi della
parete cellulare.
Va anche detto che la cellulosa sintasi è un complesso enzimatico abbastanza instabile,
tanto che dopo l’isolamento non produce più cellulosa (con legami β-1,4) ma callosio (β-D-1,3glucano).
Le sostanze pectiche, le emicellullose e le glicoproteine sono i componenti della matrice e
sono portati alla parete cellulare, come già visto in precedenza, mediante le vescicole dei
dittiosomi.
Il tipo di composto della matrice, sintetizzato e secreto dalla cellula in un determinato
momento, dipende dallo stato di sviluppo.
Le pectine sono tipiche delle cellule in via di sviluppo, mentre le emicellulose prevalgono
in quelle adulte che hanno terminato l’accrescimento.
Sono ben note le reazioni con le quali la maggior parte degli zuccheri-nucleotidi possono
essere sintetizzati da UDP-glucosio, di cui si riporta la struttura chimica:
- 60 -
Fiume Francesco
Fisiologia generale delle piante da orto
UDP-glucosio (UDPG)
Per sfortuna si è ancora lontani dal capire come queste reazioni avvengono nella cellula
vegetale ed in particolare che cosa controlli la produzione dei diversi tipi di polimeri nelle
differenti specie. Come le proteine entrano nelle pareti e qual è il loro specifico ruolo sono
ancora problemi in corso di soluzione. Molto importante è conoscere che cosa stabilisce il
modello di deposizione delle microfibrille di cellulosa, poiché è questo modello che controlla la
direzione dell’accrescimento cellulare ed influisce sulle forme finali delle cellule e degli organi
delle piante.
In conclusione, la parete cellulare, insieme ai vacuoli, è l’aspetto caratteristico delle
cellule vegetali, costantemente presente nei tessuti vegetativi delle alghe verdi, delle briofite e
delle piante superiori vascolari. Sarebbe sorprendente se una caratteristica così universalmente
diffusa, come la parete cellulare, non avesse un significato profondo. Se la parete cellulare non
fosse di grande importanza per le piante, nel corso dell’evoluzione sarebbe stata eliminata,
almeno in qualche specie vegetale. Ma non esistono eccezioni. Nelle foglie, nei fusti, nelle
radici e nei fiori di tutte le specie vegetali, tutte le cellule adulte sono provviste di parete. Il
vacuolo per realizzare una struttura rigida ha bisogno di una parete cellulare relativamente poco
elastica, con un’elevata resistenza alla tensione. A ciò provvede la cellulosa, di cui è in parte
costituita la parete, dotata di una resistenza alla tensione paragonabile a quella di un filo
d’acciaio armonico. L’acqua da sola è troppo fluida per essere un materiale strutturale, ma
quando è racchiusa da una membrana e da una parete la sua incomprimibilità diviene molto
importante. In tali condizioni anche l’aria diventa un materiale strutturale quando è compressa
all’interno di un recipiente flessibile, come nel caso dei pneumatici. In una cellula vegetale le
pareti di cellulosa forniscono una resistenza alla trazione e l’acqua una resistenza alla
compressione ed insieme formano una struttura rigida e resistente. Per rifarsi ad un caso pratico
che tutti hanno toccato con mano, un pallone da calcio riempito d’acqua ha le caratteristiche di
- 61 -
Fiume Francesco
Fisiologia generale delle piante da orto
una roccia. Questa rigidità strutturale delle cellule turgide spiega perché le foglie, i cauli erbacei
e le piante in generale si mantengono rigidi e per quale motivo essi si afflosciano quando le
cellule perdono acqua ed appassiscono. Le fibrille di cellulosa non hanno la capacità, con la sola
resistenza alla tensione, di mantenere la pianta eretta, ma hanno bisogno anche della resistenza
alla compressione impartita dall’acqua, oppure dalla lignina nei tessuti secondari. Le forze di
tensione e di compressione nel contrapporsi portano a realizzare una struttura “precompressa”
utilizzata dalla natura fin da quando ebbe origine la vita delle piante, ma scoperta soltanto
recentemente dagli ingegneri con il calcestruzzo precompresso.
Il legno è un esempio di struttura naturale precompressa. Le cellule destinate a divenire
legno, durante le distensione dovuta alla pressione di turgore, ricevono un graduale deposito di
lignina (resistente alla compressione) che s’interpone tra le fibrille stirate di cellulosa. Poi queste
cellule muoiono lasciando una parete cellulare costituita da fibrille di cellulosa stirate e da
lignina compressa. Questo è il legno, un materiale con caratteristiche strutturali insuperabili ed
ancora oggi insostituibile in molti campi delle attività dell’uomo. Il legno è il materiale organico
più diffuso sul nostro pianeta, possiede caratteristiche di resistenza molto apprezzate e richiede
la minore quantità di energia per la sua sintesi. La cellulosa, che da un punto di vista
termodinamico è molto più ricca del legno, richiede per la sua sintesi solo poca energia in più di
quella necessaria per la sintesi del glucosio e molto meno di quella necessaria per sintetizzare
proteine e lipidi.
MEMBRANE CELLULARI
Le membrane delle cellule vegetali sono la membrana plasmatica o membrana cellulare o
plasmalemma che racchiude la cellula e si trova all’interno della parete cellulare, il tonoplasto
che delimita il vacuolo, il sistema delle endomembrane, costituente una complessa rete fra molte
membrane cellulari in rapporto tra loro ma non con le membrane interne dei plastidi, dei
mitocondri, dei microcorpi e di altri organelli cellulari.
Le membrane cellulari, a prescindere del punto in cui sono posizionate, sono
costituzionalmente molto simili ed hanno uno spessore che va da 7 nm fino ad oltre 10 nm.
Quasi tutte le membrane della cellula vegetale sono strutturalmente e chimicamente
analoghe a quelle della cellula animale, anche se le specificità delle membrane vegetali sono tra
loro molto diverse. Così, per esempio, le membrane specifiche della fotosintesi, come quelle dei
tilacoidi, oppure quelle dei perossisomi o dei gliossisomi sono assimilabili a modelli molto
diversi rispetto a quelli normalmente studiati per la membrana plasmatica e le altre membrane
interne.
Le membrane sono costituite principalmente da proteine (per la metà fino a due terzi del
peso secco totale) e lipidi. Proteine e lipidi sono in rapporto quantitativo ed hanno struttura
chimica variabili in relazione alla membrana ed allo stato fisiologico delle cellule. In particolare,
sono evidenti alcune differenze tra le membrane del plasmalemma, del tonoplasto, del reticolo
endoplasmatico, dei dictiosomi, dei cloroplasti, dei mitocondri, dei microcorpi (microsomi o
perossisomi e gliossisomi). Relativamente ad una certa membrana cellulare, ad esempio il
plasmalemma, esistono differenze nella composizione per le diverse specie vegetali.
Lo studio strutturale del plasmalemma è stato affrontato seguendo tre grandi linee: la
morfologia, la biochimica e la fisiologia. Con differenti metodologie fu possibile ipotizzare una
serie di modelli strutturali e funzionali.
- 62 -
Fiume Francesco
Fisiologia generale delle piante da orto
Irving Langmuir dimostrò che, ponendo dei fosfolipidi su di una superficie acquosa, questi
si orientano disponendo la loro testa idrofilica verso l’acqua e le loro code idrorepellenti verso
l’aria e costituiscono un monostrato (monolayer). Egli riuscì anche a misurare l’area occupata
dalle molecole fosfolipidiche mettendo a punto un semplice strumento che prese il nome di
bilancia di Langmuir.
Gorter e Grendel nel 1925, servendosi di questa tecnica e lavorando sui globuli rossi del
sangue, suggerirono l’ipotesi che i fosfolipidi costituenti la membrana cellulare devono essere
organizzati non a formare un monolayer, ma, molto probabilmente, un bilayer, cioè un doppio
strato.
Si riuscì a descrivere, con l’impiego del microscopio elettronico, l’esistenza di una
membrana cellulare di spessore di 7 fino a 10 nm, un valore molto al di sotto del potere
risolutivo del microscopio ottico, ma ricadente perfettamente nelle capacità esplorative della
microscopia elettronica. Prima di allora non fu possibile l’osservazione diretta della membrana
cellulare mediante l’uso del microscopio ottico, anche se numerose prove ed evidenze indirette
suggerivano la possibile esistenza di un rivestimento delimitante l’ambiente cellulare interno dal
mezzo esterno, senza alcun contatto diretto per l’interposizione della parete.
Le prime osservazioni dirette al microscopio elettronico, in cellule isolate e fissate con
varie modalità, mostrano come la membrana plasmatica sia costituita da due regioni elettrodense
separate da una zona intermedia, translucida e chiara ed evidenziando una struttura a sandwich.
Queste prime osservazioni vennero confermate da ricerche realizzate attraverso la diffrazione
con raggi X che suggerivano un’organizzazione trilaminare della membrana.
Contemporaneamente, gli studi biochimici delle linee elettrodense individuarono la
presenza di strati proteici compatti, mentre la zona intermedia chiara era una matrice lipidica
eterogenea. Studi successivi confermarono l’organizzazione delle membrane cellulari in bilayers
fosfolipidici attraverso i quali avveniva il passaggio di prodotti liposolubili, mentre le sostanze
idrosolubili potevano attraversare la membrana attraverso pori acquosi. Tuttavia, le membrane
cellulari facevano registrare valori di tensione superficiale costantemente bassi, mentre se
fossero state solo dei bilayers fosfolipidici tali valori dovevano essere più alti, almeno pari a
quelli che si rilevano per le gocce d’olio.
Si suppose, allora, che lungo la membrana dovessero essere presenti delle proteine
idrosolubili provviste di carica elettrica, aventi funzione di tensioattivi (capaci quindi di ridurre
la tensione superficiale) ed in grado di stabilire interazioni elettrostatiche con le teste idrofile dei
fosfolipidi. Questo è denominato modello a sandwich dove le proteine, a forma di filamenti, si
trovavano sotto e sopra il bilayer fosfolipidico ed orientano i propri gruppi polari idrofili verso
gli strati acquosi, all’interno ed all’esterno della cellula, e quelli non polari idrofobici verso le
molecole di acidi grassi.
Nello stesso modello è prevista l’esistenza di pori acquosi che originano da una soluzione
di continuità del bilayer lipidico, la cui parete è rivestita da proteine per l’introflessione nello
stesso poro dello strato proteico esterno ed interno, con la specifica funzione di trasporto passivo
per diffusione delle sostanze solubili tra la cellula e l’ambiente esterno.
La figura 31 riporta lo schema della struttura del plasmalemma secondo il modello a
sandwich proposto da Danielli e Davson, dove le proteine filamentose impaccano i lipidi
disposti su due strati a palizzata, con le teste idrofile legate alle proteine e rivolte verso le
soluzioni acquose esterna ed interna della cellula, mentre le code idrofobe di due strati (quello
più esterno e quello più interno alla cellula) sono tra loro affacciate.
- 63 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 31 – Schema della struttura del plasmalemma, bene ordinata e tendenzialmente rigida,
secondo il modello a sandwich di Danielli e Davson, con il particolare del poro
acquoso rivestito dall’introflessione degli strati proteici verso l’esterno e l’interno
alla cellula.
Il modello a sandwich fu confermato da numerose ricerche condotte sulla birifrangenza
delle membrane, sull’analisi diffrattografica, sulla microscopia elettronica e sull’analisi
biochimica diretta.
I microscopisti elettronici supposero che questo modello fosse comune a tutte le
membrane costituenti la cellula e Robertson, in particolare, avanzò l’ipotesi che il sistema
interno delle membrane cellulari dipendesse direttamente dalla membrana plasmatica la quale
era in grado di originare, per introflessione, tutta la rete di membrane interne.
E’ questa la teoria della unit membrane che promosse una serie di ricerche sulle membrane
intracellulari che evidenziarono dettagli e differenze molto consistenti e permisero, fin dal 1960,
di accumulare critiche ed incongruenze nel modello a sandwich di Danielli e Davson e di
ridimensionare le generalizzazioni di Robertson.
Si dimostrò che le membrane cellulari potevano mostrare bassi valori di tensione
superficiale anche in assenza di proteine e che non tutte le proteine presenti sulla membrana
potevano abbassare la tensione superficiale, poiché alcune di esse hanno caratteristiche idrofobe.
Il modello a sandwich, pur essendo in perfetto accordo con le immagini che si ottengono
al microscopio elettronico, non riusciva più a soddisfare la maggior parte delle caratteristiche
- 64 -
Fiume Francesco
Fisiologia generale delle piante da orto
fisiologiche e termodinamiche del plasmalemma e delle altre membrane endocellulari. Ed anche
se gli strati esterni scuri della membrana cellulare, osservabili al microscopio elettronico,
sarebbero da attribuire alle teste polari dei fosfolipidi, molto dense agli elettroni, mentre lo strato
intermedio chiaro corrisponderebbe alle code idrofobiche dei fosfolipidi, identificabili con le
catene di carbonio degli acidi grassi, meno elettrodense, l’immagine trilaminare che fornisce il
microscopio elettronico della membrana cellulare sarebbe soltanto un artefatto dovuto alla
tecnica di allestimento dei preparati microscopici.
Gli alcoli utilizzati nella fase di disidratazione, infatti, agirebbero come denaturanti delle
proteine, srotolandole e stratificandole in corrispondenza della faccia interna ed esterna del
plasmalemma.
In relazione a stringenti evidenze sperimentali Singer e Lucy prima e Singer e Nicholson
nel 1972 proponevano un nuovo modello strutturale comune a tutti gli esseri viventi che
ammette che le membrane, indipendentemente dalla loro localizzazione nella cellula sono
apparentemente tristratificate e costituite da un bilayer fosfolipidico in cui sono immerse delle
macromolecole proteiche globulari che attraversano la membrana da parte a parte, mentre altre
proteine si legano polarmente alle teste dei fosfolipidi e si muovono in senso laterale.
Secondo questo modello la membrana non è una struttura statica, ma i lipidi e le proteine
sono in costante movimento con diffusione laterale e radiale nel piano della membrana.
La membrana assume una consistenza semifluida da cui il nome di modello a mosaico
fluido o modello trilaminare a mosaico.
La matrice della membrana plasmatica è costituita da un doppio strato di fosfolipidi con le
loro code (catene di acidi grassi) che si fronteggiano le une con le altre interagendo tra loro,
mentre le teste polari sono rivolte verso la parete cellulare (quelle di uno strato) e verso
l’ambiente intracellulare (quelle dell’altro strato).
L’interno del doppio strato della membrana è costituito dalle catene alifatiche non polari
degli acidi grassi, mentre le due superfici esterne (quella rivolta verso la parete cellulare e quella
a contatto con il citosol) possono interagire con l’ambiente acquoso perché costituite da gruppi
polari dei fosfolipidi.
Le proteine, anziché filamentose e distese in un monostrato continuo (come nel modello a
sandwich), sono globulari e si possono distinguere in estrinseche o periferiche, associate cioè
solo con le superfici del doppio strato lipidico e ad esse legate attraverso interazioni
elettrostatiche o legami idrogeno, ed intrinseche o integrali o costitutive che penetrano nello
spessore della membrana oppure l’attraversano da parte a parte (proteine trans-membrana). La
porzione della molecola proteica inclusa nello strato lipidico è idrofoba, mentre la porzione che
emerge da una parte o dall’altra del bilayer e idrofila.
Le proteine estrinseche ed intrinseche, come già accennato, danno luogo, nell’ambito del
doppio strato lipidico, ad una serie di movimenti laterali e radiali, poiché esse sono costituite da
aminoacidi polari che consentono loro di stabilire con le teste dei fosfolipidi una serie di legami
dinamici che si formano e si disfano continuamente.
Ciò determina una sorta di rotolamento continuo delle proteine estrinseche lungo le due
superfici del doppio strato ed un galleggiamento e una deriva delle proteine integrali e delle
proteine trans-membrana (come se fossero dei grossi iceberg) in un oceano di fosfolipidi.
La figura 32 cerca di interpretare graficamente il modello a mosaico fluido di Singer e
Nicholson.
- 65 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 32 – Schema del modello a mosaico fluido di Singer e Nicholson.
Fosfolipidi di membrana
Molto variabile è la quantità e la natura dei lipidi delle membrane cellulari, anche se è
possibile individuare alcune caratteristiche comuni alle molecole lipidiche di tutte le membrane
biologiche.
I lipidi delle membrane di cellule procarioti sono rappresentati quasi esclusivamente da
fosfolipidi e glicolipidi, mentre nelle cellule eucarioti si trovano, oltre quelli ora citati, anche
sfingolipidi e colesterolo.
Tuttavia, sulla composizione lipidica della membrana incide anche la capacità metabolica
propria della cellula. Questo fatto non è di secondaria importanza, perché una cellula può o
meno acilare gli acidi grassi che gli pervengono, oppure, al contrario, può modificarli ed
indirizzarli verso una catena metabolica funzionante all'interno della stessa cellula. Ancora,
potrà esterificare nei fosfolipidi alcuni acidi grassi dopo il loro allungamento a VLC-PUFA, o
esterificarli dopo la retroconversione a molecole più corte.
I principali fosfolipidi del plasmalemma sono la fosfatidilserina, la fosfatidiletanolamina,
la fosfatidilcolina e la sfingomielina. Fatta eccezione per la sfingomielina, questi fosfolipidi
hanno il glicerolo (indicato in rosso) come molecola base che esterifica due dei suoi gruppi
alcolici con due molecole di acidi grassi (nello schema, uno saturo ed uno insaturo con un
doppio legame), rispettivamente, mentre il terzo gruppo alcolico è occupato da un gruppo polare
- 66 -
Fiume Francesco
Fisiologia generale delle piante da orto
costituito dalla serina (rappresentata in verde), o dalla etanolamina (in arancione), o dalla colina
(in blu):
fosfatidilserina
fosfatidiletanolamina
- 67 -
fosfatidilcolina
Fiume Francesco
Fisiologia generale delle piante da orto
Come si può osservare nello schema, le due catene di acidi grassi partono dal glicerolo e,
in virtù del fatto che sono apolari ed idrofobe, si proiettano in direzione opposta a quella del
gruppo polare. Queste molecole provviste di una testa polare e di un’estremità opposta apolare
sono dette anfipatiche. Osservando nello schema la direzione delle due code di un fosfolipide si
nota un’assenza di collinearità tra le due catene di acido grasso (l’acido grasso insaturo con un
doppio legame devia leggermente a destra) e ciò è dovuto al fatto che in presenza di doppi
legami la catena alifatica presenta ripiegamenti.
Altri lipidi del plasmalemma sono i glicolipidi ed il colesterolo, anch’essi anfipatici poiché
una parte della loro molecola (testa) interagisce con i solventi acquosi, mentre la restante parte
(coda) mostra affinità con i solventi organici.
Ciò spiega il motivo della grande rappresentatività quantitativa dei fosfolipidi nella
membrana cellulare e la scarsa presenza di trigliceridi, di colesterolo e la completa assenza degli
steroidi. Il colesterolo, in particolare, fino al 1958, per la sua scarsità, non era stato mai trovato
nelle piante per cui si riteneva che esso appartenesse soltanto al mondo animale; in quell’anno
esso fu ritrovato nelle alghe rosse e negli anni sessanta fu scoperto nella patata e poi in altre
specie, ma sempre in piccola quantità, rispetto, ad esempio, ai fosfolipidi.
Il comportamento del colesterolo nella membrana plasmatica è spiegato nella figura 33
dove si può osservare che il gruppo polare OH si dispone nella regione dove sono presenti le
teste polari dei fosfolipidie con il nucleo ciclopentanoperidrofenantrenico nella regione occupata
dalle code non polari degli acidi grassi.
Fig. 33 – Struttura del colesterolo e posizionamento della sua molecola, scarsamente presente
nelle cellule vegetali, fra i fosfolipidi della membrana cellulare.
- 68 -
Fiume Francesco
Fisiologia generale delle piante da orto
Nella stessa figura è possibile notare il ripiegamento verso destra della catena alifatica
dell’acido grasso insaturo (provvisto di doppi legami) in corrispondenza del quale si costituisce
una regione più fluida dove si posiziona la catena alifatica del colesterolo ed i tratti meno
organizzati delle code fosfolipidiche caratterizzate dalla presenza di doppi legami.
Quando le molecole anfipatiche (fosfolipidi, colesterolo ed altre) si trovano a contatto con
soluzioni acquose (ad esempio il plasmalemma che, all’interno ed all’esterno della cellula, è a
contatto con acqua) esse si dispongono in doppio strato, cioè in uno strato bimolecolare o
bilayer dello spessore di circa 7 nm.
Tutte le code non polari, aventi qualità idrofobe poiché incapaci di costituire legami
idrogeno con l’acqua, si dispongono all’interno del doppio strato, a stretto contatto le une con le
altre, senza stabilire contatti con l’acqua ed interagendo tra loro.
Le teste polari si affacciano su entrambi i lati della membrana e, per le loro caratteristiche
idrofile, sono a contatto con le soluzioni acquose con cui interagiscono formando legami
idrogeno con l’acqua.
Il bilayer fosfolipidico presenta caratteristiche dinamiche correlate alla fluidità (quindi alla
viscosità) che dipende in primo luogo dalla natura dei lipidi e dal tipo della testa polare:
fosfolipidi costituiti da una coda di acidi grassi insaturi e da una testa formata da fosfatidilcolina
induce aumento della fluidità e diminuzione della viscosità e quindi un incremento di moto dei
componenti della membrana; al contrario, fosfolipidi costituiti da una coda di acidi grassi saturi
e da una testa formata da fosfatidiletanolammina induce diminuzione della fluidità ed aumento
della viscosità e, pertanto, un decremento di moto dei componenti della membrana.
Va anche specificato che le variazioni di moto riguardano in particolare le porzioni
terminali della molecola che modificano così la loro flessibilità e pertanto sono in grado di
ripiegarsi e di interagire con le molecole vicine, mentre le teste polari ed il primo tratto della
catena alifatica sono abbastanza rigidi e praticamente immobili.
Non soltanto la natura dei componenti chimici dei fosfolipidi agiscono sulle caratteristiche
dinamiche del doppio strato fosfolipidico, ma anche la temperatura che maggiormente ne
influenza la fluidità ed inversamente la viscosità.
Temperature abbastanza elevate che superano i 35 °C inducono fluidità e movimento
all’intero bistrato, mentre il raffreddamento della membrana produce diminuzione della fluidità
fino alla solidificazione del doppio strato nel suo complesso.
La temperatura alla quale si verifica il cambiamento strutturale, dinamico, di fluidità o
viscosità della membrana si chiama temperatura di transizione.
La temperatura delle membrane cellulari, la composizione delle catene dei vari acidi grassi
ed il grado di saturazione degli stessi possono essere influenzati da fattori esogeni ed ambientali
nel giro di poco tempo e quando questi fattori si discostano notevolmente e per un certo periodo
dai livelli di normalità diventa possibile che si abbiano riflessi importanti sulla fisiologia del
plasmalemma e delle altre membrane e possano originarsi delle membranopatie che favoriscono
l’insorgenza di fenomeni di patologia della cellula, dei tessuti e dell’intera pianta.
L’analisi del comportamento fisico-chimico dei fosfolipidi in presenza di acqua permette
di comprendere le proprietà della membrana cellulare conferite dal bilayer fosfolipidico.
Un monostrato di fosfolipidi di dispone, in un ambiente acquoso, con le teste idrofile a
contatto con l’acqua e con le code idrofobe in posizione opposta e a contatto con l’aria, come
nello schema seguente:
- 69 -
Fiume Francesco
Fisiologia generale delle piante da orto
Un bistrato di fosfolipidi si posiziona sempre con le teste idrofile a contatto con l’acqua e
con le code idrofobe in maniera tale che queste rimangono solo a contatto con l’aria, come nello
schema seguente:
Se una goccia di fosfolipidi viene sospesa in acqua e la sospensione viene sottoposta ad un
campo di microonde generate da ultrasuoni (sonicazione) si ottiene la frammentazione della
goccia fosfolipidica in tante piccole vescicole che vanno a costituire una fase perfettamente
separata dall’acqua.
Lo stesso effetto si ottiene se i fosfolipidi sono posti in un solvente organico che viene
fatto evaporare lentamente. Sulla parete del contenitore si forma un monostrato lipidico che,
fatto interagire con l’acqua, si frammenta in piccole vescicole che si disperdono nel mezzo per
costituire una sospensione in cui i lipidi sono separati dall’acqua.
Ogni vescicola sospesa nell’acqua si chiama liposoma, è costituita da circa 5.000.000/μm2
di molecole fosfolipidiche, ha un diametro compreso tra 20 nm e 1 μm e può essere uni o
plurilamellare (figura 34).
- 70 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 34 – Sezione e struttura tridimensionale di un liposoma sferico. All’esterno ed all’interno
della vescicola sono esposte le teste idrofile dei fosfolipidi, mentre nel suo spessore
sono affacciate le une alle altre le code idrofobe.
I liposomi hanno una struttura molto simile a quella del plasmalemma cellulare, sono
bistratificati e nel loro interno racchiudono acqua, per cui le teste polari idrofile rivestono la
superficie esterna e quella della cavità interna, ambedue in contatto con l’ambiente acquoso; le
code alifatiche apolari, idrofobe dello strato fosfolipidico interno sono contro quelle dello strato
fosfolipidico esterno e, pertanto, protette da ogni contatto con l’acqua.
I liposomi sono costituiti da uno o più tipi di fosfolipidi, possono fondersi tra loro
scambiando i soluti dell’acqua in essi contenuta ed anche fondersi con le membrane cellulari.
Questa caratteristica ha permesso di usare i liposomi come vettori per l’introduzione nelle
cellule, in coltura ed in vivo, di differenti sostanze, senza che esse subissero modificazioni
chimiche per idrolisi parziale o totale, come farmaci, proteine enzimatiche e nucleotidi.
I liposomi permettono di studiare le proprietà delle membrane biologiche e tramite la
spettroscopia di risonanza di spin elettronico (ESR) è possibile rilevare caratteristiche dei
fosfolipidi costitutivi, con particolare riferimento al loro movimento.
In condizioni di buona fluidità del liposoma, a temperature che superano i 30 °C, si è
rilevato che le molecole fosfolipidiche di uno stesso strato scambiano facilmente e rapidamente
la loro posizione, tanto che in un secondo possono effettuare 107 scambi, diffondendo da
un’estremità all’altra di un liposoma di dimensioni simili a quelle di un organello cellulare.
Se il movimento dei fosfolipidi appartenenti allo stesso strato è una regola che conferisce
grande dinamicità alla struttura, lo scambio (denominato flip-flop) tra uno strato e l’altro di un
liposoma bilamellare è abbastanza raro, trascurabile da un punto di vista biologico e quando
avviene si verifica in modo assai lento.
Questo tipo di movimento è termodinamicamente sfavorito poiché il gruppo di testa
apolare deve attraversare una catena apolare nel cuore del doppio strato.
Inoltre, la rarità dell’evento e la bassa velocità di scambio flip-flop dei componenti da un
lato all’altro del bilayer genera un’asimmetria dei due strati fosfolipidici delle membrane ed una
differente loro composizione chimica che si mantiene nel tempo.
- 71 -
Fiume Francesco
Fisiologia generale delle piante da orto
I liposomi rappresentano un modello perfetto per lo studio della fluidità delle membrane
cellulari.
La modificazione della temperatura dei liposomi formati da un solo tipo di fosfolipidi o
anche da miscele di essi ha dimostrato che più è lunga la catena alifatica costituente la coda
apolare, tanto più bassa è la fluidità dei liposomi (e per analogia delle membrane cellulari) e
tanto più elevata è la tendenza dei fosfolipidi a formare strutture paracristalline per le interazioni
idrofobiche tra le catene di acidi grassi, quando la temperatura diminuisce.
Inoltre, minore è la lunghezza della catena di acidi grassi e maggiore è il numero dei doppi
legami insaturi contenuti (nei punti dove si trova il doppio legame la catena non è lineare, ma si
piega a gomito), tanto più elevata è la fluidità del bilayer fosfolipidico.
La cristallizzazione delle catene alifatiche di acidi grassi costituenti le code apolari dei
fosfolipidi, correlata alla diminuzione di fluidità, a seguito della diminuzione della temperatura,
è un evento sicuramente drammatico per le membrane della cellula e per la sopravvivenza
dell’organismo vegetale.
Per fortuna nelle membrane cellulari esistono concentrazioni elevate di colesterolo che
insieme all’alta percentuale di acidi grassi insaturi rendono difficile la cristallizazione delle
catene di acidi grassi delle code fosfolipidiche ed assicurano la necessaria fluidità anche a bassa
temperatura.
Va tuttavia chiarito che la membrana cellulare non deve essere eccessivamente fluida,
poiché deve possedere una sufficiente stabilità.
Gli acidi grassi saturi sono in grado di mantenere entro i limiti ottimali la fluidità della
membrana senza aumentarla troppo, mentre il colesterolo conferisce un certo grado di stabilità.
Una quota cospicua di colesterolo, in particolare, si ripartisce a livello dei due versanti
della membrana cellulare conferendone stabilità, pur diminuendone la fluidità.
Un acido grasso saturo può impacchettarsi strettamente con altre catene sature e la
struttura risultante è talmente compatta da assumere una configurazione spaziale a forma di
reticolo cristallino che induce alla membrana scarsa fluidità.
La sola presenza di un doppio legame, nella catena alifatica dell’acido grasso, determina
un ripiegamento a gomito della stessa catena, con la conseguenza che lo spazio occupato è
maggiore e la fluidità migliora, poiché uno stretto impacchettamento tra le catene carboniose
degli acidi grassi non riesce a costituirsi.
La figura 35 riporta i modelli spaziali (forme anioniche) di un acido grasso saturo (acido
stearico), monoinsaturo (acido oleico) e diinsaturo (acido linoleico).
In ogni caso, non esiste una relazione lineare tra il numero dei doppi legami presenti nelle
catene alifatiche e l’incremento della fluidità, poiché quando si costituiscono più di due o tre
doppi legami la molecola dell’acido grasso assume una disposizione spaziale ad elica e ciò
determina un impacchettamento più stretto, anche in relazione all’accorciamento della catena
alifatica determinato per l’appunto dalla disposizione elicoidale.
La disposizione spaziale del doppio legame influenza il grado di fluidità della membrana
in quanto capace di determinare una collocazione diversa dei due isomeri “cis“ e “trans“ nel
contesto della stessa struttura.
Gli isomeri “trans” degli acidi grassi monoinsaturi hanno una configurazione molto simile
a quella estesa degli acidi grassi saturi (figura 36), occupando circa lo stesso spazio e
determinando, come questi ultimi, una riduzione della fluidità della membrana. Tuttavia,
l’afflusso di acidi grassi trans-monoinsaturi al doppio strato lipidico non determina sensibili
- 72 -
Fiume Francesco
Fisiologia generale delle piante da orto
cambiamenti di fluidità della membrana, poiché questi isomeri vanno a sostituire un acido
grasso nella posizione 1 del glicerolo, piuttosto che occupare la posizione 2 dove abitualmente si
collocano gli isomeri cis-monoinsaturi (figura 37).
Fig. 35 – Modelli spaziali di un acido grasso saturo, monoinsaturo e diinsaturo. Osservando i tre
modelli si comprende come l’impacchettamento di più molecole di acido stearico
possa avvenire in maniera molto stretta (per la rettilinearità della catena alifatica)
rispetto a quello dell’acido oleico ed ancor meno dell’acido linoleico che presentano,
ciascuno, uno e due ripiegamenti. Si può ancora notare che, in senso lineare, la
lunghezza della catena alifatica diminuisce all’aumentare dei legami insaturi per la
presenza di un ripiegamento in corrispondenza di ognuno di tali legami.
- 73 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 36 – Nella geometria dei doppi legami degli acidi grassi si osserva che la configurazione
della catena satura è molto più somigliante alla configurazione della catena con
doppio legame “trans” che non a quella della catena con doppio legame “cis”.
Dall’osservazione si può dedurre come l’impacchettamento di lipidi ad acidi grassi a
catena satura ed a catena con doppio legame trans può avvenire con un simile grado
di compattezza (che induce poi nella membrana cellulare lo stesso grado di fluidità),
mentre l’impacchettamento di lipidi di acidi grassi con doppio legame “cis” deve
essere forzatamente meno stretto, poiché le catene alifatiche occupano più spazio nel
senso trasversale.
Lo scambio di acili è il mezzo più efficace per mantenere un certo grado di fluidità a
livello della membrana cellulare. Infatti, lo scambio tra coppie di acidi grassi delle code apolari
dei fosfolipidi, quali il 18:0/18:0 ed il 18:1/18:1 determina la formazione di due nuove coppie
18:0/18:1 che producono un certo grado di fluidità di membrana (figura 37).
La stabilità della membrana viene assicurata anche da molte proteine integrali le quali,
formando con i fosfolipidi dei complessi lipoproteici, immobilizzano le molecole lipidiche
interessate. La proteine, come la citocromossidasi, capaci di rivestirsi completamente di
fosfolipidi e rendendoli immobili, sono dette molecole proteiche penetranti e le molecole
lipidiche immobili che rivestono queste proteine costituiscono i lipidi di delimitazione (figura
38).
La quantità totale di fosfolipidi che vanno a costituire i lipidi di delimitazione è
proporzionale al numero di molecole proteiche penetranti che possiede la membrana. Bisogna,
tuttavia, sottolineare che, in relazione alla fluidità della membrana, i complessi lipoproteici,
formati dalle proteine penetranti e dai lipidi di delimitazione, possono subire movimenti di
traslazione. Tali movimenti possono realizzarsi unicamente nel piano della membrana, cioè
nell’ambito del proprio strato di appartenenza del bilayer, come dimostra la stabilità della
configurazione asimmetrica della membrana. Una proteina intrinseca, infatti, per le
caratteristiche idrofile delle proprie estremità, non possiede sufficiente energia per attraversare
lo spessore idrofobo del doppio strato e raggiungere l’altro lato della membrana, che per questo
- 74 -
Fiume Francesco
Fisiologia generale delle piante da orto
motivo può considerarsi come un fluido soltanto per due dimensioni dove le proteine si trovano
soltanto in soluzione parziale.
Fig. 37 – Scambio di acili tra coppie di acidi grassi delle code apolari dei fosfolipidi. A sinistra,
il fosfolipide possiede code apolari i cui acidi grassi sono ambedue acido stearico
(saturo), che conferiscono alla membrana scarsissima fluidità; al centro, il fosfolipide
possiede code apolari i cui acidi grassi sono ambedue acido oleico (monoinsaturo),
che impartisce alla membrana alta fluidità; a destra i due fosfolipidi hanno dato luogo
ad uno scambio di acili, costituendo un fosfolipide con una coda di acido stearico
(18:0) e l’altra coda di acido oleico (18:1), impartendo alla membrana caratteristiche
intermedie di fluidità rispetto agli altri due.
- 75 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 38 – Nel modello a mosaico fluido delle membrane cellulari esistono dei complessi
lipoproteici costituiti da una particolare classe di lipidi che si stratificano (lipidi di
delimitazione, indicati in rosso) sulla superficie di molte proteine integrali (molecole
proteiche penetranti).
La presenza delle proteine nella membrana cellulare è dimostrata dalle proprie
caratteristiche di impedenza elettrica che sono molto diverse da quelle dei liposomi che sono
sprovviste di molecole proteiche. I valori di impedenza elettrica di una membrana cellulare sono
di circa 1.000 ohm/cm2 e questo dato può ottenersi soltanto per i liposomi in cui siano presenti,
oltre i fosfolipidi, anche le proteine.
Proteine di membrana
Le proteine di membrana appartengono a diverse categorie, sono molto eterogenee
riguardo al peso molecolare, alla composizione aminoacidica ed alla mobilità elettroforetica e la
maggior parte di esse è costituita da proteine semplici oppure da associazioni a carboidrati
(glicoproteine) e contribuiscono all’integrità ed alla stabilità della membrana cellulare. I metodi
per la determinazione delle proteine di membrana si fondano sull’estrazione selettiva e sulla loro
caratterizzazione elettroforetica. Le proteine che contraggono rapporti idrofobici con i
fosfolipidi sono la frazione più elevata di tutte le proteine di membrana e possono essere estratte
solo dopo la dissoluzione del doppio strato con detergenti o solventi organici; sono queste le
proteine integrali. Le proteine periferiche, che sono legate in modo polare alla superficie del
doppio strato lipidico e rappresentano una quota piuttosto ridotta, possono essere rimosse dalla
membrana mediante soluzioni saline con metodi basati sull’aumento della forza ionica.
Il triton X-100, tra quelli non ionici, e SDS (sodio dodecilsolfato), tra quelli ionici, sono i
detergenti più usati a caldo per la rimozione delle proteine di membrana. Queste possono essere
sottoposte ad elettroforesi, in un gel di poliacrilamide contenente SDS, per essere separate in
base al loro peso molecolare ed essere riconosciute.
- 76 -
Fiume Francesco
Fisiologia generale delle piante da orto
Le proteine di membrana possono essere estrinseche o periferiche, associate solo alla
superficie (extracellulare o intracellulare) della membrana, e proteine intrinseche, integrali o
costitutive che penetrano profondamente nel doppio strato lipidico o anche l’attraversano.
Le proteine estrinseche aderiscono debolmente alla membrana, legandosi alle sue
estremità polari dei lipidi mediante interazioni idrofile o prendendo contatto con le proteine
intrinseche e possono essere facilmente rimosse cambiando semplicemente la forza ionica del
mezzo o aggiungendo agenti chelanti come EDTA (etilendiamminotetracetato).
Le proteine intrinseche sono globulari ed anfipatiche, poiché hanno un’estremità idrofoba
ed una idrofila, come le molecole del bistrato lipidico, ma, rispetto ad esse, sono di gran lunga di
dimensioni maggiori. L’estremità idrofoba della proteina è a stretto contatto con le code
idrofobe dei lipidi, vale a dire che la parte idrofoba di queste proteine reagisce estesamente con
la catena carboniosa della coda dei fosfolipidi, così che non può venire a contatto con le
soluzioni acquose, mentre l’estremità idrofila è allineata con le teste idrofile lipidiche o si
protende al di fuori per immergersi nel liquido circondante la membrana. Alcune di queste
proteine sono tanto grandi da oltrepassare lo spessore della membrana, solo da una parte o da
ambedue. Nel primo caso esse hanno un tratto idrofobo (quello incluso nello spessore della
membrana) ed una parte idrofoba (quello che si proietta al di fuori dello spessore della
membrana), mentre nel secondo caso esse hanno un solo tratto idrofobo (contenuto nello
spessore del doppio strato lipidico ) e due estremità idrofile sporgenti. Sono queste le proteine
penetranti e trans-membrana (figura 39). Gli aminoacidi idrofili ed idrofobi di queste proteine
hanno caratteristiche che consentono un elevato grado di interazione con i fosfolipidi di
membrana e nelle spanner proteins essi sono caratterizzati da una sequenza molto distesa che
attraversa diverse volte il bistrato lipidico della membrana (figura 39).
Fig. 39 – Proteine di membrana. A: interazioni degli aminoacidi delle proteine di membrana con
i fosfolipidi del doppio strato; B: una spanner protein (in verde) caratterizzata da una
sequenza aminoacidica attraversante diverse volte il bistrato lipidico, proteina
intrinseca (in blu) e proteina estrinseca (in azzurro). La sezione delle proteine
globulari (penetranti e trans-membrana) mostra la distinzione tra la zona idrofila
(dello stesso colore della proteina) e la zona idrofoba (in rosso), contenuta nello
spessore del doppio strato e provvista di raggruppamenti idrofobici che permettono la
stretta interazione con le code fosfolipidiche.
- 77 -
Fiume Francesco
Fisiologia generale delle piante da orto
Da un punto di vista meccanico, queste particolarità fanno della membrana cellulare una
struttura flessibile e molto resistente paragonabile ad una tela costituita da un doppio tessuto e
dove ogni tanto il filo di cotone produce un’imbastitura (la sequenza di aminoacidi ed in
particolare quella delle spanner proteins) che induce in quelle aree una maggiore resistenza a
qualunque tipo di lacerazione.
Mentre le proteine estrinseche non possiedono gruppi idrofobi e sono completamente a
contatto con le soluzioni acquose del mezzo intra o extracellulare, le proteine intrinseche
praticano due ambienti differenti.
Per la parte che sporge dallo spessore della membrana esse sono esposte all’acqua,
all’interno o all’esterno della cellula, per la restante parte esse sono immerse in una sospensione
oleosa.
Pertanto, per mantenere la stabilità in questo duplice ambiente, la proteina estrinseca deve
obbligatoriamente possedere caratteristiche anfipatiche, alla stessa stregua dei fosfolipidi di
membrana.
Tali caratteristiche sono determinate dalle proprietà degli aminoacidi costituenti la
proteina.
Di conseguenza la porzione idrofila deve contenere principalmente aminoacidi idrofili e
cioè la lisina, l’istidina, l’arginina, l’acido aspartico, l’acido glutammico, la serina e la treonina.
La porzione idrofoba immersa nel doppio strato lipidico è costituita per la maggior parte
da aminoacidi idrofobi, con proprietà lipofile.
Va anche detto che la porzione di proteina intrinseca con caratteristiche idrofobiche ed
immersa nel doppio strato lipidico varia dinamicamente nel tempo, in relazione alle funzioni che
la cellula svolge.
In altre parole, le proteine intrinseche non sono staticamente immerse nel doppio strato
lipidico, ma la loro linea di galleggiamento muta continuamente in rapporto a quanto affondano
nel bilayer.
Questo diviene possibile poiché le proteine intrinseche sono globulari e principalmente a
struttura terziaria.
Giova ricordare che le proteine globulari, a differenza di quelle fibrose le cui catene
polipeptidiche sono disposte parallelamente a formare lunghe fibre o fogli, hanno catene
polipeptidiche singole, strettamente ed in modo compatto ripiegate in una forma sferica o
globulare chiamata struttura terziaria (figura 40).
Il grado di affondamento dinamico delle proteine intrinseche è dovuto, inoltre, al fatto che
una certa sequenza di aminoacidi permette più di una configurazione spaziale, anche se in uno
stretto intervallo di possibilità, che ogni configurazione spaziale è caratterizzata da un
determinato livello energetico ed, infine, che l’ambiente recepisce soltanto ciò che la proteina
espone e che una certa molecola proteica ha caratteristiche che sono quelle proprie dei residui
aminoacidici che, a contatto con un certo ambiente assumono un’adatta configurazione spaziale.
Sulla base di questi concetti diventa intuitivo che un polipeptide si ritroverà più o meno
immerso nel bistrato lipidico in relazione alla quantità di residui di aminoacidi idrofobici che la
configurazione spaziale acquisita gli consentirà di esporre. Una modificazione della
configurazione spaziale porterà la proteina globulare ad un differente grado di immersione.
Inoltre, ogni cambiamento della configurazione spaziale comporta un salto energetico,
generalmente verso l’alto, poiché il polipeptide tenderà ad acquisire quella configurazione a
minor dispendio di energia, e quale conseguenza di un apporto energetico le proteine intrinseche
- 78 -
Fiume Francesco
Fisiologia generale delle piante da orto
di membrana saranno capaci di variare il proprio grado di interazione con il doppio strato
lipidico e modificare la profondità di galleggiamento nello spessore del bilayer.
Fig. 40 – Schema di proteine fibrose (A) e di proteine globulari (B e C), le prime mancanti
mentre le seconde tipiche della membrana cellulare. La struttura primaria si riferisce
allo scheletro covalente della catena polipeptidica ed alla sequenza dei suoi residui
aminoacidici; la struttura secondaria indica una disposizione regolare e ricorrente in
una dimensione nello spazio della catena polipeptidica ed è evidente nelle proteine
fibrose e può esistere anche in segmenti delle catene polipeptidiche delle proteine
globulari; la struttura terziaria riguarda il modo in cui la catena polipeptidica è
ripiegata tridimensionalmente a formare la struttura compatta e strettamente avvolta
delle proteine globulari tipiche delle membrane cellulari (B); la struttura quaternaria
interessa il modo in cui le singole catene polipeptidiche di una proteina costituita da
due o più catene sono disposte l’una rispetto all’altra (C). La maggior parte delle
proteine più grandi contengono due o più catene polipeptidiche, in genere non unite
da legami covalenti.
Un esempio di quanto discusso deriva da una proteina intrinseca di grande rilevanza
biologica, la citocromossidasi, un’ossidasi terminale che con la sua attività accetta elettroni
trasferendoli all’ossigeno e terminandone il flusso nel momento in cui gli stessi elettroni sono
catturati dall’ossigeno insieme a 2 protoni (2 H+), provenienti dalla matrice mitocondriale, con
produzione di H2O. Questo enzima della membrana interna dei mitocondri, costituente
l’elemento terminale della catena di trasferimento degli elettroni implicata nella sintesi di ATP è
lungo circa 5,5 nm, largo 6 nm e profondo da 8 a 8,5 nm. La profondità è più che sufficiente per
consentire alle molecole dell’enzima di penetrare completamente nel bistrato lipidico, che ha
uno spessore di soli 4,5 nm, lasciando sporgere da una parte e dall’altra della superficie della
membrana mitocondriale, allo stesso modo, le sue estremità idrofile. In relazione alla propria
attività, questo enzima cambia la propria linea di galleggiamento rispetto al film lipidico della
membrana.
Il movimento delle proteine di membrana non avviene soltanto secondo una direzione
perpendicolare al piano del plasmalemma con la modificazione del grado di affondamento nel
doppio strato lipidico, ma anche in senso traslatorio. Le proteine possono liberamente muoversi
nel piano fosfolipidico così come è dimostrato in alcuni esperimenti realizzati con la marcatura
mediante anticorpi aventi gruppi cromofori.
- 79 -
Fiume Francesco
Fisiologia generale delle piante da orto
Le proteine di due cellule, ciascuna appartenente a specie diverse, sono state marcate con
anticorpi coniugati con flurocromi di colore diverso.
Quelle di una specie con fluorescina, verde, e quelle dell’altra specie con rodamina, rossa.
Mediante trattamento con glicole polietilenico è stato provocato la fusione delle due cellule con
la formazione di una cellula ibrida (eterocarion).
Dopo la fusione è stato possibile osservare una metà della membrana dell’eterocarion
colorata in verde ed una metà colorata in rosso; ciascuna metà corrisponde ad una delle due
cellule di cui è stata indotta la fusione e ciascuna colorazione e dovuta al gruppo cromoforo
verde legato alle proteine di membrana (tramite l’anticorpo) derivate da una cellula ed a quello
rosso legato alle proteine di membrana derivate dall’altra cellula.
Dopo alcuni minuti la distinzione tra le colorazioni è meno netta e dopo un’ora i due colori
si trovano uniformemente dispersi e fusi tra loro. Ciò indica che già dopo alcuni minuti dalla
fusione, i fluorocromi degli anticorpi e quindi le proteine ad essi legate hanno cominciato a
spostarsi, dimostrando la loro dinamicità ed il fatto che esse sono dotate di un attivo e libero
movimento nel piano fosfolipidico.
E’ stata anche misurata la velocità del rimescolamento dei colori e quindi degli anticorpi a
cui erano legate le proteine di membrana ed è stato osservato che la velocità del loro movimento
è funzione della temperatura, crescendo al suo aumento fino ad un certo limite (dopo il quale
diminuisce), e della composizione lipidica delle membrane che a sua volta ne influenza la
viscosità. Ad esempio, un aumento di viscosità dello strato fosfolipidico diminuisce la velocità
di spostamento delle proteine.
Va anche detto che se una proteina integrale è libera di spostarsi nello spessore della
membrana, questo movimento è pur sempre relativo, poiché non può esplorare l’intera area a
sua disposizione.
In primo luogo esistono scarse possibilità che possa ritornare al punto di partenza e nel suo
percorso vi sono probabilità che la proteina possa essere bloccata, aggregata ad altre molecole o
internalizzata, cioè guidata verso l’interno della cellula.
Tuttavia, l’equilibrio che si viene a determinare tra i tanti possibili destini di una molecola
proteica è tale che comunque risulta essere il maggiore determinante funzionale della membrana
cellulare.
Con molta probabilità, molti spostamenti delle proteine integrali di membrana sono
mediati da varie componenti del citoscheletro, formato dai microfilamenti e dai microtubuli
costituenti un’impalcatura che si modifica continuamente.
Molte proteine si distribuiscono nel piano della membrana in maniera ordinata e non
casuale, come pure si osserva una loro inusuale concentrazione in particolari e ristrette aree del
plasmalemma, come nel caso della endocitosi mediata da recettori, grazie alle strette
interconnessioni esistenti tra molecole proteiche di membrana e strutture del citoscheletro
cellulare.
L’endocitosi consiste nell’assunzione di materiali esterni da parte della cellula che può
avvenire con tre differenti modalità: per fagocitosi, tipico delle amebe, per pinocitosi consistente
nell’assunzione aspecifica di piccoli quantitativi di liquido extracellulare dove è contenuto
materiale più diverso, compreso macromolecole di interesse metabolico, per endocitosi mediata
da recettori.
Quest’ultima è un processo altamente specifico poiché appositi recettori disposti sulla
membrana selezionano le macromolecole del liquido extracellulare e guidano il loro ingresso
- 80 -
Fiume Francesco
Fisiologia generale delle piante da orto
nella cellula (internalizzazione) anche quando sono ad elevata diluizione o sono mescolate con
altre molecole.
I recettori sono delle proteine integrali, spesso delle glicoproteine, in grado di raccogliere
il segnale ambientale che consente la selezione di macromolecole esterne ed il contatto con le
stesse, diretto o indiretto attraverso molecole ponte, ed operare successivamente
l’internalizzazione.
Ogni recettore della membrana cellulare si lega stereochimicamente alla molecola (detta
ligando) che ha riconosciuto ed innesca movimenti di invaginazione della membrana
sottostante, che ingloba il ligando ancorato al recettore e lo racchiude in una vescicola destinata
a fondersi nel citoplasma con i lisosomi, i cui enzimi digeriscono il materiale macromolecolare
costituente il ligante.
Le fossette da cui inizia l’invaginazione della membrana sono chiamate fossette
ammantate o rivestite (coated pits) e presentano delle spinosità addossate alla membrana
plasmatica dalla parte del citoplasma.
Queste spinosità sono la porzione dei recettori di membrana che sporge dal lato
citoplasmatico della membrana stessa.
Quando le fossette ammantate si approfondano nel citoplasma e si chiudono, circondando
il materiale in esse contenute, costituiscono quelle che si denominano vescicole ammantate o
rivestite (coated vesicles).
Il materiale elettrodenso che riveste le fossette e le vescicole è di natura proteica e la
proteina più comune è la clatrina, proteina fibrosa simile a quella schematizzata nella figura
40A, con peso molecolare di 180 kD, associata ad un polipeptide minore, con peso molecolare
di 35 kD.
La clatrina con una sua estremità si lega alla porzione dei recettori di membrana
costituente la spinosità della fossetta o della vescicola ammantata, mentre con l’altra estremità si
unisce all’actina dei microfilamenti del citoscheletro superficiale, una proteina globulare di peso
molecolare di 42,3 kD, presente in tutte le cellule eucariotiche.
Questo induce che le componenti citoscheletriche cambino continuamente la propria
organizzazione - stabilendo, così delle interazioni tra proteine intrinseche della membrana e
citoscheletro cellulare che svolgono, pertanto, un ruolo cardine nella fisiologia della cellula
vegetale - e che il legame di proteine recettoriali con particolari fattori extracellulari possa
determinare modificazioni nell’organizzazione spaziale del citoscheletro, responsabili dei
cambiamenti che caratterizzano alcune fasi del ciclo e del differenziamento delle cellule
vegetali.
La clatrina forma dei polimeri, chiamati triskelion, ciascuno costituito da tre braccia
proteiche, divaricate tra di loro a 120° e tenute insieme da polipeptidi centrali.
Ogni triskelion contiene tre molecole di clatrina alternate a tre polipeptidi minori per un
peso molecolare totale di 635 kD).
I triskelion si assemblano formando strutture a paniere, a maglie esagonali e pentagonali,
che nel formarsi costringono la membrana delle fossette ammantate ad invaginarsi nel
citoplasma, chiudendosi poi per formare la vescicola ammantata (figura 41).
Nonostante i ligandi siano dispersi nella sospensione extracellulare a diluizioni molto alte,
essi riescono ad entrare in contatto ed a legarsi specificamente con i rispettivi recettori.
- 81 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 41 – Coinvolgimento delle proteine di membrana nell’endocitosi mediata da recettori. In
alto, accoppiamento ligando-recettore, con formazione della fossetta rivestita e poi
della vescicola ammantata; subito sotto, particolare schematico di costituzione dei
cestelli a maglie pentagonali ed esagonali, a partire dalle molecole di clatrina
formanti, assieme ad altri polipeptidi di minor peso molecolare, sistemi a tre bracci
(triskelion); ancora sotto, microfotografie dei triskelion e dei cestelli di clatrina; in
basso, rappresentazione schematica delle fasi dell’endocitosi fino alla restituzione dei
recettori alla membrana plasmatica (riciclaggio dei recettori).
- 82 -
Fiume Francesco
Fisiologia generale delle piante da orto
Ciò induce la polimerizzazione della clatrina – che si organizza per formare dei panieri o
cestelli, costituenti ciascuno la vescicola ammantata nel cui lume, legati ai recettori di
membrana, si trovano i ligandi – e la successiva internalizzazione delle stesse vescicole.
Queste si spostano nel citoplasma, si fondono tra loro formando gli endosomi (vescicole
più grandi), i quali perdono la clatrina che ritorna in superficie al plasmalemma per essere
nuovamente utilizzata.
Successivamente, l’endosoma si fonde poi con una vescicola disaccoppiante detta CURL
(acronimo di Compartment of Uncoupling of Receptor and Ligant) la quale è caratterizzata da
un pH uguale a 5. La fusione di CURL con l’endosoma favorisce la separazione tra il recettore
ed il ligando. I recettori sono ormai liberi e possono concentrarsi in una ristretta zona della
nuova grande ed allungata vescicola endosoma-CURL. Questa zona è un’area di membrana che
si stacca completamente dall’endosoma-CURL formando una vescicola carica di recettori che va
al plasmalemma riportandovi le proteine integrali con funzione recettoriale. Si verifica quello
che si chiama riciclaggio dei recettori, per cui le molecole proteiche recettrici non debbono
essere nuovamente sintetizzate ad ogni ciclo di endocitosi, non vi sono problemi di accumulo di
residui e smaltimento di proteine e si realizza il ripristino di quella parte di membrana cellulare
andata perduta con l’internalizzazione delle vescicole ammantate. La restante porzione di
endosoma contenente il ligando resta nel citoplasma, si fonde con i lisosomi e va a costituire un
lisosoma secondario nel quale si verifica l’idrolisi dello stesso ligando per opera di enzimi litici.
Nella figura 41 è riportato uno schema che sintetizza le varie fasi dell’endocitosi mediata
da recettori: accoppiamento ligando-recettore, formazione della fossetta rivestita e
polimerizzazione delle molecole di clatrina assieme ad altri polipeptidi per costituire un cestello,
internalizzazione del ligando sotto forma di vescicola ammantata, fusione delle vescicole e
produzione dell’endosoma, fusione di questo con la vescicola CURL, disaccoppiamento
ligando-recettore e formazione di una vescicola contenente i soli recettori e di una vescicola con
le molecole di ligando, fusione di quest’ultima con un lisosoma primario e produzione di un
lisosoma secondario contenente il ligando poi idrolizzato da enzimi litici dello stesso lisosoma,
trasferimento della vescicola con recettori al plasmalemma (riciclaggio dei recettori).
Attraverso l’endocitosi la cellula può prelevare differenti materiali dall’ambiente esterno
attraverso le invaginazioni della membrana plasmatica le quali, come descritto in modo
particolareggiato, si staccano e si muovono con il loro contenuto nel citoplasma.
Nei batteri, nelle vere muffe mucillaginose e nelle muffe mucillaginose cellulari spesso il
materiale trasferito dall’esterno della cellula al suo interno è solido, in tal caso questa particolare
endocitosi è chiamata fagocitosi, dal termine greco phagein che significa mangiare. La
fagocitosi è innescata dal contatto fra la membrana cellulare che esercita l’attività fagocitaria e
la superficie di grandi aggregati.
L’assunzione aspecifica di particelle diluite in un liquido è chiamata pinocitosi, dal greco
pinein che vuol dire bere.
Essa avviene non soltanto in organismi unicellulari, ma anche in piante pluricellulari ed è
distinta in macropinocitosi e micropinocitosi, in base al diametro delle gocciole di liquido
penetrate nella cellula ed all’entità delle modificazioni manifestate dalla membrana cellulare.
Nella macropinocitosi, visibile anche al microscopio ottico, la cellula incamera gocciole con
diametro fino ad un minimo di 0,2 μm, mediante il sollevamento di pliche e la successiva
invaginazione della membrana sottostante alle gocciole.
- 83 -
Fiume Francesco
Fisiologia generale delle piante da orto
Il processo che, come la fagocitosi coinvolge vari elementi del citoscheletro, richiede
energia rilasciata dall’idrolisi di ATP.
La micropinocitosi, evidenziabile soltanto al microscopio elettronico, poiché le gocciole
assunte hanno diametri medi di 65 nm, comporta la penetrazione del liquido nella cellula
attraverso aperture tubulari dette caveole, al cui termine si forma la vescicola pinocitotica che
internalizza il liquido ed il suo contenuto.
Molte attività di pinocitosi non sono altro che endocitosi mediata da recettori, poiché
richiedono la presenza di recettori di membrana che selezionano il materiale da assumere.
L’endocitosi può anche procedere in senso opposto a quello descritto.
Molte sostanze vengono espulse dalla cellula per trasferimento dall’interno della cellula
verso il plasmalemma e fusione con quest’ultimo delle vescicole o di speciali vacuoli che le
contengono.
Un esempio di questo processo è il ruolo delle vescicole del Golgi nella formazione della
parete cellulare, di cui si è detto a proposito della sua genesi e del suo arricchimento (fig. 19).
Queste vescicole, contenenti precursori della parete cellulare, si spostano dall’interno verso la
superficie della cellula fino a raggiungere il plasmalemma.
La loro membrana limitante si fonde con essa ed il contenuto viene espulso nella regione
di formazione della parete. Questa endocitosi inversa è chiamata esocitosi.
Si ricorda che il processo ora descritto inizia nel reticolo endoplasmatico ruvido, dal quale
gemmano vescicole di transizione che attraversano la faccia di formazione del dittiosoma e
producono le cisterne giovani che maturando si trasformano in cisterne vecchie che a loro volta
gemmano dalla faccia di maturazione del dittiosoma verso la periferia cellulare come vescicole
secretorie.
Queste vescicole sono trasferite, su tracce microtubulari del citoscheletro, fino al
citoplasma superficiale.
Qui i microfilamenti della trama periferica che circonda la cellula costituiscono
un’importante barriera che può essere superata solo con variazioni locali dei cationi Ca2+,
innescate da segnali ormonali recepiti dalla cellula per mezzo di specifici recettori.
L’entrata di cationi calcio porta all’attivazione di alcune actin severing proteins che non
sono altro che proteine che tagliano i filamenti di actina che, agendo sui microfilamenti della
trama periferica, aprono varchi nella stessa trama e permettono alle vescicole secretorie di
raggiungere le zone attive di membrana, sedi dove le stesse vescicole possono scaricare
all’esterno del plasmalemma il proprio contenuto che va ad integrarsi nella parete cellulare che
in tal modo aumenta le dimensioni in tutte le direzioni.
Le vescicole di secrezione, contenenti i materiali da riversare all’esterno del plasmalemma
per esocitosi, si fondono prima con la stessa membrana plasmatica.
La fusione inizia con il collabimento dello strato lipidico esterno delle vescicole con
quello lipidico interno della membrana.
Si forma transitoriamente una struttura pentalaminare delle membrane in via di
coalescenza: in corrispondenza della superficie di contatto e di fusione tra la membrana della
vescicola e la membrana plasmatica, il terzo strato esterno della prima membrana (della
vescicola) si fonde con il terzo strato interno della seconda (del plasmalemma) per formare un
unico strato; questo, insieme ai due strati residui della vescicola ed ai due residui del
plasmalemma costituiscono un pentastrato.
- 84 -
Fiume Francesco
Fisiologia generale delle piante da orto
A fenomeno completato, si ricostituisce una normale membrana trilaminare a mosaico
(modello di Singer e Nicholson).
Nella figura 42 è riportato uno schema di esocitosi in cui una vescicola matura di
transizione (a) proveniente dal reticolo endoplasmatico ruvido, dove è stata sintetizzata, migra
fino alla membrana plasmatica con la quale si fonde iniziando il collabimento del proprio strato
lipidico esterno con quello interno della stessa membrana e determinando transitoriamente una
struttura pentalaminare del plasmalemma (b) in via di coalescenza (c); a fenomeno completato si
ricostituisce una normale membrana trilaminare (d) costituita da materiale fornito dalla stessa
vescicola, mentre il contenuto di quest’ultima viene versato nella regione della parete (e) ed
essere assimilato dalla stessa (f).
Fig. 42 – Esempio schematico di esocitosi in una cellula vegetale. Una vescicola matura di
transizione (a) migra dal reticolo endoplasmatico ruvido fino al plasmalemma con la
quale si fonde, iniziando il collabimento del proprio strato lipidico esterno con quello
interno della membrana cellulare e determinando transitoriamente una struttura di
membrana pentalaminare (b) in via di coalescenza (c); a fenomeno completato si
ricostituisce una normale membrana trilaminare (d), mentre il contenuto della
vescicola viene versato nella regione della parete (e) ed assimilato dalla stessa (f).
Con questo meccanismo la vescicola trasporta membrana neosintetizzata e nuovi materiali che
arricchiscono, rispettivamente, il plasmalemma e la parete cellulare.
Per questo motivo la composizione proteica delle nuove aree della membrana cellulare è diversa
da quella del preesistente plasmalemma.
Queste aree derivate dalle vescicole di secrezione sono ben presto ricoperte sulla faccia interna
da molecole di clatrina o, più verosimilmente, le vescicole mature escono dall’apparato del
Golgi già rivestite da una parete di clatrina e sono destinate a diventare le fossette rivestite di
cicli di endocitosi mediata da recettori.
In altri termini le citomembrane utilizzate per l’esocitosi sono riciclate come vescicole
endocitotiche e rinviate al reticolo endoplasmatico, secondo un ciclo descritto nella figura 43.
- 85 -
Fiume Francesco
Fisiologia generale delle piante da orto
Fig. 43 – Rapporti funzionali tra esocitosi ed endocitosi. Le membrane cellulari utilizzate per la
secrezione di materiali neosintetizzati a livello del reticolo endoplasmatico (esocitosi)
sono riciclate come vescicole ammantate (endocitosi) e rinviate all’apparato del Golgi.
La citomembrana (in rosso) costituente la fossetta esocitotica (a), derivata dalla
vescicola secretoria, viene riciclata per costituire la membrana della fossetta rivestita
(b) e poi quella della vescicola ammantata, nell’endocitosi mediata da recettori. Il
materiale della fossetta esocitotica viene assimilato dalla parete cellulare, mentre il
materiale che entra nella cellula dall’ambiente esterno si lega ai recettori di membrana
come ligando.
- 86 -