27 APRILE
Prova in itinere
La cellula introduzione
Macromolecole: DNA e cromatina
Proteine
DNA replicazione
Trascrizione
Traduzione
Controllo dell’epressione genica
Citoscheletro
Organelli
Membrana plasmatica e trasporto
15 domande in 45 min (3 min/domanda)
TUTTI QUESTI
PROCESSI
NECESSITANO DI
ENERGIA
NEI MECCANISMI 1 E 3
LA PROTEINA
MANTIENE LA SUA
CONFORMAZIONE
Traslocatori proteici situati
nella membrana
NEL MECCANISMO 2 LA
PROTEINA DEVE
DISTENDERSI
Viaggiano a bordo di vescicole di
trasporto per andare dal RE
all’Apparato di Golgi, endosomi,
lisosomi e superficie cellulare
Esocitosi costitutiva ed esocitosi regolata (ormoni, muco , enzimi digestivi)
Per arrivare ai lisosomi i materiali destinati alla degradazione fanno percorsi diversi
Fagocitosi
Pinocitosi
Autofagia (degradazione di parti consumate della cellula)
1. La cellula deve saper rispondere a sollecitazioni dell’ambiente (sensori)
2. Le sostanze alimentari devono entrare i prodotti di rifiuto uscire (canali)
3.
La membrana cresce se la cellule cresce (si estende aggiungendo
nuova membrana), la membrana si può deformare senza strapparsi
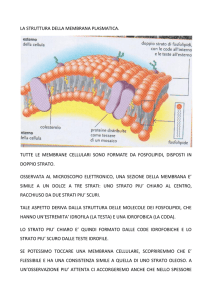
Struttura della Membrana
•
Le membrane cellulari consistono in un doppio strato
continuo di molecole lipidiche in cui sono immerse le
proteine.
•
Le molecole lipidiche della membrana presentano sia
regioni idrofiliche sia regioni idrofobiche.
Molecole anfipatiche (idrofobiche, idrofiliche)
in azzurro testa idrofilica
Galattosio
fosfatidil
Fosfatidilcolina
La molecola della fosfatidilcolina è il fosfolipide più comune nella membrana cellulare, è costituita da cinque
parti: i. colina, ii. fosfato (H3PO4, Pi, ione), iii. glicerolo, iv. coda idrocarburica (acido grasso insaturo/saturo),
v. coda idrocarburica (i-iii, testa idrofilica, iv-v, coda idrofobica)
Molecola idrofilica
stabiliscono legami IDROGENO con le molecole di acqua
Molecola idrofobica
non stabiliscono legami con le molecole di acqua
Le molecole d’acqua formano legami idrogeno tra loro e racchiudono la
molecola idrofoba apolare in una sorta di gabbia
Le molecole dei grassi sono idrofobiche
Le molecole fosfolipidiche sono anfipatiche (idrofobiche, idrofiliche)
in azzurro testa idrofilica
•
Poste in soluzioni acquose, si associano
spontaneamente in doppi strati e formano comparti
chiusi capaci di autosigillarsi se lacerati.
La forma sferica è stabile perché evita alle code idrocarburiche idrofobiche
di rimanere esposte all’acqua
Essenziale per la sopravvivenza della cellula
Meccanismo tale per cui la membrana si ripara (buchi) quando si rompe
La membrana cellulare è flessibile
si piega: 25 nm dimensione minima delle vescicole
che può racchiudere
Meno di una
volta al mese
per molecola
lipidica
La membrana cellulare è fluida
i fosfolipidi si spostano e si scambiano di posto con gli altri
PIU’ FLUIDA SE:
1- catene acidi grassi sono più corte, <14-24 atomi di Carbonio
2- se le catene di acidi grassi sono INSATURI (più doppi legami, maggiore capacità
di legare H e di formare anse laterali)
3- meno colesterolo
La membrana cellulare è asimmetrica
Colina e sfingomielina
Serina e etanolammina e fosfatidilinositolo
• Il doppio strato lipidico è asimmetrico, i fosfolipidi ed i glicolipidi si
distribuiscono in modo asimmetrico
• Fasfatidilcolina (rosso), sfingomielina (marrone), fasfatidilserina
(verde chiaro), fasfatidilinositolo (verde scuro), fasfatidiletanolamina
(giallo)
• Tutte le molecole glicolipidiche (azzurri) si trovano nel monostrato
esterno della membrana, il colesterolo (grigio) si distribuisce
equamente su entrambi i lati
Faccia citosolica della membrana
Faccia non citosolica della membrana
Le vescicole membranose hanno origine per gemmazione e fusione dagli organelli
citoplasmatici (es. Golgi) e mantengono una polarità (i glicolipidi sono nella metà della
membrana non rivolta al citosol)
Classe funzionale
Esempio di proteina
Funzione specifica
Vettori
Pompa Na+
Pompa attivamente Na+ fuori dalla cellula e K+
dentro la cellula
Connettori molecole
di adesione
Integrine
Collegano i filamenti intracellulari di actina a
proteine della matrice extracellulare
Recettori
Recettore del fattore di
crescita piastrinico
(PDGF)
Lega il PDGF extracellulare e d conseguenza
genera segnali intracellulari che inducono la
cellula a crescere e dividersi
Enzimi
Adenilato ciclasi
Catalizza la produzione di AMP ciclico
intracellulare in risposta a segnali extracellulari
•
Le proteine transmembrana si estendono attraverso il
doppio strato lipidico, generalmente assumendo una
conformazione ad elica α singola o multipla, ma talvolta
anche come piano β avvolto a manicotto.
A. B. C. Proteine integrali di membrana
Regioni idrofiliche e
regioni idrofobiche
Elica α anfipatica
Legame
covalente con
molecola lipidica
(zig zag rosso)
D. Proteine
periferiche di
membrana
Legame non
covalente con
altre proteine
(A) Proteine Transmembrana
(A) Proteine Transmembrana
(A) Proteine Transmembrana
Le catene laterali degli aminoacidi idrofobici (verde) da un lato di ogni elica entrano in
contatto con le code idrocarburiche che pure sono idrofobiche, mentre le catene laterali
idrofiliche (rosso) al lato opposto formano un pro pieno di acqua
La membrana plasmatica è di per se stessa fragile e sottile
(10000 membrane una sopra l’altra per formare lo spessore di una pagina di libro)
CITOSOL
Tutte le membrana plasmatica cellulari sono rinforzate e sostenute da un impalcatura
proteica fissata per mezzo di proteine transmembrana.
Tali proteine si organizzano in un trama di proteine fibrose (lo strato corticale o cortex
cellulare) aderente alla faccia citosolica che determina la forma cellulare e e le proprietà
meccaniche della membrana plasmatica
Nel globulo rosso lo strato corticale è formato soprattutto da spettrina
quando è geneticamente anomala gli eritrociti sono fragili e si ha l’anemia
• Dato che la membrana si comporta come un fluido bidimensionale, i suoi lipidi e
anche molte delle sue proteine possono muoversi liberamente nel piano del doppio
strato lipidico.
• La cellula sa però confinare certe proteine di membrana in zone circoscritte in distretti
funzionali denominati domini di membrana; la mobilità laterale delle proteine nella
membrana plasmatica subisce quindi limitazioni di vario genere
• Le proteine possono venire ancorate allo strato corticale interno (A), a molecole della
matrice extracellulare fuori dalla cellula (B), a proteine situate sulla superficie di
un’altra cellula.
• Possono esistere delle barriere alla diffusione (barrette nere) che confinano certe
proteine in un particolare dominio di membrana.
• Nelle cellule intestinali le proteine sono confinate in un particolare dominio di membrana
plasmatica.
• La proteina A (proteine di trasporto attive nell’assumere sostanze nutritive) e la proteina B
(trasporto di soluti fuori dalle cellule epiteliali verso il sangue ed i tessuti) possono diffondere
lateralmente nel dominio di membrana cui appartengono ma non nell’altro a causa di un
particolare tipo di connessione con le cellule adiacenti.
• Le giunzioni occludenti sigillano i vari comparti di membrana (proteine apposite formano una
cintura ininterrotta attorno alla cellula a stabiliscono della saldature).
La struttura delle membrane
•
•
•
•
Molte delle proteine e alcuni dei lipidi esposti sulla superficie
esterna della cellula portano attaccate catene di zuccheri, che
contribuiscono a proteggere e lubrificare la superficie cellulare e
sono attivi nel riconoscimento tra cellule.
Quasi tutte le membrane cellulari sono attaccate a proteine che
fanno da impalcatura di sostegno.
Ne è un esempio la rete intessuta da proteine fibrose che forma il
cortex, strato corticale posto proprio sotto la membrana cellulare.
Anche se molte proteine diffondono rapidamente nel piano della
membrana, le cellule hanno modi per confinarle in domini circoscritti
e per immobilizzarne certe, attaccandole alle macromolecole
intracellulari o extracellulari.
• Le cellule eucariotiche sono rivestite da glucidi che formano il glicocalice
• Il glicocalice (strato a carboidrati) è costituito da catene laterali
oligosaccaridiche attaccate a glicolipidi e alle glicoproteine di membrana
• Al glicocalice possono contribuire anche le glicoproteine e i proteoglicani
secreti dalla cellula e quindi riassorbiti subito dalla cellula
• Tutti i carboidrati si trovano dal lato extracellulare
• Aiutano a proteggere e lubrificare la superficie dal danneggiamento
meccanico (assorbendo acqua gli oligo ed i polisaccardi rendono scivolosa
la superficie)
• Il glicocalice non solo aiuta a proteggere e lubrificare la superficie dal danneggiamento meccanico
ma funge anche da segnale di riconoscimento e nell’adesione cellulare
• Le lectine sono specializzate nel riconoscere particolari catene oligosaccaridiche e legarle con gli
stessi meccanismi e legami di riconoscimento che regolano l’interazione tra le proteine
• Le catene laterali oligosaccaridiche delle glicoproteine
• Mentre le proteine si uniscono tutte linearmente con legami identici gli zuccheri possono farlo in
vario modo in varie sequenze e anche in catene ramificate
• La diversificazione dei legami fa si che il glicocalice funga da divisa di riconoscimento per la
cellula (uovo spermatozoo)
Trasporto di Membrana
I doppi strati lipidici sono impermeabili le membrane plasmatiche no
La membrana plasmatica
•
Lo strato lipidico delle membrane cellulari è permeabile alle molecole
piccole e apolari, come l’ossigeno e il diossido di carbonio, ma anche a
molecole polari purché piccolissime, come l’acqua (diffusione semplice vs.
diffusione facilitata, e.g. acquaporine).
•
E’ invece impermeabilissimo a quasi tutte le molecole idrosolubili e a tutti gli
ioni.
•
Il trasferimento di sostanze nutritive, metaboliti e ioni attraverso la
membrana plasmatici e le membrane intracellulari è demandato alle
proteine di trasporto che esse contengono,
•
Le membrane cellulari contengono tutta una serie di proteine di trasporto,
ciascuna incaricata del trasferimento di un particolare tipo di soluto
attraverso la membrana.
•
Le proteine di trasporto si possono distinguere in due categorie: le proteine
vettore e le proteine canale.
• Una molecola diffonderà
tanto più rapidamente
attraverso il doppio
strato lipidico quanto più
è piccola e soprattutto
quanto meno interagisce
favorevolmente con
l’acqua (cioè quanto
minore è la sua polarità)
• Molte delle molecole
che servono alla cellula
come nutrimento sono
troppo grandi e polari
per superare un doppio
strato lipidico puro
La cellula deve contenere quantità uguali di cariche positive e negative
(cioè deve essere elettricamente neutra)
Componente
Concentrazione
intracellulare (mM)
Concentrazione
extracellulare (mM)
Na+
5-15
145
K+
140
5
Mg2+ (ioni liberi)
0,5 (20 mM se incluso quello
complessato a proteine)
1-2
Ca2+ (ioni liberi)
contenuto in organelli
10-4 (1-2 mM se incluso quello
complessato a proteine)
1-2
H+
7x10-5 (10-7,2 M o pH 7,2)
4x10-5 (10-7,4 M o pH 7,4)
5-15
110
Cationi
Anioni
ClProteine, acidi nucleici, metaboliti con
gruppi fosfato e carbossilici, HCO-3,
PO43-
• Anche se le cariche elettriche all’esterno e all’interno sono
bilanciate e in equilibrio, intorno alla membrana plasmatica si
rilevano eccessi contenuti di carica netta positiva o negativa
• Questi squilibri generano una differenza di voltaggio detta
POTENZIALE di MEMBRANA compreso tra -20 e -200 mV
• Il potenziale è negativo perchè l’interno della cellula ha una carica
netta negativa rispetto all’esterno
• Questo potenziale è alla base di alcuni meccanismi di trasporto e
della generazione dell’impulso nervoso
Le molecole di piccole dimensioni entrano nella cellule
attraverso un vettore o un canale
• Una proteina vettore va incontro ad una
serie di modificazioni conformazionali
per trasferire piccole molecole
idrosolubili attraverso il doppio strato
lipidico
• La proteina canale forma un poro
idrofilico che trapassa il doppio strato
attraverso cui specifici ioni inorganici
possono diffondere
• Le proteine canale sono più rapide
nel trasporto delle proteine vettore
• I canali ionici esistono in
conformazione chiusa e aperta, e
trasportano solo in quella aperta, la
loro chiusura o apertura è di solito
controllata da uno stimolo esterno o
da condizioni interne della cellula
TRASPORTO PASSIVO
• Proteine canali
• Proteine vettori=trasportatori
Gradiente chimico o di concentrazione
Gradiente elettrochimico
TRASPORTO ATTIVO
• Proteine vettori=trasportatori
Accoppiato
ATP-dipendente
FOTO dipendente
Un soluto che abbia una concentrazione più alta fuori dalla cellula entrerà
spontaneamente dentro la cellula per trasporto passivo (diffusione facilitata)
purché abbia nella membrana un canale od un vettore apposito
Contro gradiente ci vogliono proteine vettore in grado di
attingere energia per alimentare il passaggio (trasporto attivo)
Ogni membrana cellulare ha la sua serie caratteristica di proteine vettore
TRASPORTO PASSIVO: gradiente di concentrazione
Un cambiamento di conformazione di una proteina vettore
potrebbe mediare il trasporto passivo di un soluto come il glucosio
La transizione tra i due stati avviene a caso,
è indipendente dal soluto e reversibile
TRASPORTO PASSIVO: gradiente elettrochimico
La forza motrice netta (gradiente elettrochimico) che tende a
spostare un soluto carico (ione) attraverso la membrana è la
somma del gradiente di concentrazione e della differenza di
potenziale tra i due lati della membrana (potenziale di membrana)
attira i soluti
carichi
positivamente
attira i soluti
carichi
negativamente
L’ampiezza della freccia indica l’entità del gradiente
Situazione usuale lato citoplasmatico carico negativamente
il Na+ che è concentrato fuori dalla cellula quando può cercherà di entrare nella cellula
Il K+ che è più concentrato all’interno tenderebbe ad uscire secondo gradiente di concentrazione ma essendo il gradiente
elettrochimico modesto il K tende a non uscire dalla cellula
Il trasporto attivo
Trasportatore accoppiato: accoppiano il trasporto di un soluto contro gradiente
al trasporto di un soluto secondo gradiente
Le pompe ad ATP: accoppiano il trasporto contro gradiente all’idrolisi di ATP
Le pompe fotoalimentate: accoppiano il trasporto contro gradiente
all’assorbimento di energia luminosa
Trasporto accoppiato
Le cellule animali utilizzano il gradiente di Na+ per
assumere attivamente sostanze nutritive
La pompa sodio potassio svolge un ruolo essenziale nelle cellule animali
Assorbe il 30% del consumo totale di ATP
Agisce come pompa di sentina per espellere Na+ dalla cellula che rientra attraverso proteine vettrici e
canali ionici e così facendo mantiene concentrazioni citosoliche diverse rispetto ai fluidi extracellulari
(10-30 volte inferiore Na+ e 10-30 volte superiore K+)
La pompa Na+-K+ contribuisce a mantenere l’equilibrio osmotico nella cellula animale
La membrana è permeabile all’acqua per mantenere costante la concentrazione di soluti va da zone con soluti poco
concentrati (e acqua molto concentrata) a zone con soluti molto concentrati (e poca acqua) OSMOSI
fosforilazione della pompa
cambio conformazionale
Grazie al trasporto attivo fatto dalle pompe le concentrazioni ioniche ai due lati
della membrane sono lontane dall’equilibrio
Alcuni esempi di proteine vettore
Proteina vettore
Collocazione
Fonte di energia
Funzione
Vettore del glucosio
Membrana plasmatica di quasi tutte le
cellule animali
Nessuna
Importazione passiva di
glucosio
Pompa del glucosio Na+dipendente
Membrana plasmatica apicale delle
cellule intestinali e renali
Gradiente di Na+
importazione attiva di
glucosio
Scambiatore Na+-H+
Membrana plasmatica delle cellule
animali
Gradiente di Na+
Esportazione attiva ioni H+
regolazione del Ph
Pompa Na+-K+ (Na+-K+-ATPasi)
Membrana plasmatica di quasi tutte le
cellule animali
Idrolisi di ATP
Esportazione attiva di Na+ e
importazione attiva di K+
Pompa del Ca2+ (Ca2+-ATPasi)
Membrana plasmatica delle cellule
eucariotiche
Idrolisi di ATP
Esportazione attiva di Ca2+
Pompa protonica (H+-ATPasi)
Membrana plasmatica delle cellule
vegetali, fungine e di alcuni batteri
Idrolisi di ATP
Esportazione attiva di H+
dalla cellula
Pompa protonica (H+-ATPasi)
Membrane lisosomiali delle cellule
animali e membrane vacuolari delle
cellule vegetali e fungine
Idrolisi di ATP
Esportazione attiva di H+ dal
citosol nel vacuolo
Batteriorodopsina
Membrana plasmatica di alcuni batteri
Luce
Esportazione attiva di H+
fuori dalla cellula