Tavola Rotonda
Miglioramento genetico delle specie ortofrutticole, produzione
sementiera e norme fitosanitarie
COORDINATORE:
C. FIDEGHELLI
Istituto Sperimentale per la Frutticoltura, MiPAF Roma
La trasmissione per seme dei fitovirus, in particolare nelle specie ortofrutticole
Maurizio Conti
Istituto di Fitovirologia Applicata, CNR, Torino, Italia
INTRODUZIONE
La trasmissione per seme è oggi nota per circa un quinto dei fitovirus descritti (Barba, 1991;
Mink, 1993; Johansen et al., 1994). Per la precisione, facendo riferimento
al più recente
aggiornamento del ‘International Committee on Taxonomy of Viruses’ (ICTV : van Regenmortel et
al., 2000) essa risulta segnalata per membri di 26 dei 66 generi virali appartenenti a 8 delle 16
famiglie che includono virus dei vegetali, per un totale di 130 patogeni (Tab. 1). Si ritiene tuttavia
che il fenomeno sia alquanto più comune di quanto oggi non risulti, interessando forse un terzo o
più dei fitovirus, stima che appare fondata se si considera che l’incremento delle ricerche portò il
numero di virus trasmessi per seme dagli 8 conosciuti all’inizio degli anni Cinquanta (Smith, 1951)
a ben 156 verso la fine degli anni Ottanta (Agarwal e Sinclair, 1987). Successive verifiche hanno
ridimensionato questo dato mettendo in evidenza sia segnalazioni improprie per svariati motivi
(identificazione dei patogeni approssimativa e procedure sperimentali poco attendibili, ad esempio),
sia l’esistenza di sinonimie tra alcuni dei virus menzionati (Mink, 1993).
Poiché il seme risulta dalla fusione di due gameti, uno della pianta madre (ovulo), ed uno
della pianta fecondatrice (polline) – che è la stessa pianta nelle specie autogame - la trasmissione
per seme può avvenire attraverso l’uno, l’altro o entrambi i gameti, o le piante progenitrici. Pertanto
la trasmissione per seme e quella per polline sono così strettamente correlate che l’argomento può
essere trattato in modo esauriente solo riferendosi ad entrambe. Vi sono, tuttavia, problematiche
specificamente pertinenti ciascuna delle due: la trasmissione per seme sensu stricto considera in
dettaglio, ad esempio, i fattori che influenzano i movimenti dei virus sia dai tessuti della pianta
madre a quelli del seme che da quelli dell’embrione alla plantula che ne deriva, mentre la
trasmissione per polline esamina argomenti come gli effetti dei virus sulla germinazione del granulo
pollinico, la possibilità di replicazione virale all’interno di questo, l’eventualità che il polline infetti
– oltre al seme – la pianta stessa che viene fecondata.
La trasmissione per seme di un virus può interessare una o più delle sue piante ospiti e
presenta elevata specificità, tanto da riguardare talora soltanto qualche cultivar della stessa specie
(non tutte), oppure da realizzarsi tramite meccanismi differenti in specie e cultivar diverse. La
percentuale di trasmissione, intesa come proporzione di semenzali infetti rispetto al totale ottenuto
dal seme in esame, può variare da 0 (assenza di trasmissione) a 100% - valore massimo riscontrato
occasionalmente per ‘bean common mosaic virus’ (BCMV) in fagiolo, ‘squash mosaic virus
(SqMV) in melone e ‘alfalfa mosaic virus’ (AMV) in erba medica (Mink, 1993) - ma di norma si
attesta su valori medio- bassi (1 – 20%, indicativamente). Essa è condizionata dalle interazioni
virus/pianta ospite, dallo stadio fenologico della pianta al momento dell’infezione - punti che sono
discussi più avanti - da alcuni parametri ambientali (soprattutto la temperatura) e dalla presenza
dell’infezione in una o entrambe le piante parentali. Circa quest’ultimo punto, Bennet (1969) riporta
i casi seguenti: (i) ‘elm mosaic virus’ (EMV) che si trasmette per seme in Ulmus americana sia
attraverso il polline che l’ovulo: la percentuale di trasmissione è del 30,5% quando solo il polline
proviene da pianta infetta, del 48% quando entrambi i parentali sono infetti; (ii) ‘lychnis ringspot
virus’ (LRSV) che nella specie dioica Lychnis divaricata è trasmesso per seme in misura del 18,6%
quando solo la pianta impollinatrice è infetta, del 30,6% quando solo la pianta madre è infetta e del
33,6% quando lo sono entrambe.
L’entità della trasmissione per seme, tuttavia, non sempre è correlata all’ impatto economico
in pieno campo, nel senso che elevate incidenze di infezione sulle colture possono corrispondere
anche a bassa trasmissione per seme, e viceversa. Infezioni da seme di livello anche trascurabile ma
riguardanti virus soggetti a diffusione secondaria per mezzo di vettori (potyvirus, cucumovirus ed
altri virus non-persistenti trasmessi da afidi; nepo- e tobravirus trasmessi da nematodi) sono
sufficienti per introdurre i patogeni in aree coltivate infestate dai vettori ed originare improvvise,
gravi epidemie. La produzione di lattuga, ad esempio, può essere gravemente compromessa da
trasmissione per seme di ‘lettuce mosaic virus’ (LMV) in percentuale appena percettibile (0,001),
seguita da una massiccia diffusione del virus da parte degli afidi vettori (Ryder, 1973).
Analogamente, è stato stimato che la contaminazione dello 0,1% del seme di arachide da ‘peanut
mottle virus’ (PnMV), altro potyvirus trasmesso da afidi, può tradursi nella presenza finale di circa
50.000 piante infette/ha nelle coltivazioni (Adams e Kuhn, 1977).
La trasmissione per seme costituisce una strategia di sopravvivenza estremamente valida per
i virus poiché garantisce loro un legame “protetto” e duraturo tra cicli colturali successivi. Questa
funzione assume particolare importanza per virus che hanno gamma di ospiti ristretta, che infettano
soltanto specie annuali, che sono trasmessi da vettori in modo non persistente, che possiedono
vettori di limitata mobilità (virus trasmessi attraverso il terreno, ad esempio). I virus che ne
traggono maggior beneficio, tuttavia, sono senza dubbio ‘barley stripe mosaic virus’ (BSMV) e gli
alfa- e betacriptovirus poiché, per quanto oggi noto, la trasmissione per seme rappresenta per essi
l’unica via di sopravvivenza in natura.
PROCESSI DI TRASMISSIONE
La trasmissione per seme dei virus può avvenire secondo diversi meccanismi:
1. Per contaminazione della superficie esterna del seme o di parti del seme di origine materna,
come il tegumento esterno e interno (trasmissione non embrionale).
2. Per infezione dell’embrione (trasmissione embrionale) che può aver luogo mediante:
(a) infezione indiretta, per invasione di tessuti e organi di riproduzione (ovario, cellule madri
delle megaspore, cellule madri del polline) prima dell’embriogenesi;
(b) infezione diretta, per contaminazione dell’embrione nel corso dell’embriogenesi.
1. Trasmissione non embrionale
Se ne conoscono pochi casi, tutti riguardanti virus con particelle molto stabili, resistenti alla
degradazione e trasmissibili per inoculazione meccanica. Si tratta dei tobamovirus ‘tobacco mosaic
virus’ (TMV) , ‘tomato mosaic virus’ (ToMV), che possono ritrovarsi sia sulla superficie che nel
tegumento e nell’endosperma dei semi di pomodoro e peperone, e ‘cucumber green mottle mosaic
virus’ (CGMMV), che può contaminare il tegumento dei semi di cetriolo. La trasmissione ai
giovani semenzali si verifica per inoculazione meccanica, attraverso microlesioni provocate
principalmente dalla rottura di peli fogliari durante la manipolazione delle piante (soprattutto con le
operazioni di trapianto o ripiccaggio). Altro caso, unico nel suo genere, è rappresentato dal
necrovirus ‘melon necrotic spot’ (MNSV) che sopravvive nel tegumento interno dei semi di cetriolo
e di qui può essere trasmesso ai giovani semenzali dal fungo chitridiaceo Olpidium bornavanus.
2.a. Trasmissione embrionale indiretta
Studi ultrastrutturali hanno dimostrato che alcuni virus sono capaci di contaminare
l’embrione indirettamente (ossia prima della sua formazione) mettendo in evidenza la presenza di
particelle virali nelle cellule madri delle megaspore, in quelle del polline e nel polline stesso
(Carroll e Mayhew, 1976; Gaspar et al., 1984). Particelle di ‘tobacco rattle virus’ (TRV), ad
esempio, sono state osservate in cellule madri premeiotiche del polline di pomodoro e, in stadio più
avanzato, nel polline stesso, mentre la presenza di AMV è stata accertata nel citoplasma del polline
di erba medica. Entrambi i virus sono trasmessi per seme contaminato da polline infetto. ‘Tobacco
ringspot virus’ (TRSV) è stato osservato sia nel megagametofito che nel polline di soia, specie nella
quale il virus si trasmette in elevata percentuale verosimilmente perché – in base a quanto accertato
– è capace di invadere il tessuto meristematico e quindi di infettare la cellula madre della
megaspora.
Il gruppo di T. W. Carroll ha condotto studi molto approfonditi sulla trasmissione per seme
di BSMV nell’orzo rilevando le modificazioni citologiche che si verificano nei meristemi fiorali
durante la meiosi e la formazione dell’embrione in relazione alla distribuzione di un ceppo del virus
trasmissibile (MI-1) e di uno non trasmissibile (NSP) per seme nei tessuti di riproduzione. Essi
osservarono che: (i) la formazione dello strato di callosio intorno alle megaspore ed alle cellule
madri del polline ha luogo negli stadi premeiotici; (ii) non si rileva la presenza di plasmodesmi che
mettano in comunicazione la cellula madre della megaspora con le circostanti cellule nucellari; (iii)
dopo la fecondazione non si osservano plasmodesmi che connettano gli embrioni in via di sviluppo
con i tessuti circostanti (Carroll, 1972 e 1981; Carroll e Mayhew, 1976a). Dei due ceppi virali
studiati, quello trasmesso per seme risultò presente nei tessuti delle cellule madri delle megaspore e
del polline, oltrechè nell’ovario e nel polline, mentre quello non trasmissibile non fu mai osservato
in questi tessuti. Carroll e collaboratori ne conclusero che la trasmissibilità per seme di BSMV è
determinata dalla capacità del virus di invadere i meristemi riproduttivi maschili e femminili in stadi
molto precoci del loro sviluppo, contaminando successivamente l’embrione per via indiretta.
La marcata influenza dello stadio di sviluppo della pianta al momento dell’inoculazione sull’
entità della trasmissione per seme è stata più volte citata come elemento a sostegno della
contaminazione indiretta dell’embrione (ossia presenza del virus nei tessuti riproduttivi prima
dell’embriogenesi). In effetti, l’inoculazione della pianta in stadi fenologici avanzati o molto
avanzati induce generalmente livelli di trasmissione per seme assai modesti, annullandosi anche del
tutto se la pianta viene infettata durante o dopo la fioritura (Davis e Hampton, 1986; Goodman et
al., 1979).
Al contrario, l’infezione del seme per introduzione del virus nel sacco embrionale ad opera
del polline si verifica, ovviamente, nel corso della fioritura. Nel caso specifico, semi infetti possono
essere prodotti anche da piante madri esenti dal virus in questione. I casi di trasmissione per polline
più noti in letteratura sono quelli di ‘prune dwarf virus’ (PDV) e di ‘prunus necrotic ringspot virus’
(PNRSV) in diverse prunoidee (Prunus spp), e di BCMV in fagiolo, oltre a quello già citato di
BSMV in orzo.
L’eventualità che il virus possa diffondersi dagli embrioni contaminati via polline ai tessuti
adiacenti della pianta madre e infettarla, è stata presa in considerazione e studiata (cf., ad es.,
Johansen et al., 1994). Ricerche recenti hanno chiarito che la trasmissione da polline a pianta è
effettivamente possibile, ad esempio per ilarvirus di specie ortensi e da frutto, ma sembra aver
luogo soltanto con la mediazione di tripidi (e forse anche di altri insetti, soprattutto pronubi), i quali
trasportano sul loro corpo granuli pollinici ricoperti in superficie di particelle virali, che vengono
poi inoculate meccanicamente nelle piante visitate (Mink, 1993). Questo tipo di trasmissione è stato
riscontrato anche nella diffusione di ‘pelargonium zonate spot virus ‘ (PZSV) da piante spontanee a
coltivazioni di pomodoro (Vovlas et al., 1989), sempre ad opera di tripidi.
2.b. Trasmissione embrionale diretta. L’infezione diretta del seme si verifica in seguito al
trasferimento del virus direttamente dalla pianta madre all’embrione, ma i meccanismi di questo
processo sono ancora poco conosciuti. Prime informazioni sono state ottenute con ricerche sulla
combinazione pisello cv. Vedette/ ‘pea seedborne mosaic virus’ (PSbMV), nella quale non si
riscontra trasmissione per polline (Wang e Maule, 1993). I movimenti di PSbMV nei baccelli e nei
semi in via di sviluppo sono stati rilevati mediante ELISA, saggi di immuno-citochimica e di
ibridazione in situ. Il virus è stato rinvenuto nel funicolo prima della fecondazione ma è risultato
presente soltanto sporadicamente negli ovuli non fecondati. Dopo la fecondazione, esso è stato
facilmente individuato nei tessuti sia della testa e dell’endosperma che del sospensore embrionale.
Tuttavia una vasta diffusione del virus nella testa e nell’endosperma è stata osservata soltanto in una
cv di pisello nella quale PSbMV è trasmesso per seme ma non in un’altra, non soggetta a
trasmissione per seme, nella quale la diffusione del virus nei tessuti citati è risultata molto limitata.
In qual modo il virus penetri nell’embrione non è ancora chiarito ma è stata formulata l’ipotesi che
ciò si realizzi al punto di contatto tra la testa e il sospensore dell’embrione (Wang e Maule, 1993).
La parete cellulare al punto di contatto è alquanto convoluta ma priva di plasmodesmi: Wang e
Maule (1993) hanno suggerito che PSbMV potrebbe attraversarla con un meccanismo non meglio
specificato che non richiede la presenza di plasmodesmi o, in alternativa, che il virus stesso possa
indurre la formazione di plasmodesmi e in tal modo invadere l’embrione direttamente.
PSbMV, contrariamente a quanto riscontrato nella cv Vedette, è trasmissibile per polline
nella cv di pisello Dual in percentuale del 4% circa (Kohnen, 1992). Le indagini condotte sulla
combinazione PSbMV/pisello ‘Dual’ hanno evidenziato che 4 di 96 semi ottenuti da fiori di piante
sane sottoposti a castrazione e fecondati con polline di piante affette dal virus hanno dato origine a
semenzali infetti. La fecondazione di fiori di piante infette da PSbMV con polline proveniente da
piante sane ha indotto la trasmissione per seme del virus in misura del 38%. Questi risultati
indicano che nella combinazione virus/pianta ospite presa in esame l’infezione dell’embrione può
verificarsi sia direttamente che indirettamente.
L’ostacolo principale all’invasione diretta dell’embrione da parte dei virus sembra essere
l’apparente separazione tra polline, ovario e sacco embrionale da un lato e tessuti parentali
dall’altro. Tale separazione deriva dalla scomparsa dei plasmodesmi tra la cellule madri delle
megaspore e il tessuto nucellare, e tra le cellule madri del polline e le cellule del cosiddetto
‘tappeto’ nel corso della meiosi. Inoltre, durante la meiosi, si forma intorno alle megaspore ed alle
cellule madri del polline uno strato di callosio, tipicamente interrotto intorno al sacco embrionale
neoformato. Successivamente, lo strato di callosio attorno al polline ed al sacco embrionale viene
riassorbito e rimpiazzato da altri tessuti, mentre una spessa parete di pectocellulosa, priva di
plasmodesmi, si sviluppa attorno al sacco embrionale. Durante l’impollinazione i gametofiti
maschile e femminile restano separati poiché la parete del budello pollinico in via di sviluppo
produce uno strato di callosio - assente soltanto alla sua estremità terminale - via via che attraversa i
tessuti dello stilo. In considerazione di quanto sopra, sembrerebbe ovvio concludere che la
contaminazione del seme dalla pianta madre possa aver luogo più facilmente mediante penetrazione
del virus nei tessuti riproduttivi prima della scomparsa dei plasmodesmi e della formazione delle
barriere protettive nominate, e quindi per infezione indiretta dell’embrione, come descritto al
paragrafo precedente.
CONCLUSIONI
Fino a qualche decina di anni fa, la trasmissione per seme dei fitovirus è stata considerata un
evento relativamente poco comune e la sua importanza sottovalutata. Prima che se ne comprendesse
l’effettiva portata, essa ha causato difficoltà non indifferenti anche alla ricerca virologica, oltre che
alla lotta contro le virosi in pieno campo. Le piante nate infette da seme, infatti, spesso non
presentano sintomi evidenti (infezione latente), hanno aspetto normale e sono pertanto scambiate
per sane. Questo fenomeno, verificandosi in alcune combinazioni virus/piante indicatrici - 'sowbane
mosaic virus' (SwMV)/ Chenopodium spp., ad esempio - ha indotto talora a ritenere di aver isolato
questi virus da piante (fonte dell' inoculo) che ne erano esenti mentre, in realtà, erano contaminate
da seme le piante utilizzate nel saggio di inoculazione. Per quanto concerne la diffusione delle
virosi in pieno campo, la trasmissione per seme - anche in bassa percentuale, come si è già detto può svolgere diverse funzioni cruciali per il ciclo epidemiologico di virus fitopatogeni, quali:
garantire la sopravvivenza dei patogeni in assenza di coltivazioni e da un ciclo colturale
all’altro attraverso il seme di specie sia coltivate (es.: BCMV in fagiolo) che spontanee (CMV in
Stellaria media);
costituire sorgenti di infezione “primarie” dalle quali prende avvio annualmente la diffusione
“secondaria” dei virus trasmessi da vettori quali afidi (AMV, CMV, LMV,ecc.) e nematodi
(TRV);
consentire la disseminazione dei patogeni a “lunga distanza”, in seguito all’introduzione di semi
infetti in aree agricole anche molto lontane dal luogo di produzione. L’importanza di questa via
di diffusione è provata dal fatto che tutti i virus trasmessi per seme in specie coltivate, soggette a
commercializzazione, sono ubiquitari;
causare la contaminazione di specie da frutto moltiplicate agamicamente, innescando l'infezione
attraverso portinnesti riprodotti per seme. È il caso di PNRSV e PDV che si trasmettono per
seme in varie specie di Prunus sp. utilizzate come portinnesti di fruttiferi;
costituire nel terreno una duratura riserva di inoculo, consistente di semi infetti che conservano
la germinabilità per lunghi periodi di tempo.
La trasmissione per seme non embrionale è propria di virus resistenti alla degradazione,
molto infettivi e facilmente trasmissibili per contatto. Per questo motivo, pur essendo limitata a
pochi casi, riveste rilevante importanza economica, in particolar modo per specie come pomodoro e
peperone in coltura protetta (TMV, ToMV). La trasmissione per seme embrionale, diretta e
indiretta, interessa virus che invadono estesamente i tessuti parenchimatici e sono trasmissibili per
inoculazione meccanica. Non sono conosciuti invece casi di trasmissione per seme di virus
localizzati esclusivamente nel tessuto floematico (luteovirus, fitoreovirus, fijivirus, ecc.), i quali
sono trasmessi da insetti in modo persistente - circolativo o propagativo - ma non mediante
inoculazione meccanica. Lo stesso vale per i fitoplasmi, agenti "virus-simili", floematici, di malattie
delle piante, che sono trasmessi in modo propagativo da cicaline e psille.
La presenza di un virus nel seme, anche nell’embrione, non sempre risulta nell’infezione del
semenzale che ne deriva: in molti casi, infatti, nel corso dei processi metabolici connessi con la
germinazione, il virus viene degradato e perde l'infettività. Ciò non pregiudica tuttavia l’utilità del
controllo sanitario delle sementi attraverso le quali determinati virus possono trasmettersi, per
accertare che ne siano esenti. Le metodologie diagnostiche consistono in saggi di germinazione e
infettività, sierologici, di immuno-microscopia elettronica e nell'utilizzazione di mezzi molecolari
sempre più sensibili e affidabili (Russo, 1982; Barba, 1991a).
BIBLIOGRAFIA
Adams D.B. e C.W. Kuhn, 1977. Seed transmission of peanut mottle virus in peanut.
Phytopathology, 67, 1126-1129.
Agarwal V.K. e J.B. Sinclair. 1987. Seedborne pathogens. In: “Priciples of seed pathology”, Boca
Raton, CRC Press, 176 pp.
Barba M., 1991. I virus trasmessi per seme. Petria 1 (Suppl. 1), 1-9.
Barba M., 1991a. Diagnostica con metodi sierologici e sonde nucleiche dei virus delle sementi.
Petria, 1 (Suppl. 1), 11-18.
Bennett C.W., 1969. Seed transmission of plant viruses. Adv. Virus Res. 14, 221-261.
Carroll T.W. 1972. Seed transmissibility of two strains of barley stripe mosaic virus. Virology, 48,
323-336.
Carroll T.W., 1981. Seedborne viruses: virus-host interactions. In: “Plant diseases and vectors:
ecology and epidemiology”, K. Maramorosch & K.F. Harris Ed., N.Y., Academic Press, 293317.
Carroll T.W. e D.E. Mayhew. 1976. Occurrence of virions in developing ovules and embryo sacs in
relation to the seed transmissibility of barley stripe mosaic virus. Can. J. Botany, 54, 24972512.
Carroll T.W. e D.E. Mayhew, 1976a. Anther and pollen infection in relation to the pollen and seed
transmissibility of two strains of barley stripe mosaic virus in barley. Can. J. Botany, 54,
1604-1621.
Davis R.F. e R.O. Hampton. 1986. Cucumber mosaic virus isolates seedborne in Phaseolus
vulgaris: serology, host-pathogen relationships, and seed transmission. Phytopathology 76,
999-1004.
Gaspar J.O., J. Vega , I.J.B. Camargo e A.S. Costa. 1984. An ultrastructural study of particle
distribution during microsporogenesis in tomato plants infected with the Brazilian tobacco
rattle virus. Can. J. Botany , 62, 372-378.
Goodman R.M., G.R. Bowers Jr. e E.H. Paschal. 1979. Identification of soybean germplasm lines
and cultivar with low incidence of soybean mosaic virus transmission through seed. Crop Sci.
19, 264-267.
Johansen E., M.C. Edwards e R.O. Hampton. 1994. Seed transmission of viruses: current
perspectives. Ann. Rev. Phytopathol. 32, 363-386.
Kohnen P.D. 1992. Partial characterization of the P4 pathotype of pea seedborne mosaic virus . Ph.
D. Thesis, Oregon St. Univ., Corvallis, OR.
Mink G.I. 1993. Pollen- and seed-transmitted viruses and viroids. Ann. Rev. Phytopathol, 31, 375402.
Regenmortel van M.H. V., C. M. Fauquet e D.H.L. Bishop. 2000. Virus taxonomy. Seventh Report
of the ICTV, Academic Press, 1162 pp.
Russo M. 1982. Moderne metodiche per il rilevamento delle infezioni virali nei semi. II.
Immunomicroscopia elettronica. La Difesa delle Piante , 5-6, 395-404.
Ryder E.G., 1973. Seed transmission of lettuce mosaic virus in mosaic resistant lettuce. J. Am. Soc.
Hortic. Sci., 98, 610-614.
Smith K.M. 1951. A latent virus in sugarbeets and marigolds. Nature 167, 1061.
Vovlas C., D. Gallitelli e M. Conti. 1989. Preliminary evidence for an unusual mode of
transmission in the ecology of pelargonium zonate spot virus (PZSV). In: Proc. 4th Int. Plant
Virus Epid. Workshop, Montpellier (FR) 3-8 sett. 1989, 302-305.
Wang D. e A.J. Maule. 1993. Seed transmission of pea seed-borne mosaic virus in pea- a process
full of surprises. Int. Congr. Virol., Glasgow (SC). Abs. W64-8.
Tabella 1. Virus trasmessi per seme
Famiglia
Partitiviridae
Genere
Alphacryptovirus
Numero(*)
30
Betacryptovirus
Comoviridae
Comovirus
6
Fabavirus
1
Nepovirus
25
Potyvirus
20
Rymovirus
1
Bymovirus
1
Tetraviridae
Sobemovirus
4
Tombusviridae
Carmovirus
4
Machlomovirus
1
Necrovirus
1
Tombusvirus
1
Tobamovirus
5
Tobravirus
2
Hordeivirus
2
Furovirus
2
Pecluvirus
2
Alfamovirus
1
Bromovirus
2
Cucumovirus
3
Ilarvirus
8
Idaeovirus
1
Trichovirus
1
Tymovirus
3
Carlavirus
3
Potyviridae
Togaviridae
Bromoviridae
Closteroviridae
(*) Dati desunti da Mink (1993) e aggiornati
Funghi trasmessi per seme nelle principali specie orticole
Giovanni Vannacci
Dipartimento di Coltivazione e Difesa delle Specie Legnose “G. Scaramuzzi”
Università di Pisa
INTRODUZIONE
Il seme rappresenta, per gran parte delle colture orticole, l’organo di riproduzione più
importante. Un seme di buona qualità è uno dei requisiti essenziali per garantire il successo della
coltura e da sempre grande attenzione è stata devoluta a quelle caratteristiche cosiddette
agronomiche, quali la germinabilità e la purezza, che più immediatamente erano correlabili con i
risultati di campo. E’, peraltro, ampiamente dimostrato che il seme può nascondere organismi
patogeni in grado di annullare completamente qualunque altra positiva caratteristica qualitativa. La
normativa italiana relativa specificatamente ai patogeni trasmessi per seme è decisamente limitata
ed una sintetica analisi è già stata pubblicata recentemente (Vannacci, 1998). In questa sede
discuteremo, nell’ottica di chi vuole valutare il ruolo del seme sul rispetto delle norme tecniche per
la commercializzazione delle piantine di ortaggi, il DM del 14 aprile 1997, con l’obiettivo di
richiamare l’attenzione su quei patogeni che, inclusi nell’allegato II, risultano essere anche
trasmessi per seme. Purtroppo, se si esamina la letteratura specifica ci si renderà conto che molti
altri patogeni, non ricompresi nella normativa in discussione, sono trasmessi per seme. Il problema
è, quindi, decisamente più complesso di quanto possa risultare dalla consultazione della tabella
riportata in questo lavoro; in fin dei conti questa norma regola la commercializzazione delle
piantine per il trapianto, settore di grande importanza, ma molto spesso, in orticoltura, si ricorre alla
semina diretta ed in tal caso le condizioni fitosanitarie del seme non risultano regolamentate da
norme specifiche. E’ lecito pensare che sia il mercato a premiare chi produce seme migliore (“più
sano”); purtroppo questo non sempre è vero in quanto non è agevole far risalire la colpa di eventi
epidemici all’uso di seme infetto quando ad esserne causa è un patogeno, ad esempio, ad habitat
tellurico e, quindi, il nesso causa – effetto si perde. D’altra parte non si deve cadere nell’eccesso
opposto, vale a dire quello di considerare seme di bassa qualità quello che alberga organismi
patogeni indipendentemente dal livello di infezione. Trattandosi di qualità si deve fare astrazione da
quei patogeni che rientrano nelle norme relative all’introduzione di patogeni non presenti nel
territorio italiano, patogeni da quarantena, e, quindi, si parla di organismi per i quali è necessario
fornire indicazioni quantitative ai fini di una corretta valutazione del loro ruolo nel determinare la
qualità del seme. Sarebbe necessario a tale proposito individuare due livelli di soglia: il primo che
separi il seme che non richiede interventi di concia da quello che, a seguito di trattamenti mirati,
può manifestare appieno la propria potenzialità produttiva, ed il secondo che separi quest’ultimo da
quei semi che non debbono essere messi in commercio (a titolo di esempio si possono vedere le
esperienze maturate per il sistema Orzo / Pyrenophora graminea, Vannacci et al., 1996). Non si
deve pensare che tutto questo debba obbligatoriamente essere demandato al legislatore, l’agricoltura
italiana è fortemente indirizzata verso produzioni di alta qualità ed anche la produzione sementiera,
se il mercato la premia, può offrire prodotti di “alta qualità” basati su programmi di certificazione
volontaria.
DM
DEL
14
APRILE
1997
RELATIVO ALLE NORME TECNICHE PER LA COMMERCIALIZZAZIONE
DELLE PIANTINE DI ORTAGGI
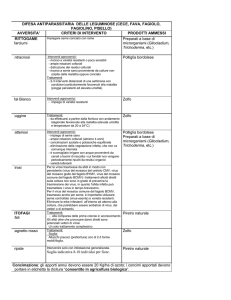
Nel decreto sono ricordate 24 colture orticole e 95 funghi fitopatogeni, sia le prime che i
secondi indicati di solito a livello di specie. Il decreto esclude esplicitamente dal suo campo di
applicazione le sementi, tuttavia quasi la metà dei funghi indicati sono trasmessi per seme; è, perciò,
evidente lo stretto nesso che esiste tra stato sanitario di quest’ultimo e l’assenza sulle piantine di
patogeni o malattie espressamente ricordati dalla normativa. Può risultare interessante ricordare che
alcuni degli organismi patogeni indicati nell’allegato, basti ricordare Sclerotinia, Sclerotium e
Fusarium, sono organismi ad habitat tellurico ed il seme rappresenta la via attraverso la quale essi
possono insediarsi in terreni dove non sono presenti. Il pericolo è tanto più grave se si considera che
non sempre i sintomi dovuti ad alcuni di essi si manifestano allo stadio di plantule e, quindi, la loro
presenza può sfuggire all’esame del materiale in uscita dai vivai.
In Tab. 1 sono state rielaborate le informazioni presenti nell’allegato II del DM lasciando indicati,
per ogni coltura, solo i funghi patogeni trasmessi per seme (Richardson, 1990). Di seguito si
discuteranno alcuni esempi. Informazioni più dettagliate potranno essere trovate in Neergaard, 1979
e Champion, 1997; per quanto riguarda i metodi di analisi fitosanitaria si può anche fare riferimento
a Vannacci, 1988.
Botrytis spp.
È ricordata Botrytis cinerea su Brassica e Lactuca e Botrytis spp. su Allium. B. cinerea può
essere presente sia sui tegumenti seminali sotto forma di spore o, più raramente, come sclerozi, sia
all’interno del seme sotto forma di micelio. Causa il marciume delle giovani piantine su cui si forma
l’inoculo secondario che diffonde l’infezione. L’analisi fitosanitaria del seme può essere condotta
su mezzo agarizzato o su carta bibula. Su Allium, oltre a B. cinerea sono presenti altre specie di
Botrytis (B. allii, B. byssoidea) ma la più importante è B. allii su Allium cepa (Fig. 1). Al pari di B.
cinerea, anche B. allii può essere presente sia sui tegumenti seminali sotto forma di spore, sia
all’interno del seme, sotto forma di micelio. L’analisi fitosanitaria prevede l’incubazione su agar
malto, ma non sempre la distinzione da altre specie di Botrytis è agevole.
Sclerotium cepivorum
È un importante patogeno su Allium, ma la sua trasmissione per seme non sembra
particolarmente frequente. Può ritrovarsi come sclerozi in lotti di seme di bassa qualità. E’ un
patogeno ad habitat tellurico.
Alternaria spp.
Sono ricordate A. porri (su Allium), A. brassicae (su Brassica) e A. solani (su Lycopersicon)
(Fig. 2 e Fig. 3). Possono essere presenti sia all’interno che all’esterno del seme; alla germinazione
riprendono l’attività e possono causare marcescenza dei semi e dei germinelli in pre-emergenza o
necrosi sulle plantule che costituiranno la fonte principale d’inoculo. Sono disponibili diversi
metodi di analisi ed il loro riconoscimento è facilitato dal fatto che hanno conidi grandi, usualmente
singoli e, A. porri e A. solani, con lunghi becchi.
Phoma spp.
Sono ricordate P. apiicola (su Apium), P. betae (su Beta) e P. lingam (su Brassica). Le tre
specie sono presenti sotto forma di micelio nei tegumenti seminali o del glomerulo, talvolta possono
essere presenti picnidi immersi nei tegumenti. Sono in grado di impedire la germinazione dei semi e
di attaccare i germinelli sia in pre- che in post-emergenza. I metodi di reperimento prevedono
l’incubazione su agar o in camera umida (Fig. 4 e Fig. 5). Per favorire lo sviluppo dei patogeni e
rendere i metodi più sensibili si può aggiungere 2,4 D al mezzo di incubazione (carta bibula
inumidita per P. lingam, agar acqua per P. betae). Poiché la virulenza in isolati diversi di P. lingam
può differire notevolmente, è disponibile un test che ne consente la valutazione.
Pythium spp.
Sono ricordati, senza indicazione della specie, su Apium, Cucumis e Lycopersicon. Sono
presenti come micelio, e talvolta come oospore, nei tegumenti. Causano la moria dei semenzali. Per
individuare la loro presenza è necessario ricorrere all’incubazione su agar nutritivo. È un patogeno
ad habitat tellurico.
Septoria apiicola
È presente, nei semi di Apium, come picnidi immersi nei tegumenti seminali o come
micelio, anche se in questo secondo caso l’inoculo sembra meno efficiente nel trasmettere la
malattia. Causa sintomi molto lievi sui germinelli, essenzialmente piccole lesioni contenenti picnidi
immaturi. E’ dopo il trapianto che i picnidi maturano e liberano grandi quantità di conidi che danno
inizio, se le condizioni ambientali lo permettono, a violenti epidemie. La determinazione della
presenza del patogeno sui semi può essere effettuata osservandoli allo stereomicroscopio per
individuare i picnidi, in alternativa si può procedere a inoculazioni artificiali su piantine sane
utilizzando l’acqua in cui sono stati immersi per 6 ore i semi del lotto da analizzare.
Fusarium spp.
Il D.M. indica Fusarium spp. per Asparagus, Cucumis, Cucurbita e Solanum melongena
mentre ricorda F. oxysporum per Lycopersicon e F. oxysporum f.sp. cepae su Allium cepa. Su
Asparagus sono segnalati diversi Fusarium, tra cui F. oxysporum, F. culmorum e F. moniliforme, su
Cucumis sono segnalati F. oxysporum (f. sp. niveum e f. sp. cucumerinum) e F. solani f. sp.
cucurbitae, su Cucurbita F. solani f. sp. cucurbitae e Fusarium spp. (questi ultimi agenti di
marciume dei semenzali), su Solanum melongena F. oxysporum e F. solani. Questi Fusarium
possono essere divisi in patogeni vascolari (F. oxysporum) e patogeni parenchimatici (tutti gli altri).
F. oxysporum vascolari sembrano essere trasmessi raramente per seme e si ritrovano all’esterno del
seme o negli strati più superficiali dei tegumenti; gli altri possono insediarsi più profondamente nei
tessuti seminali ed in alcuni casi (F. solani f. sp. cucurbitae, Fig. 6) la trasmissione per seme riveste
particolare importanza. Sono, di solito, patogeni ad habitat tellurico. In ogni caso causano lievi
sintomi sulle plantule. L’analisi fitosanitaria prevede l’incubazione su mezzo agarizzato ed il
successivo riconoscimento della colonia. Un metodo più sensibile prevede l’impiego di un mezzo
selettivo per Fusarium (es. agar di Nash and Snyder, Tuite, 1969), il successivo trapianto delle
colonie su PDA e quindi il loro riconoscimento.
Rhizoctonia solani
Ricordata su Brassica, Lycopersicon e S. melongena può essere presente all’interno dei
tegumenti seminali; causa gravi sintomi sui germinelli e sulle giovani plantule che vengono
attaccate al colletto con conseguente morte. L’analisi fitosanitaria può essere condotta su agar
nutritivo o su carta bibula inumidita. In quest’ultimo caso è necessario riconoscere i sintomi sulle
plantule, cosa non sempre agevole in quanto confondibili con quelli causati da Botrytis, Pythium,
Fusarium.
Sclerotinia
Sono ricordate Sclerotinia spp. su Brassica pekinensis e S. sclerotiorum (Fig. 7) su Cynara e
S. melongena. Il patogeno produce sclerozi che si ritrovano frammisti ai semi e che gli consentono
di sopravvivere a lungo nel terreno ma lo si può ritrovare anche come micelio nei tegumenti. Le
piantine che hanno origine da semi infetti vanno incontro a morte in pre-emergenza; gli sclerozi
distribuiti nel terreno assieme ai semi possono germinare per micelio, ed in tal caso possono colpire
le piantine giovani, o producendo le fruttificazioni gamiche, gli apoteci. Questi differenziano gli
aschi che liberano nell’aria le ascospore che sono così in grado di causare infezioni sulle parti alte
delle piante. La presenza del patogeno può essere determinata mediante l’esame della materia inerte
frammista al seme o mediante incubazione su mezzo agarizzato.
Colletotrichum lagenarium
Noto anche come C. orbiculare, viene ricordato su Cucumis e Citrullus. Si può ritrovare sia
sulla superficie dei semi che all’interno dei tessuti. Semi infetti possono non germinare o dar luogo
a germinelli con sintomi di gravità variabile in dipendenza della quantità di inoculo. Non sono
descritti metodi specifici ma per altre specie di Colletotrichum si ricorre a metodi che prevedono
l’incubazione su carta bibula o su mezzo agarizzato.
Dydimella lycopersici
Ricordata su Lycopersicon, è un patogeno principalmente ad habitat tellurico, ma per colture
effettuate in terreno sterilizzato, il seme rappresenta un importante fonte d’inoculo. È normalmente
presente come picnidi immersi nei tegumenti seminali che, talvolta risultano visibili. Poiché l’esame
del seme secco non è sufficiente per determinare la presenza/assenza del patogeno è necessario
ricorrere all’incubazione in camera umida ed al successivo esame dei sintomi sulle plantule alla
ricerca dei picnidi.
CONCLUSIONI
La discussione dei pochi esempi sopra riportati deve servire a richiamare l’attenzione del
legislatore, ma anche dei vivaisti e degli agricoltori in generale, sull’importanza che riveste lo stato
sanitario del seme non solo sulla rispondenza del materiale di moltiplicazione alle norme vigenti ma
anche, e direi principalmente, sullo stato sanitario della coltura che da tale seme avrà origine. Se si
considera che nel DM sono elencati altri organismi (batteri, virus) la cui trasmissione per seme è
certa, la necessità di utilizzare seme esente da patogeni diventa pressante. Particolare attenzione
dovrà essere prestata a quei patogeni che non manifestano sintomi ben evidenti sulle plantule (es.
Septoria) senza trascurare, tuttavia, che anche patogeni in grado di attaccarle, se presenti in
“quantità” (percentuale di semi o quantità di inoculo per seme) basse, potranno sfuggire ai controlli
in vivaio con conseguenti danni alla coltura e contenziosi tra vivaisti e acquirenti delle piantine. Tra
i patogeni elencati nel DM, infine, alcuni sono ad habitat tellurico, quali, ad esempio, S. cepivorum,
S. sclerotiorum, Fusarium spp. Il danno che essi potranno causare non sarà, perciò, limitato alla
coltura che da tali semi avrà origine, ma il patogeno si insedierà nel terreno costringendo
l’agricoltore a costosi, quando e se possibili, interventi di sterilizzazione del terreno. Già oggi alcuni
produttori di seme immettono sul mercato seme esente da particolari patogeni (es. basilico esente da
Fusarium oxysporum f. sp. basilici) in quanto l’esperienza ha dimostrato come questa sia la
strategia più efficace per tenere sotto controllo importanti fitopatie. L’impiego di seme esente da
specifici patogeni, se da un lato costituisce un prerequisito per una coltura sana, dall’altro comporta
dei disagi dovuti, principalmente, ai maggiori costi che gli agricoltori dovrebbero sostenere per
l’acquisto del seme ed alle difficoltà pratiche che potrebbero sorgere per i produttori di seme in
particolare per la lunghezza dei tempi necessari a condurre le analisi fitosanitarie. Purtroppo, per i
patogeni di nostro interesse, sono praticamente assenti metodi diagnostici innovativi (Vannacci,
1998) e si deve fare riferimento ai metodi classici con tempi, per singola analisi, che di norma si
aggirano almeno sui 7 – 10 giorni. La ricerca, anche in questo settore, potrà dare le risposte giuste
se risorse, umane e finanziarie, saranno indirizzate verso questi obiettivi.
BIBLIOGRAFIA
Champion, R. 1997. Identifier les champignons transmis par les semences. I.N.R.A. Paris, France,.
398 p.
Neergaard, P. 1979. Seed pathology. The Macmillan press LTD, London and Basingstoke, 1191 p.
Richardson, M.J. 1990. An annotated list of seed-borne diseases. International Seed Testing
Association, Zurich, Switzerland.
Tuite, J. 1969. Plant Pathology Methods. Burgess Publishing Company, 239 p.
Vannacci, G. 1988. Analisi sanitaria delle sementi: aspetti metodologici. Quaderno dell’E.N.S.E.
Milano 41, 32p.
Vannacci G., Aragona M., Fanti S. e Porta-Puglia A. 1996. Principali aspetti fitopatologici legati al
seme. L’Informatore Agrario, 52 (32), 57-63.
Vannacci, G. 1998. Diagnosi di patogeni fungini trasmessi per seme oggetto di legislazione
fitosanitaria. Notiziario sulla Protezione delle Piante 9:85-95.
Fig. 1. Conidiofori e conidi di Botrytis allii (a sinistra) e B. cinerea (a destra). La lunghezza di
queste strutture è carattere distintivo tra le due specie
Fig. 2. Dopo incubazione in camera umida sono evidenti, su seme di cavolo, i grossi conidi di
Alternaria brassicae
Fig. 3. Conidi e micelio di Alternaria solani su seme di pomodoro
Fig. 4. Phoma lingam è riconoscibile per la produzione di picnidi essudanti gocce di conidi immersi
in una mucillaggine color ametista
Fig. 5. Particolare del picnidio di Phoma lingam
Fig. 6. Fruttificazioni agamiche di
Fusarium solani su seme di
zucchino
Fig. 7. Sclerozi di Sclerotinia sclerotiorum da seme di lattuga
Tabella 1. Elenco dei patogeni trasmessi per seme presenti nell’allegato II del DM 14 aprile 1997
relativo alle norme tecniche per la commercializzazione delle piantine di ortaggi
Genere o specie
Allium ascalonicum
Organismi nocivi e
malattie
Botrytis spp.
Sclerotium cepivorum
Genere o specie
Cichorium intybus
Cucumis melo
Organismi nocivi e malattie
Fusarium spp.
Botrytis spp.
Cucumis sativus
Sclerotium cepivorum
F. oxysporum f.sp. cepae
Sclerotium cepivorum
Cucurbita maxima
Fusarium spp.
Pythium spp.
Alternaria porri
Sclerotium cepivorum
Sclerotium cepivorum
Cucurbita pepo
Fusarium spp.
Cynara cardunculus
Sclerotinia sclerotiorum
Cynara scolymus
Sclerotinia sclerotiorum
Asparagus officinalis
Phoma apiicola
Pythium spp.
Septoria apiicola
Fusarium spp.
Foeniculum vulgare
Cercospora foeniculi
Beta vulgaris
Phoma betae
Lactuca sativa
Botrytis cinerea
Bremia lactucae
Brassica oleracea
Alternaria brassicae
Mycosphaerella spp.
Phoma lingam
Plasmodiophora
brassicae
Rhizoctonia solani
Lycopersicon
lycopersicum
Brassica pekinensis
Alternaria brassicae
Botrytis cinerea
Mycosphaerella spp.
Phoma lingam
Plasmodiophora
brassicae
Sclerotinia spp.
Phytophthora capsici
Rheum spp.
Alternaria solani
Cladosporium fulvum
Didymella lycopersici
Fusarium oxysporum
Phytophthora nicotianae
Pythium spp.
Rhizoctonia solani
Verticillium spp.
-
Allium cepa
Allium fistulosum
Allium porrum
Allium sativum
Apium graveolens
Capsicum annuum
Cichorium endivia
-
Solanum melongena
-
Fusarium spp.
Rhizoctonia solani
Sclerotinia sclerotiorum
Verticillium spp.
Correlazione tra tecniche di miglioramento genetico e trasmissione dei virus
per via gamica
D. Bassi1, G.P.Martelli2
1
Istituto di Coltivazioni Arboree, Università degli Studi di Milano
2
Dipartimento di Protezione delle Piante e Microbiologia Applicata, Università degli Studi di Bari
Introduzione
Il seme e, in misura minore, il polline costituiscono vie naturali attraverso le quali patogeni
intracellulari delle piante quali virus e viroidi si trasmettono alla progenie degli ospiti (trasmissione
verticale) e si diffondono nell'ambiente. I meccanismi che sottendono alla trasmissione ed i
problemi che ne derivano sono stati illustrati, oltre che da M. Conti nel presente Convegno, in
numerose messe a punto bibliografiche, alle più recenti delle quali (Mink, 1993; Johansen et al.,
1994; Maury et al., 1998), si rimanda per gli approfondimenti e la bibliografia.
La trasmissione attraverso il seme è proprietà intriseca dei membri di non meno di 25 dei
generi virali attualmente noti, mentre nei viroidi essa è stata riscontrata in una diecina di casi.
Qualche anno addietro Stace-Smith e Hamilton (1988) riportavano che circa il 18% dei virus noti
all'epoca erano passibili di tramissione per seme in uno o più ospiti e stimavano che un terzo della
popolazione dei fitovirus fosse in grado di seguire la stessa via in almeno un ospite. La situazione,
pertanto, è tutt'altro che incoraggiante, soprattutto se si considera che, normalmente, la
trasmissione per seme si accompagna ad altri metodi di diffusione (per vettori, per esempio) che
aumentano fortemente l’efficienza della disseminazione dei virus in natura.
Benché, in linea di massima, la trasmissione per seme sia più frequente nelle piante erbacee
che in quelle legnose, cui si fa specifico riferimento nella presente nota, nelle prime essa non
costituisce un intoppo al miglioramento genetico perché trovare parentali sani non è difficile e,
comunque la selezione avviene su di un numero elevato di individui la maggioranza dei quali non è
infetta.
Invece, nel caso dei fruttiferi (intesi in senso lato, sì da includere vite e olivo), la presenza di
virus e viroidi in molte collezioni varietali utilizzate come fonte di materiale per il miglioramento
genetico (polline o semi), rende estremamente attuale la messa a punto di appositi protocolli che
consentano di escludere, o comunque controllare, la presenza di agenti che possano compromettere
lo stato sanitario e, quindi, la diffusione del materiale di propagazione. E’ noto infatti che le piante
"sane" (esenti da virus patogeni) offrono migliori prestazioni in termini di sviluppo vegetativo,
minore sensibilità ad altre malattie, maggiore produttività e qualità dei frutti (si veda tra gli altri
Jakubczyk et al., 1997).
Sebbene sembri definitivamente accertato che il virus della vaiolatura del susino (PPV),
agente della Sharka, di gran lunga la più dannosa virosi delle drupacee, non si trasmetta per seme
(Myrta et al., 1998; Pasquini et al., 2000), è noto che altri virus sono molto diffusi in queste specie.
Tra questi, sono da ricordare gli agenti del mosaico del melo (ApMV) nel mandorlo, del nanismo
del susino (PDV) nel ciliegio, della maculatura anulare necrotica dei Prunus (PNRSV) nel pesco,
ciliegio e susino e della maculatura clorotica fogliare (ACSLV) nell’albicocco (Desvignes, 1999),
presenti negli impianti commerciali con percentuali di infezione che giungono anche all’80%. Di
essi, alcuni si diffondono col polline e/o col seme.
In particolare, i virus da tenere d'occhio sono: (a) nelle drupacee, gli ilarvirus PDV e
PNRSV, entrambi trasmessi per seme con percentuali variabili dal 2 al 90% e, soprattutto il
secondo, anche per polline; i nepovirus dell'accartoccimento fogliare del ciliegio (CLRV) trasmesso
nel noce per seme e polline anche alle piante madri e della maculatura anulare latente della fragola
(SLRSV); (b) nella vite, i nepovirus latente bulgaro (GBLV), del mosaico con rosettamento del
pesco (PRMV), mentre preoccupa assai meno il virus dell'arricciamento (GFLV) che è presente sia
nell'endosperma che nel polline ma che si trasmette con estrema rarità ai semenzali. Un potenziale
pericolo potrebbe essere rappresentato dal foveavirus associato alla butteratura della Vitis rupestris
(RSPaV) che è presente nel polline; (c) nell'olivo, il CLRV e il necrovirus latente 1 (OLV-1) di cui
è stata di recente accertata la trasmissione per seme (M. Saponari, T. El Beaino e V. Savino,
comunicazione personale). Gli agrumi, invece, apparentemente godono di una condizione
privilegiata perché nessuno dei virus importanti segnalati nell’area Mediterranea sembra passare
alla progenie per via gamica (si veda tra gli altri D’Onghia et al., 2000).
Come si è accennato, alcuni viroidi sono stati rinvenuti nel polline e nel seme degli ospiti ed
utilizzano anche queste vie per la loro sopravvivenza a disseminazione. Mentre non si hanno notizie
sulla trasmissione per seme dell’agente del mosaico latente del pesco (PLMVd), uno dei più
temibili viroidi dei fruttiferi, e dell’exocortite degli agrumi (CEVd) negli agrumi stessi, questo
meccanismo di trasmissione è stato riscontrato per tutti e cinque i viroidi della vite: picchiettatura
gialla 1 e 2 (GYSVd-1 e GYSVd-2), nanismo del luppolo (HSVd), viroide australiano della vite
(GAVd) e CEVd (Wan Chow Wa e Symons ,1999).
Il problema pertanto esiste e non è recente, ma due fatti lo rendono particolarmente
urgente. Da una parte, la scarsa attenzione posta in passato (anche dalle Istituzioni scientifiche) allo
stato sanitario del materiale vegetale di scambio, che ha contribuito alla diffusione di piante
infette, ovvero ne ha dilazionato l'entrata nel mercato in attesa della disponibilità di materiale sano
e, dall’altra, la promulgazione di provvedimenti legislativi nazionali ed internazionali che hanno
reso obbligatoria la diffusione di materiale sano, lasciandone la responsabilità ai costitutori. Del
resto, evidenti motivi legati alla scarse prestazioni quali-quantitative di cultivar virosate, rendono
sempre più pressante, anche per i costitutori, la messa in opera di procedure che consentano l’uso
di materiale sano in tutte le fasi del lungo cammino che porta dall’effettuazione dell’incrocio alla
diffusione della nuova varietà.
Di seguito si illustrano alcune precauzioni da osservare per ottenere materiale propagativo
di fruttiferi esente da malattie infettive in tutte le fasi del processo di miglioramento genetico,
tenendo anche presente quanto già pubblicato in materia (Barba et. al., 1999; Lankes, 2000; Vicchi
et al., 1997).
Disponibilità di piante madri sane
La presenza di agenti infettivi trasmissibili attraverso la via zigotica (polline e seme) rende
necessaria la disponibilità di piante madri (sia portaseme che per la raccolta del polline) che ne
siano prive. Ciò renderebbe possibile la teorica produzione di semi completamente esenti da agenti
infettivi e, quindi, la loro disponibilità per l’incrocio. Nella pratica, però, tale ideale condizione è
difficilmente conseguibile perché: (a) le piante madri sono normalmente mantenute in pieno campo,
ove sono esposte alle visite dei vettori. Anche se è possibile ipotizzarne il mantenimento in strutture
a prova di insetti, l’allevamento avverrebbe in vaso, con conseguente riduzione delle dimensioni e
scarsa resa in frutti e semi. Dal punto di vista pratico, l’allevamento delle piante madri in condizioni
controllate è possibile solo per la raccolta di polline, per la quale sono sufficienti soggetti di taglia
ridotta; (b) non è sempre facile prevedere con largo anticipo la necessità di impiego di un certo
genitore, per cui attendere la disponibilità di materiale sano potrebbe dilazionare troppo l’uso di
prezioso materiale genetico per gli incroci.
Occorre comunque tener presente che la non utilizzazione di piante madri infette, o
comunque la loro eliminazione può in alcuni casi essere sconsigliabile, anche se prescritta da
specifica legislazione, specie se la distruzione comporta la perdita irrimediabile di materiale
genetico particolarmente interessante e non più reperibile, come possono esserlo semenzali o
selezioni ottenute nello svolgimento di programmi di incrocio, ovvero esemplari di vecchio
germoplasma non più recuperabile da altre fonti. In questi casi, l’unicità genetica del materiale
infetto renderebbe indispensabile il suo risanamento per non compromettere irreparabilmente il
proseguimento del programma di miglioramento. Tuttavia, nelle more degli interventi risanatorii, la
conservazione del germoplasma va effettuata in condizioni di isolamento (serre a vetro o a rete a
prova di insetti), obbligatoriamente nel caso di infezioni da agenti particolarmente temibili (da
quarantena, ad esempio).
Controllo dello stato sanitario delle progenie (semi)
La trasmissione per seme dipende dalla capacità di un virus di invadere e replicarsi nei
tessuti riproduttivi dell’ospite e, susseguentemente, di passare indenne attraverso le modificazioni
fisiologiche associate alla maturazione dei semi. Se ciò avviene, il virus si installa nel seme nel
quale, tuttavia, può avere localizzazioni diverse (tegumenti, endosperma, embrione). Ciò non è di
poco conto perché solo l’invasione embrionale determina il passaggio del virus al semenzale e,
quindi, l’effettivo trasferimento alla progenie. Ne risulta che non tutti i semi che contengono virus
generino delle plantule infette per cui, nel valutare la potenziale trasmissibilità per seme di un
qualsivoglia agente infettivo è indispensabile saggiare semi sezionati nelle varie componenti.
Controllo dello stato sanitario delle progenie (semenzali)
L’osservanza delle precauzioni di cui al punto precedente (utilizzo di genitori sani) può non
essere sempre possibile, in particolare quando ricorrano: (a) inderogabili esigenze del programma di
miglioramento che possono richiedere l’utilizzo di genotipi (sia come portaseme che come
produttori di polline) per i quali non esistono fonti sane; (b) impossibilità di collocare i campi di
selezione a distanza di sicurezza da eventuali fonti d’infezione rappresentate da coltivazioni
commerciali, situazione questa che facilmente si incontra nel nostro Paese per le specie di
maggiore importanza. Occorre inoltre tener presente la diffusa presenza di fruttiferi ornamentali in
parchi e giardini e di specie selvatiche affini in luoghi incolti.
Nel caso si debba necessariamente ricorrere per gli incroci a uno od entrambi i genitori
affetti da patogeni trasmissibili per polline o per seme, si può procedere alla determinazione dello
stato sanitario dei semenzali nei primi stadi vegetativi (dopo l’emissione della terza-quinta foglia)
con metodi sierologici (ELISA) o molecolari (PCR a singolo o doppio stadio) (Martelli, 1999) sì da
scartare immediatamente i soggetti infetti. È comunque norma prudenziale ripetere il saggio a
distanza di alcuni mesi per dare il tempo ai virus che eventualmente siano sfuggiti alla prima
analisi, di moltiplicarsi e raggiungere una concentrazione rilevabile ai saggi. Peraltro, anche se i
semenzali derivano da parentali allevati con ogni possibile precauzione e ritenuti sani, occorre
comunque controllarne lo stato sanitario nel campo di allevamento.
Motivi pratici legati al numero, spesso cospicuo, dei semenzali allevati, rendono
problematica l'adeguata osservazione e la valutazione sanitaria della totalità di essi, anche tenendo
presente che la maggior parte è destinata ad una vita piuttosto breve (da 3-5 anni nel pesco a 7-8
anni per susino europeo e melo). Elementari motivi di prudenza richiederebbero la sollecita
eliminazione dei soggetti non meritevoli di moltiplicazione, senza attendere la valutazione
dell’intero campo di selezione (cioè che tutti i semenzali entrino in produzione), in considerazione
del fatto che la loro messa a frutto non è contemporanea, a causa della eterogeneità che li rende
individui unici.
Controllo dello stato sanitario delle selezioni
Al momento della scelta del semenzale per la successiva fase di moltiplicazione (una volta
che questo sia stato giudicato interessante per un qualche aspetto), occorre mettere in pratica alcune
precauzioni. In primo luogo, osservazioni visive che escludano la presenza conclamata di malattie
virali e, parallelamente, l’esecuzione di saggi di laboratorio e biologici (indexaggio) che consentano
maggiore sicurezza nell’accertamento dello stato sanitario. Per la moltiplicazione del semenzale
selezionato si possono seguire strade diverse, a seconda della risposta ottenuta dalle analisi
diagnostiche:
(a) se i saggi di laboratorio sono negativi, si può procedere alla moltiplicazione del semenzale in
vivaio, in attesa dell’esito dell’indexaggio. Se questo è disponibile prima del risveglio vegetativo
che segue all’innesto, si potrà eliminare dal vivaio il materiale infetto. Nel caso però che i risultati
dell’indexaggio non siano noti prima del risveglio vegetativo della pianta innestata ed esistano
fondati sospetti sulla presenza di agenti infettivi trasmessi da afidi, è consigliabile la temporanea
collocazione in vaso delle piante moltiplicate, conservandole in strutture di quarantena a prova di
insetto;
(b) se i saggi di laboratorio sono positivi, la destinazione della selezione può dipendere dalla
pericolosità dell’agente infettivo. Se trattasi di virus oggetto di lotta obbligatoria (PPV delle
drupacee, CTV degli agrumi, ad esempio), il costitutore dovrà valutare l’opportunità di procedere
alla eliminazione della selezione, ovvero al suo risanamento prima dell’introduzione in pieno
campo; tale decisione dipenderà evidentemente dal rapporto tra costo previsto e valore pomologico
della selezione;
(c) nel caso in cui i saggi biologici e di laboratorio rivelino la presenza di agenti trasmissibili per
seme o per polline, ma non particolarmente gravi, occorrerà in primo luogo valutare l’opportunità di
procedere al risanamento prima di andare avanti con la selezione, nella consapevolezza che sarà
comunque necessario intervenire prima dell’eventuale licenziamento o cessione del materiale a
terzi.
(d) se anche l’indexaggio è negativo, i soggetti potranno essere collocati in pieno campo.
Avvertenze per la propagazione del materiale selezionato
Indipendentemente da quanto specificato al punto precedente, la propagazione del materiale
selezionato deve avvenire tenendo presente la necessità di: (a) usare portinnesti certificati (quando
la propagazione avviene per innesto), (b) collocare e mantenere in strutture protette da insetti e
nematodi almeno due piante sane per ogni genotipo, per tutta la durata della fase di selezione.
Occorre infatti tener presente che al termine delle fase selettiva (che spesso richiede l’esecuzione di
almeno due momenti successivi di selezione, e quindi di propagazione e di permanenza in campo),
trascorrono 8-10 anni, con conseguenti possibilità di reinfezione in campo. Tale situazione
porterebbe alla pratica impossibilità, al momento della diffusione commerciale, di poter distribuire
il materiale di moltiplicazione delle selezioni valide, a meno che non si sia provveduto al
mantenimento di alcuni soggetti in condizioni di garanzia di sanità fin dall’inizio del processo
selettivo.
Considerazioni conclusive
Il pericolo che nel corso delle attività di miglioramento genetico si possa incorrere in
problemi sanitari è pertanto attuale e da non prendere alla leggera. Nel passato, a causa delle
meno avanzate conoscenze sull'ecologia dei virus dei fruttiferi e dello scarso interesse mostrato dai
costitutori per gli aspetti sanitari, non vi è stato tra genetisti e virologi quel rapporto di
collaborazione che sembra oggi consolidarsi. Da una parte, la consapevolezza che le infezioni virali
sono diffuse e dannose (il caso della Sharka è emblematico) e che possono mettere a repentaglio
anni di attività di studio e, dall'altra, l'esistenza di provvedimenti legislativi (CAC, certificazione)
che impongono determinati standard sanitari, costituiscono spinte sufficientemente forti per la
ricerca di quelle sinergie atte a diminuire i pericoli, se non ad annularli del tutto. Si dispone oggi di
metodiche diagnostiche sufficientemente sensibili ed affidabili (la cui continua evoluzione fa
prevedere ulteriori affinamenti e semplificazioni applicative) che hanno già grandemente giovato
al vivaismo frutticolo (Martelli, 1999) e permettono il monitoraggio puntuale e continuo del
materiale vegetale nelle varie fasi del processo migliorativo. Poiché anche nel nostro Paese
esistono le condizioni a che si instaurino più stretti legami tra settori disciplinari la cui
complementarietà è indubbia perché non farne tesoro? Se gli esempi già in atto faranno scuola, ci si
avvia verso un futuro scientificamente più roseo.
BIBLIOGRAFIA
Barba M., Bassi D., Catara A., Tribulato E. 1999. Produzione di nuove accessioni nel rispetto degli
standard richiesti dalle normative per la commercializzazione e certificazione del materiale
vivaistico. Atti del Convegno Nazionale Certificazione delle produzioni vivaistiche,
Locorotondo e Bari, 1999: 68-85.
D’Onghia A.M., Djelouah K., Savino V. 2000. Serological detection of Citrus psorosis virus in
seeds but not in seedlings of infected mandarin and sour orange. Journal of Plant Pathology
82: 233-235.
Desvignes J.C. 1999. Maladies à Virus des Arbres Fruitiers. Editions Centre Technique
Interprofessionel des Fruits et Légumes, Paris, pp 202.
Jakubczyk H., Lata B., Sadowski A., Whitehead P. 1997. Virus free plant material as a basis for
integrated and quality systems in fruit production. International Seminar on Ecological aspects
of nutrition and alternatives for herbicides in horticulture, Varsavia, 1997. Rozwoj SGGWAR, Fundacja: 38.
Johansen E., Edwards M.C., Hampton R.O. 1994. Seed transmission of viruses: Current
perpectives. Annual Review of Phytopathology, 32: 363-385
Lankes, C. 2000. The new certification scheme and its consequences for the breeders. Acta
Horticulturae 538: 681-683.
Martelli G.P. 1999. La diagnosi virologica al servizio del vivaismo frutticolo. Frutticoltura 61 (4):
85-88.
Maury Y., Buby C., Khetarpal R.K. 1998. Seed certification for viruses. In: Plant Virus Diseases
Control. A. Hadidi, R.K. Kheterpal, H. Koganezawa (Eds.), 237-248. American
Phytopathological Socieity Press, St. Paul, USA.
Mink G.I. 1993. Pollen- and seed-transmitted viruses and viroids. Annual Review of Phytopathology
31: 375-402
Myrta, A., Di Terlizzi B., Savino V. 1998. Study on the transmission of plum pox potyvirus through
seeds. Phytopathologia Mediterranea 37:41-44.
Pasquini G., Simeoni A. M., Conte L., Barba M. 2000. RT-PCR evidence of the non-transmission
through seeds of Plum pox virus strains D and M. Journal of Plant Pathology 82: 221-226.
Stace-Smith R,. Hamilton R.I., 1988. Inoculum thresholds of seedborne pathogens. Viruses
Phytopathology 78: 875-880.
Vicchi V., Babini A. R., Buscaroli C., Rizzo M. 1997. Miglioramento genetico e selezione sanitaria:
binomio vincente per il vivaismo del futuro. Rivista di Frutticoltura, 10: 19-24.
Wan Chow Wa Y.F., Symons R.H., 1999. Transmission of viroids via grape seeds. Journal of
Phytopathology 147: 285-291.
Interventi di:
P. Caggiano, vivaista orticolo, Cooperativa Vivaistica Meridionale, Battipaglia (Sa)
M. Leis, vivaista frutticolo, Consigliere del C. d. A. Consorzio Italiano Vivaisti, Ferrara
G. Merisio, Ente Nazionale Sementi Elette, Milano
I. Ponti, Servizio Fitosanitario Regione Emilia Romagna, Bologna
S. Sansavini, Dipartimento di Colture Arboree, Università degli Studi di Bologna
F. De Corato, rappresentante Associazione Italiana Sementi, Andria (Ba)