
LA GENETICA DEI
MICRORGANISMI
ovvero
Anche i batteri fanno
sesso…
(cioe’ si scambiano geni!)
La coltivazione dei batteri
Una colonia è formata da
circa 107cellule.
Clone = discendenti di
un’unica cellula.
Mappatura nei procarioti
Strategie analoghe
Incroci tra ceppi che differiscono per marcatori genetici e
identificazione e conta dei ricombinanti
Differenze
Eucarioti: scambio tra
cromosomi appaiati alla
meiosi che produce
2 prodotti reciproci
Procarioti: no meiosi,
ricombinazione non reciproca
e prodotti non reciproci
Trasferimento unidirezionale
Scambi mai tra cromosomi interi ma
tra il cromosoma principale di un ceppo (ricevente)
e frammenti di DNA dell’altro (donatore)
Marcatori genetici? Capacità di crescere
Resistenza agli antibiotici o ai virus
Mutanti
Nutrizionali (auxotrofi)
Fonte di carbonio
Forma delle colonie
Meccanismi mediante i quali avviene
scambio di materiale genetico tra batteri:
Trasformazione
Coniugazione
DNA libero
contatto cellule
Trasduzione
mediato da fagi
La trasformazione
scoperta da F. Griffith nel 1928 in Streptococcus pneumoniae.
Trasferimento di materale genetico da un batterio
all’altro mediato da frammenti di DNA extracellulare.
Nella trasformazione frammenti
isolati di DNA vengono assorbiti
dall’esterno
all’interno
della
cellula
La trasformazione a livello
molecolare
La mappa per
trasformazione
Con la
trasformazione è
possibile mappare
i geni: quanto più
due marcatori
sono vicini, tanto
più probabile è la
loro
cotrasformazione.
ordini possibili: p-q-o e p-o-q
La scoperta
della
coniugazione - 1
Nel 1946 Joshua Lederberg e
Edward Tatum dimostrarono
che due ceppi di E.coli
auxotrofi (E. coli A met- biothr+ leu+ thi+ ed E. coli B met+
bio+ thr- leu- thi-), posti a
contatto, possono scambiarsi
materiale genetico e creare
dei batteri prototrofi.
Colonie prototrofe: 1x10-7
La scoperta della coniugazione - 2
Alla fine degli anni ‘40
Bernard Davis scopre
che se i due ceppi
sono separati da un
filtro attraverso cui
possono passare
molecole (ma non batteri) non si
ha la formazione di prototrofi: il
contatto fisico tra i due ceppi è
indispensabile!
Il fattore di fertilità (F)
W. Hayes (1953): il trasferimento genico avviene in
una sola direzione: da un donatore (maschio) ad un
ricevente (femmina).
Hayes trovò un donatore “sterile” (cioè incapace di
trasferire l’informazione) che si era trasformato in
un ricevente.
Ipotesi
capacità di fungere da donatore è una condizione ereditaria
determinata da una fattore di fertilità F.
Ceppi con F sono donatori (F+), quelli senza F sono riceventi (F-).
Fattore F: plasmide che si replica nel citoplasma batterico in modo autonomo.
La scoperta della
coniugazione - 3
I ceppi Hfr
Luca Cavalli Sforza scoprì un nuovo ceppo
derivato da un F+ chiamato high frequency of
recombination (Hfr). L’incrocio HFR x F- dava
1000 volte più ricombinanti dell’incrocio F+ x
F-, ma nessuna cellula F- diventava F+.
Il ceppo Hfr si forma in seguito all’integrazione di F
nel cromosoma batterico.
Quando si mescolano cellule F+ con cellule F-:
il fattore F si integra nel cromosoma batterico con una bassa frequenza.
I geni batterici si trasferiscono ma con bassa frequenza.
Nell’incrocio Hfr x F- invece:
tutti i batteri donatori hanno F integrato, frequenza di ricombinanti è alta.
L’integrazione
di F
Ricombinazione tra due anelli di
DNA, in seguito ad un singolo
crossing over, con formazione di
un anello unico che è la somma
dei due precedenti.
In questo modo un ceppo F+ (con
F plasmide libero) diventa HFR.
Una volta inserito, il plasmide F
mantiene la sua capacità di
riconoscere un batterio F- e di
iniziare la coniugazione.
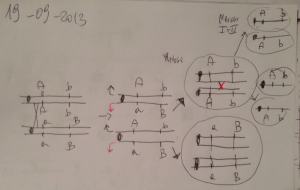
Il trasferimento di HFR - 1
Il trasferimento di F integrato (HFR) comincia sempre a partire
dall’origine di replicazione O. Quindi il ricevente, per poter
diventare F+, dovrebbe ricevere tutto il cromosoma batterico
del donatore.
O
lac
pro
thr
pur
gal
O
HfrH
pur
his
gli
thi
lac
pro
thr
il fattore F
gal
Il trasferimento
di HFR - 1
Poiché il pilo sessuale è instabile,
il trasferimento non è mai completo,
per cui il ricevente resta F- ma riceve
una parte di DNA batterico donatore
che può eventualmente essere integrata
dal ricevente.
lac
pro
thr
pur
gal
O
HfrH
hi
g
s li
thi
lac
pur
pro
thr
gal
Il trasferimento di HFR - 2
A questo punto, il ricevente non ha ricevuto un DNA circolare, ma
lineare! Questo potrà eventualmente essere integrato nel
genoma del ricevente per ricombinazione sfruttando l’omologia
del DNA batterico, e solo in seguito ad un numero pari di
crossing over.
La coniugazione interrotta
Elie Wollman e François Jacob, 1957
Si può sfruttare la labilità del
pilo e l’inserzione di F per
mappare
i
geni
di
un
cromosoma batterico.
Le inserzioni di F sono casuali
sia come punto di inserzione, sia
come orientamento, quindi ogni
ceppo batterico HFR trasferirà
geni nel ricevente con un ordine
che riflette la disposizione dei
geni lungo il cromosoma.
La procedura
I due tipi di cellule vengono
mescolate in gran quantità in
un terreno liquido a 37°C. A
vari tempi vengono prelevati
dei campioni della coltura,
vengono
agitati
(per
interrompere artificialmente
la coniugazione) e piastrati su
terreni selettivi contenenti
streptomicina per uccidere le
cellule Hfr. Quello che si
ottiene è una mappa a tempo
del
cromosoma
batterico
donatore.
Esempio: donatore HfrH thr+leu+aziRtonR
lac+gal+strS ricevente: F- thr leu aziStonS lac gal
strR
Le mappe a tempo
Wollman e Jacob scoprirono che:
Ogni allele del donatore appariva nel ricevente F- dopo un
intervallo di tempo ben preciso dall’inizio della coniugazione.
Gli alleli del donatore si presentavano sempre in una specifica
sequenza.
I marcatori che entravano più tardi comparivano in un numero
minore di cellule.
Da queste osservazioni dedussero che il trasferimento avviene a
partire da un punto ben preciso sul cromosoma del donatore chiamato
origine O e prosegue secondo modalità lineare.
Il gradiente di trasferimento
Anche se la coniugazione non viene
interrotta il ponte di coniugazione
si rompe spontaneamente. Pertanto
solo di rado il cromosoma viene
trasferito per intero.
La rottura spontanea può
verificarsi in qualsiasi momento.
Ciò determina un gradiente di
trasferimento
il marcatore più vicino all’origine
(es. met) verrà trasferito con la
frequenza più alta
Costruzione della
mappa
Come unità di misura si usa la distanza
di tempo (in minuti) impiegato dagli
alleli del donatore ad entrare nel
ricevente.
Solo dopo 2 ore i riceventi diventano F+
il fattore F entra per ultimo.
Il cromosoma batterico è circolare
Usando vari ceppi Hfr ed ottenendo mappe diverse, Campbell ipotizzò
che il cromosoma batterico fosse circolare.
Il saggio a tre punti nei batteri
La sesduzione,
ovvero F’
Nel 1959 E. Adelberg, in un ceppo
Hfr il fattore F può excidersi e
generare un ceppo F+.Se però si
excide in maniera errata può
incorporare il gene batterico
accanto al quale era inserito. In
questo caso prende il nome di F’. Il
plasmide F’ può coniugare e
inserire il gene che ha incorporato
in un batterio ricevente
(sesduzione) che diventa F+
La trasduzione
trasferimento di geni
batterici mediato da
batteriofagi
Ciclo di
crescita del
fago temperato
lambda
Trasduzione generalizzata e
trasduzione specializzata
I fagi che effettuano la trasduzione generalizzata
veicolano qualsiasi porzione del genoma dell’ospite
I fagi che effettuano la trasduzione specializzata
trasferiscono solo porzioni specifiche
La trasduzione generalizzata
(Joshua Lederberg e Norton Zinder 1951)
Salmonella typhymurium
phe+trp+tyr+ met-his-
x
phe-trp-tyr- met+his+
ricombinanti prototrofi anche se messi in un tubo ad U
vettore della ricombinazione: il fago P22
Il meccanismo della trasduzione
generalizzata è stato chiarito da
Ikeda e Tomizawa con il fago P1 nel
1965
Trasduzione
generalizzata
NB nella testa del fago non
restano geni fagici!!
Con la trasduzione generalizzata si possono
stabilire relazioni di associazione tra i geni
La cotrasduzione: il trasferimento di due
geni batterici (molto vicini) ad opera dello
stesso fago
donatore leu+thr+aziR x
thr
leu azi
thr
ricevente leu-thr-aziS
oppure
leu azi
thr
azi leu
Come si calcola la
frequenza di cotrasduzione?
donatore a+b+ x
ricevente a b
a+b
a b+
trasduttanti
a+b+
frequenza di
cotrasduzione =
numero dei trasduttanti
per entrambi i marcatori
numero di trasduttanti totali
selezionando per a+
freq. di cotrasduzione =
selezionando per b+
freq. di cotrasduzione =
(a+b+)
x 100%
(a+b)+(a+b+)
(a+b+)
x 100%
(a b+)+(a+b+)
x 100%
Trasduzione specializzata
Il profago λ si integra con un crossing over a
livello del sito di attacco di lambda tra i geni gal e
bio
Meccanismo della trasduzione specializzata
Riassunto
Mappatura del genoma fagico
Fenotipi fagici : placche di lisi e specificità
d’ospite (ceppo batterico che un fago è in
grado di lisare)
Marcatori del fago T2:
h+ lisa il ceppo B di coli ma non il ceppoB/2
h
lisa sia il ceppo B che il B/2
r+ placche piccole con margini indistinti
r placche grandi con margini netti
quando fagi h+ crescono su uno
strato misto di cellule B e B/2
formano placche torbide perchè
lisano solo B e i batteri B2 crescono
nelle placche provocando torbidità
delle placche. I fagi h formano
invece placche chiare
ceppo B
Il lisato si piastra su una
miscela di ceppi B e B/2
Frequenza di
ricombinazione
placche (h+ r+) + (h r )
tra h ed r
=
placche totali
x 100









![1 LEZIONE TFPC - genetica batterica [modalità compatibilità]](http://s1.studylibit.com/store/data/001115261_1-c0368de7ca28d13cd5cf5bbccd6e599a-300x300.png)