CONIUGAZIONE
BATTERICA
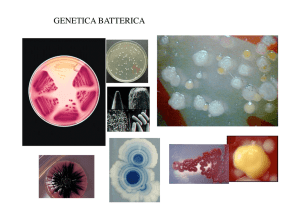
Come si coltivano batteri in laboratorio
Terreno minimo: acqua, sali
inorganici, fonte di carbonio
Batteri prototrofici:
crescono in terreno minimo
Batteri auxotrofici:
Incapaci di crescere in
terreno minimo, se non
complementato da altre
sostanze
Cosa si vede
Genoma batterico:
- Non delimitato da membrana
- Singolo cromosoma circolare
- Quasi totale assenza di sequenze non
codificanti
- Non avviene la meiosi
Le cellule figlie sono sempre uguali alle
cellule madri?
Esiste un qualche tipo di “unione sessuale”?
Lederberg e Tatum:
la scoperta della coniugazione
Evento simile alla riproduzione
sessuata
Attraverso il poro
passano solo i soluti
e non I batteri
Coniugazione richiede il contatto fisico
Pilo di coniugazione: ponte e
poro con cui le cellule si
scambiano materiale genetico
Per caso Hayes (‘53) scopre un altro tipo di passaggio
di informazione: una variante del ceppo originale che
non produce ricombinanti quando incrociata al
ricevente.
La coniugazione e’ diseguale: Una cellula e’ donatore
(di una parte del materiale genetico), l’altra (mutante)
e’ solo ricevente, esiste una sorta di asimmetria
sessuale. In contatto con ceppi wild-type, il difetto era
recuperato: Fattore di fertilita’ (F) era trasmissibile.
F e’ un plasmide, che esprime geni per la
generazione del pilo. Si duplica con replicazione
rotante, in cui trasferisce un singolo filamento e una
copia rimane in donatore
Il fattore F è un PLASMIDE
Molecola di DNA a doppia elica, circolare
Circa 100 kbp
Replica in modo autonomo dal cromosoma
- origine di replicazione “vegetativa”
Contiene geni
- per la propria replicazione
- per costruire il “pilus sessuale”
- per “coniugare”
I ceppi Hfr
Luigi Luca Cavalli Sforza scopre un ceppo F+ con
delle particolarità:
1. Incrociato con F- produce un numero di
ricombinanti 1000 volte superiore ad un F+
normale (da qui il nome High Frequency of
Recombination)
2. Negli incroci Hfr x F- nessun F- era convertito in
F+ o Hfr (diversamente dagli incroci F+ x F-)
F+: met+ x F-: met- = 1/10-7 ricombinanti (ma tutta
la popolazione diventa F+)
Hfr: met- x F-: met- = 1/10-4 ricombinanti (ma tutta
la popolazione rimane F-)
Il ceppo Hfr deriva dall’integrazione del fattore F nel
cromosoma batterico à durante la coniugazione il
fattore F promuove il trasferimento di materiale
cromosomico con alta efficienza
à trasforma il cromosoma batterico in un
maxiplasmide
I geni del donatore possono essere trasferiti in tutto o in parte nel ricevente;
questi possono essere incorporati nel ricevente mediante crossing over
(doppio) e quindi ricombinare (creazione di ricombinanti).
DIPLOIDE PARZIALE
(ricorda questo nella
genetica dell'operatore)
Perché in un incrocio F+ x F si ottengono ricombinanti per i
marcatori cromosomici?
• In una popolazione di batteri F+ può succedere che il
plasmide F si integri nel cromosoma batterico. Questi eventi
sono (quasi) casuali e rari (frequenza circa 10-6)
• Si producono così cloni indipendenti di cellule “Hfr” mescolate
alla stragrande maggioranza di cellule F+.
• Solo questa sottopopolazione di batteri “Hfr” è responsabile
del trasferimento del cromosoma negli incroci F+x F e quindi
della ricombinazione (a bassa frequenza) di marcatori
cromosomici.
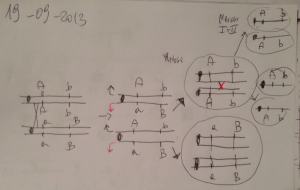
Coniugazione interrotta (da una agitazione meccanica):
gli alleli entravano nel ricevente F- secondo un ordine preciso = si
formano meno ricombinanti contenenti i geni più tardivi
Hfr azir tonr gal+ strs X F- azis tons lac- gal- str+
nella coniugazione il trasferimento del DNA a singolo filamento
inizia da un punto fisso del cromosoma: la ORIGINE, che e’ il
punto in cui e’ inserito F. F e’ trasferito per ultimo
Mappa genetica a tempo (unità di misura: minuti)
Alle volte il ceppo F fuoriesce con
meccanismi simili (ma in ordine
inverso) alla sua entrata: alle volte
il meccanismo e’ imperfetto che
include frammenti di genoma
batterico.
• Questi plasmidi (F’) sono usati per
creare diploidi parziali (per studi
di dominanza ed epistasi di un
certo gene o gruppo di geni)
• I plasmidi sono il modo con cui si
trasporta, amplifica e manipola il
DNA. ESSENZIALI IN BIOLOGIA
MOLECOLARE
Trasformazione