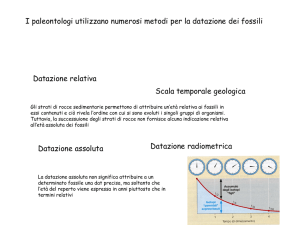
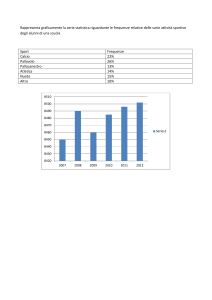
Variabilità genetica
La variabiltà della struttura genetica
esistente nelle popolazioni è superiore a
quella riconducibile alle differenze
fenotipiche. Essa può essere misurata
analizzando i polimorfismi della sequenza
di DNA e individuando i diversi genotipi.
Polimorfismo: proprietà di un gene di avere 2 o più alleli con frequenze
maggiori dell’ 1%.
Nella specie umana due individui differiscono in media per 3 milioni di
nucleotidi (0.1 % del genoma)
Polimorfismi e Marcatori genetici
Tipo di marcatore
N° di loci
Caratteristiche
Gruppi sanguigni
1910-1960
Circa 20
Genotipo non facilmente desumibile.
Posizioni fisiche dei geni non note.
Varianti mobilità
elettroforetica
proteine sieriche
1960-1975
Circa 30
Posizioni fisiche dei geni non note.
Polimorfismo limitato
Tipi HLA
1970-
1
Un solo gruppo di associazione
RFLP del DNA
1975-
> 105
potenzialmente
2 alleli/locus. Southern o PCR. Facile
localizzazione fisica
VNTR del DNA
(minisatelliti)
1985-
> 104
potenzialmente
Molti alleli/locus. Southern. Più
frequenti alle estremità dei cromosomi
VNTR del DNA
(microsatelliti)
1989-
> 105
potenzialmente
Molti alleli/locus. PCR automatizzata.
Tutto il genoma.
SNPs del DNA
(minisatelliti)
1998-
> 4.106
Arrays
GENETICA DI POPOLAZIONI
La genetica di popolazione studia la composizione dei
geni all’interno di un gruppo di individui cioè le diverse
forme alleliche e i diversi genotipi, le variazioni che
interessano questi geni nel tempo e le differenze
esistenti tra vari gruppi o popolazioni
Frequenze genotipiche
e frequenze alleliche di una popolazione
In un sistema a due alleli, A e a:
f(AA)=
Num. di individui AA
f(aa)=
Num. di individui aa
N
N
f(Aa)=
Num. di individui Aa
N
p =f(A)=
2 x AA + Aa
2N
q =f(a)=
2 x aa + Aa
2N
Frequenze alleliche di un locus legato all’X
In un sistema a due alleli, XA e Xa:
p =f(XA)=
2 XAXA + XAa + XAY
2xfemm.+1xmaschi
q =f(Xa)=
2 XaXa + XAa + XaY
2xfemm.+1xmaschi
FREQUENZE GENOTIPICHE GRUPPO SANGUIGNO MN
Population
M/M
M/N
N/N
Eskimo
0.835
0.156
0.009
Egyptian
0.278
0.489
0.233
Chinese
Australian
aborigine
0.332
0.486
0.182
0.024
0.304
0.672
Stratificazione della popolazione umana in
Gruppi Etnici
Effetto della riproduzione sessuale sulla variabilità.
L’equilibrio di Hardy-Weinberg
In una popolazione ampia, caratterizzata da accoppiamento casuale, ogni genotipo
si accoppia in modo proporzionale alla sua frequenza. Quindi, le frequenze dei tipi di
gameti A e a nella popolazione saranno p e q.
A (p)
a (q)
A (p)
a (q)
A/A
(p2)
A/a
(pq)
A/a
(pq)
a/a
(q2)
Quindi, dopo un ciclo riproduttivo, le frequenze genotipiche nella popolazione saranno
p2 per A/A, 2pq per A/a, and q2 per a/a.
Quindi: p2 + 2pq + q2 = 1
Legge di Hardy-Weinberg
•
•
Condizioni necessarie:
1. popolazione infinitamente grande.
2. accoppiamento casuale
3. assenza di mutazione, migrazione e
selezione naturale
Enunciato:
Se queste condizioni sono rispettate le
frequenze genotipiche non variano nel
corso delle generazioni e dipendono dalle
frequenze alleliche secondo l’equazione
p2 + 2pq + q2 = 1
Frequenze genotipiche di un locus legato all’X
In equilibrio di Hardy-Weinberg:
femmine
maschi
f(XAXA)= p2
f(XAY)= p
f(XaXa)= q2
f(XaY)= q
f(XAXa)= 2pq
Relazioni tra i genotipi e le frequenze alleliche
Applicazione della legge
1. Verifica delle proporzioni di Hardy-Weinberg
Locus Perossidasi degli alberi di Pino Ponderosa del Colorado
Genotipi
R2R2
R2R3
R3R3
Valori osservati
135
44
11
Sono in equilibrio ?
Calcoliamo dapprima le frequenze alleliche
p =f(R2)=
2 x R2R2 + R2R3
2N
q =f(R3)=
2 x R3R3 + R2R3
2N
=
2(135) +44
2(190)
= 0.174
= 0.826
p+q=1
poi calcoliamo le frequenze genotipiche attese dall’equilibrio di
Hardy-Weinberg:
R2R2=p2=(0.826)2=0.683
R2R3=2pq=2(0.826)(0.174)=0.287
R3R3=q2=(0.174)2=0.03
moltiplicando queste frequenze attese per il numero totale di
individui N si ottiene il valore numerico atteso per ogni genotipo:
Valori attesi
Valori osservati
R2R2=0.683 x 190 = 129.8
135
R2R3=0.287 x 190 = 54.5
44
R3R3=0.03 x 190 = 5.7
11
Conclusione: è altamente improbabile che i genotipi siano in
equilibrio di Hardy-Weinberg
Esempi di popolazioni in equilibrio
FREQUENZE GENOTIPICHE
OSSERVATE
FREQUENZE GENOTIPICHE
ATTESE
Population
M/M
M/N
N/N
M/M
M/N
N/N
Eskimo
0.835
0.156
0.009
0.834
0.159
0.008
Egyptian
0.278
0.489
0.233
0.274
0.499
0.228
Chinese
0.332
0.486
0.182
0.331
0.488
0.181
0.024
0.304
0.672
0.031
0.290
0.679
Australian
aborigine
2. Stima delle frequenze alleliche per mezzo della legge di HardyWeinberg.
Es. Carattere autosomico recessivo: Fibrosi cistica
Incidenza della malattia 1/2000
quindi q2 = 0.0005
q=
0.0005 = 0.02
p = 1-0.02= 0.98
f(Aa)= 2pq = 2 x 0.98 x 0.02 = 0.0392
4% portatori allele fibrosi cistica nella popolazione caucasica
Fattori che alterano le frequenze alleliche
•
•
•
•
•
Mutazione
Migrazione
Accoppiamenti non casuali
Selezione
Eventi casuali che si verificano nelle
piccole popolazioni (deriva genetica)
La variabilità derivante da mutazione
I tassi di mutazione spontanea sono molto bassi, tanto
che la mutazione da sola (in assenza di selezione o
deriva genetica) non può spiegare l’ evoluzione delle
popolazioni e delle specie
Point-Mutation Rates in Different Organisms
Organism
Gene
Bacteriophage
Host range
Escherichia coli
Phage resistance
Zea mays (corn)
R (color factor)
Y (yellow seeds)
Drosophila melanogaster
Average lethal
Mutation rate per
generation
9
2.5 × 10
2 × 10
8
4
2.9 × 10
2 × 10
2.6 × 10
6
5
L’azione esercitata dalla mutazione è lenta
µ
sono necessarie 70000 generazioni per dimezzare la frequenza di p
La variabilità derivante da migrazione
La migrazione causa:
• aumento delle dimensioni di una
popolazione
• riduce la divergenza tra popolazioni
diverse
• si oppone agli effetti della deriva genetica
La selezione naturale
•Selezione naturale: sopravvivenza e riproduzione
differenziale dei genotipi caratterizzati da fitness diversa
•Fitness darwiniana: valore che indica la probabilità
relativa di sopravvivenza e di riproduzione media di un
genotipo in un determinato ambiente
Teoria dell’evoluzione (C. Darwin)
1. Principio della variabilità
2. Principio dell’ereditarietà
3. Principio della selezione
C. Darwin
Effetto della selezione di una mutazione
vantaggiosa sulla frequenza allelica
Esempio di selezione naturale e ruolo
dell’ambiente
Polimorfismo bilanciato (vantaggio
dell’eterozigote)
Si verifica quando la fitness dell’eterozigote non è
intermedia ma più estrema rispetto a quelle degli
omozigoti. In questo caso la selezione non porta alla
scomparsa di un allele ma tende a raggiungere una
frequenza allelica intermedia
Anemia falciforme e malaria
Distribuzione della malaria
Distribuzione dell’allele HbS
Un esempio di vantaggio dell’eterozigote è quello che ha causato
l’espansione dell’anemia falciforme nelle zone in cui la malaria è
endemica
Deriva genetica. Variazione delle frequenze
alleliche dovuta al caso
Simulazione di Deriva genetica
La Deriva genetica si osserva nelle popolazioni di piccole dimensioni
E’ dovuta ad un errore di campionamento
La deriva genetica causa:
• La perdita di variabilità genetica all’interno
delle popolazioni
• la divergenza tra popolazioni diverse
• la fluttuazione delle frequenze alleliche di
una popolazione nel tempo
Effetto del fondatore
Isola di Tristan da Cuna
Colonizzata da W. Glass nel 1817
Arrivo di marinai scampati a naufragio e donne trasferite da S. Elena
Pololazione di 100 abitanti nel 1855
1856, Morte di Glass. Emigrazione e riduzione popolazione a 33 abitanti
1885, dopo espansione, nuova riduzione per morte di 15 uomini ed
emigrazione delle vedove
Attualmente, 300 abitanti. Elevata incidenza di asma.
Effetto “collo di bottiglia”
Incroci non casuali
•Inbreeding o Inincrocio: accoppiamento tra
consanguinei
•Esoincrocio o Esogamia: accoppiamento
preferenzialmente tra non consanguinei
L’inincrocio aumenta l’omozigosità.
L’esogamia aumenta l’eterozigosità.
Riassunto degli effetti delle forze evolutive
sulla variabilità genetica
Esercizio
La lana Bianca dipende da un allele B dominante e la lana nera dal
suo allele recessivo b. Si supponga che un campione di 900 pecore
abbia dato i seguenti risultati: 891 pecore con lana bianca e 9 pecore
con lana nera. Si calcolino le frequenze degli alleli B e b e quelle dei
tre genotipi BB, Bb e bb nella popolazione, supponendo che sia
rispettato l’equilibrio di Hardy-Weinberg.