IL MUSCOLO E I MECCANISMI DI PRODUZIONE ENERGETICA
1. Il muscolo
di Roberto Biella
Il muscolo è un organo che, mediante complesse reazioni elettrochimiche, produce l’energia meccanica
necessaria per realizzare i movimenti dei segmenti corporei.
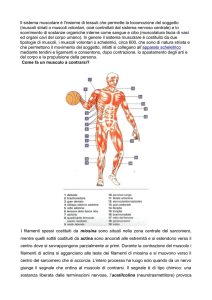
Il muscolo è costituito da:
Uno o più ventri muscolari (parte carnosa contrattile)
due o più parti tendinee che collegano la parte carnosa alle ossa (parte connettivale)
Tendine
Tendine
Ventre
Il tessuto muscolare si caratterizza per la capacità contrattile cioè di accorciarsi ed esercitare forza sotto
stimolazione nervosa proveniente dal cervello.
Il nome di alcuni muscoli dipende dalla loro forma oltre che dal punto in cui sono situati nel nostro corpo. Il
muscolo esemplificato sopra è un classico muscolo fusiforme. Se il ventre muscolare si divide in due,
originando due tendini si avrà un muscolo bicipite (due ventri e tre tendini in totale). Esistono muscoli
tricipiti (polpaccio e braccio) e quadricipiti (coscia). Esistono anche muscoli piatti (addominali retti e obliqui)
e anulari (sfinterici), a ventaglio (pettorale, gran dorsale, deltoide).
Esistono diversi modi di inserzione della parte tendinea sulle ossa. Se il tendine è a sezione cilindrica assume
l’aspetto di un cordone (tendini della mano e del polso), se invece è piatto si dice nastriforme (es. quello
del quadricipite sotto la rotula) ma può essere piatto ma molto esteso; in questo caso si dice aponeurosi
(es. l’inserzione sullo sterno e sulla clavicola del muscolo pettorale).
Il ventre muscolare è costituito da cellule altamente specializzate (fibre muscolari.) e da tessuto connettivo
che avvolgono le varie suddivisioni strutturali (intero muscolo, fasci muscolari, fibre muscolari). In questa
componente connettivale sono presenti le ramificazioni dell’apparato circolatorio e del sistema nervoso.
La fibra muscolare ha dimensioni variabili: spessore che varia da pochi micron fino a 150 micron (circa la
dimensione di un capello); lunghezza da pochi mm a diversi cm.(fino a 30cm. nel sartorio)
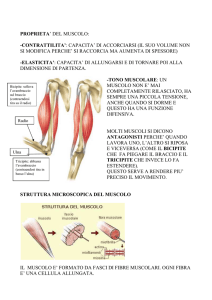
Caratteristiche del muscolo sono la:
TONICITA’ (leggero stato di contrazione sempre presente nel muscolo che permette di mantenere
ad es. una giusta postura)
CONTRATTILITA’ (cioè la capacità di variare la propria dimensione)
ELASTICITA’ (il muscolo si comporta come un elastico – restituisce della forza dopo essersi
allungato purchè sia sempre contratto).
Esistono 3 tipi di contrazione:
1) CONCENTRICA
2) ECCENTRICA
3) ISOMETRICA/STATICA
1) CONCENTRICA: si ha un accorciamento della fibra muscolare, quindi un avvicinamento dei due capi
ossei.
2) ECCENTRICA: c’è contrazione, ma vi è un allontanamento progressivo dei capi ossei o comunque un
cedimento sotto una forza esterna eccessiva (es. corsa in discesa, arresto, fase di ammortizzazione
nell’appoggio durante la corsa).
3) ISOMETRICA (se massimale, altrimenti STATICA): non vi è movimento esteriore perché la forza
applicata è pari a quella esterna (resistenza). Qui si può sviluppare il massimo della forza.
L’orientamento delle fibre rispetto alla direzione di trazione esercitata sui tendini-inserzioni, ha notevole
importanza in ordine di forza e ampiezza di accorciamento del muscolo stesso.
Le fibre muscolari sono disposte parallelamente o obliquamente rispetto all’asse longitudinale del muscolo
(angolo di pennazione).
Disposizione parallela (es. muscolo fusiforme) = maggior accorciamento , movimenti più ampi e veloci.
Disposizione obliqua (es. muscolo pennato) = minore accorciamento, ma più forza. Tale disposizione è
tipica dei muscoli pennati, bipennati e multipennati.
La forza del muscolo (P), a parità di diametro anatomico (d), è infatti proporzionale alla sezione trasversale
alla disposizione delle fibre (Sz).
La possibilità di accorciamento (A) del muscolo invece è proporzionale alla lunghezza (LF) delle fibre.
Confronto muscolo fusiforme vs penniforme:
d1=d2 (stessa dimensione)
Sz1<(Sz2a+Sz2b) quindi P1<P2
Lf1>Lf2 quindi A1>A2
Sz2a
Sz1
1
Sz2b
2
1
A1
2
A2
Nei muscoli del nostro corpo prevalgono le disposizioni oblique.
Muscolo striato scheletrico
Le fibre muscolari sono lunghe anche parecchi cm.,
con sezione di qualche decina di micron (20- 30).
Analizzando un muscolo dall’esterno verso l’interno
possiamo osservare le sottostrutture:
muscolo
fasci muscolari
fibre
miofibrille
miofilamenti.
La miofibrilla è composta da miofilamenti proteici di
actina e miosina disposti parallelamente e
longitudinalmente alla fibra. Tali filamenti si
dispongono alternatamente gli uni agli altri e in modo
che si sovrappongano parzialmente. In questo modo
ci sono delle zone in cui ci sono solo filamenti di
actina, altre solo quelle di miosina e altre ancora dove
ci sono entrambe. Questa particolare disposizione dei
due tipi di filamenti determinata la striatura
trasversale
caratteristica
della
muscolatura
scheletrica.
Infatti la fibra muscolare striata appare come una
successione di bande chiare e scure. L’UNITA’
FONDAMENTALE CONTRATTILE E’ IL SARCOMERO cioè quella zona compresa tra due linee Z.
Sapendo dove un muscolo si inserisce sulle ossa (sempre a
cavallo di una articolazione) e sapendo il movimento che può
compiere l’articolazione, è possibile comprendere quali
muscoli intervengono nel compiere certi esercizi.
SEMPRE IN RIFERIMENTO A UNA AZIONE O ALLA STATICA SU UNA ARTICOLAZIONE, un muscolo si dice:
AGONISTA nello svolgere un movimento di una
articolazione vs. l’ANTAGONISTA che fa compiere
il movimento in senso contrario (es. bicipite e
tricipite nella flessione e nella estensione
dell’avambraccio).
SINERGICO con un altro muscolo che svolge lo
stesso movimento.
FISSATORE che serve per bloccare un segmento osseo o una articolazione affinchè un’altra si possa
muovere (es. nei piegamenti a terra sulle braccia i muscoli addominali sono fissatori del tronco che
altrimenti si inarcherebbe).
E’ importante porre particolare attenzione a non confondere il lavoro cedente (negativo) a favore della
gravità come un movimento di muscoli antagonisti (es. movimento di
ritorno dell’abbassamento dell’avambraccio nel sollevamento di un
manubrio, movimento di piegamento delle gambe verso il basso). Il
movimento di andata (positivo in accorciamento) seguito dal
movimento di ritorno (negativo-cedente in allungamento a favore della
gravità) sono movimenti che prevedono la contrazione dello stesso
muscolo agonista con una contrazione concentrica (andata) e poi
eccentrica (ritorno).
2. La contrazione muscolare e i meccanismi di produzione energetica
Durante la contrazione i miofilamenti non modificano la
loro lunghezza, ma l’accorciamento del sarcomero è dovuto
esclusivamente ad uno scorrimento dei filamenti sottili negli
spazi esistenti tra quelli spessi, cioè nel muscolo contratto
si modifica esclusivamente il grado di sovrapposizione dei
filamenti stessi.
Lo scorrimento dei filamenti avviene per la creazione di
legami obliqui tra i filamenti. Questi legami, di tipo chimico
e catalizzati da ioni Calcio liberatisi per l'azione dell'impulso
nervoso, avvengono per interposizione di una molecola di
ATP
(adenosin-tri-fosfato).
Questi
legami
durano
pochissimi microsecondi dopodiché il legame si spezza e
l'ATP si scinde in ADP (adenosin-di-fosfato) e fosforo libero.
Per proseguire nella contrazione muscolare occorre
ricostruire il legame tra il fosforo perso e l'ADP che ridiventa
ATP e ciò richiede energia di tipo chimico proveniente da
un'altra parte (meccanismi di produzione energetica dal
metabolismo degli alimenti).
L'ATP è dunque l'unica sostanza che il muscolo
riconosce per poter contrarsi. L'ATP deve essere
considerata come una batteria carica; una volta
scaricata (ADP) non si butta ma deve essere
ricaricata prendendo dell'energia chimica da
un'altra fonte per ricostruire il legame altamente
energetico tra ADP e Fosforo che permette
“l’aggancio” obliquo tra filamenti di actina e
miosina e il relativo scorrimento gli uni sugli
altri.
L’energia chimica proveniente dal metabolismo
degli alimenti, una volta ricaricata l’ADP in ATP,
si trasforma durante la contrazione muscolare in lavoro meccanico + calore. Il lavoro meccanico (L) è dato
dalla forza esercitata dal muscolo (P) per il suo accorciamento (s).
Esistono 3 tipi di meccanismi di produzione di energia chimica per ricaricare l'ATP per il lavoro muscolare:
- MECCANISMO ANAEROBICO ALATTACIDO
- MECCANISMO ANAEROBICO LATTACIDO
- MECCANISMO AEROBICO
IL PROCESSO ANAEROBICO1 ALATTACIDO2 (CP + ATPR) è un meccanismo ad alta potenza ma che fornisce
una energia limitata pari a 8-10” circa di sforzo massimale, che quindi si esaurisce velocemente e non produce
nessuna sostanza dannosa per il muscolo. Tale meccanismo ricava l'energia dall'ATP di riserva e il CP
(creatin-fosfato o fosfocreatina) che si accumulano nel muscolo, previo allenamento, come riserve
energetiche di immediato utilizzo. L'ATP di riserva e il CP insieme formano il Pool dei Fosfati. E' il legame
altamente energetico del fosforo (sia dell'ATP che del creatinfosfato) che fornisce l'energia per ricaricare l'ADP
all'interno della cellula muscolare.
MECCANISMO ANAEROBICO ALATTACIDO
ATP
PC
ATPasi
ADP + P + Energia
P + C + Energia
fosfocreatinchinasi
POOL DEI
FOSFATI
IL PROCESSO ANAEROBICO LATTACIDO3 è un meccanismo tale per cui il glicogeno (che proviene dal
glucosio) si scinde in 2 molecole di piruvato e libera energia che ricarica l’ADP in ATP. Il piruvato (acido
piruvico) si trasforma rapidamente in acido lattico (AL) che è una sostanza tossica il cui accumulo nel muscolo
inibisce la contrazione muscolare. Ciò avviene a seguito di uno sforzo intensissimo e portato sino allo stremo
per alcune decine di secondi (o per scatti ripetuti a distanza di pochi secondi). L’acido lattico che si accumula
progressivamente nei muscoli che l’hanno prodotto, procura una specie di avvelenamento che noi avvertiamo
come appesantimento muscolare sino al forte rallentamento delle contrazioni muscolari, che ci costringe a
ridurre l’intensità dello sforzo e ad interrompere l’attività. Al termine dello sforzo, dopo qualche minuto (35 minuti) l’acido lattico passa dai muscoli al sangue e la sensazione di fatica diventa generale e intensa
soprattutto per i soggetti non allenati (con nausea, conati di vomito, mal di testa). Successivamente una parte
dell’acido lattico va al fegato dove viene trasformato in glicogeno e in parte ossidato da altri muscoli (tra cui
il cuore) specie quelli non particolarmente impegnati (es. quelli delle braccia durante la corsa).
La potenza (energia nell’unità di tempo) proveniente da questo meccanismo è piuttosto elevata ma non
molto duratura, consentendo sforzi intensi e massimali solo fino a 40- 60“ (es. gara dei 400 mt
dell’atletica).
L’acido lattico può accumularsi anche lentamente per sforzi di intensità sub massimale o medio alta protratti
sino allo stremo (es. gara di corsa campestre) o per scatti brevi ma ripetuti con poco recupero (es. partita di
pallacanestro o calcio).
MECCANISMO ANAEROBICO LATTACIDO
GLICOGENO (GLUCOSIO )
glicolisi anaerobica
AL(ac.lattico) + En
Il PROCESSO AEROBICO4 si avvale dell’ossigeno portato ai muscoli dal sistema respiro-cardio-circolatorio,
che per attivarsi impiega parecchi secondi per arrivare alla sua massima efficienza (dopo circa 2-3 minuti) e
di questo ne sono testimoni il nostro respiro e battito cardiaco che non si innalzano immediatamente
(inerzia di attivazione). Inoltre il lavoro dei muscoli respiratori e del cuore fa consumare ulteriore energia.
L’ossigeno brucia o meglio ossida e scinde i grassi e gli zuccheri liberando energia che ricarica l’ADP in
ATP. L’anidride carbonica prodotta viene portata attraverso il sangue ai polmoni ed eliminata attraverso
l’apparato respiratorio, l’acqua viene espulsa attraverso la sudorazione.
MECCANISMO AEROBICO
Zuccheri e grassi + O2
1
ciclo di Krebs
ANAEROBICO = senza ossigeno
ALATTACIDO = senza produzione di acido lattico
3
LATTACIDO = con produzione di acido lattico
4
AEROBICO = con intervento dell’ossigeno
2
CO2 + H2O + En
L'ossidazione degli zuccheri è tanto maggiore quanto più è intensa l'attività che si compie. Ad intensità
poco elevate si bruciano più grassi che zuccheri.
La potenza del meccanismo aerobico non è molto elevata ma molto duratura in quanto il corpo dispone di
ampie riserve di glicogeno (che è uno zucchero) nei muscoli e nel fegato e di grassi (panicoli adiposi).
La potenza del meccanismo aerobico, e quindi la capacità di prestazione negli esercizi fisici di resistenza
(es. corsa di fondo), è condizionata dalla disponibilità di ossigeno ai muscoli e cioè:
dall'efficienza dell'apparato respiratorio
dalla trasportabilità dell'O2 del sangue (concentrazione di globuli rossi e di emoglobina e quindi di ferro
nel sangue e nella dieta)
dall'efficienza del cuore (ampiezza delle sue cavità per pompare tanto sangue)
dalla capillarizzazione attorno alla fibra muscolare (sviluppo del sistema di irrigazione dei capillari)
dalla capacità di elaborazione chimica delle complesse reazioni (ciclo di Krebs) che dal glucosio/grassi
e O2 portano alla produzione di energia (ricarica dell’ATP). Ciò dipende dalla grandezza e
concentrazione di mitocondri che sono dei corpuscoli all'interno della cellula muscolare dove
avvengono queste reazioni - sono dei veri e propri laboratori chimici in cui si producono gli enzimi che
catalizzano il ciclo di Krebs per produrre energia per ricaricare ATP.
Quando compiamo uno sforzo elevato e breve (meno di un minuto) utilizziamo i meccanismi anaerobici
poiché quello aerobico non è in grado di fornire in tempo la sua energia proprio per la sua inerzia di
attivazione. Ma come mai subito dopo abbiamo il "fiatone"? Il meccanismo aerobico interviene infatti per
ripristinare le condizioni iniziali (ricarica dei pool dei fosfati, combustione dell'acido lattico) e questo
processo di recupero, detto pagamento del debito di ossigeno, richiede energia chimica che viene fornita,
successivamente allo sforzo, dal lento e quasi inesauribile meccanismo aerobico.
L'attesa necessaria per ripristinare le condizioni iniziale e quindi per pagare il debito di ossigeno, si dice
tempo di recupero o di ripristino.
Tempo minimo di lavoro per contrarre il
debito di O2
Meccanismo
Alattacido
Meccanismo
Anaerobico lattacido
8’’
40’’
Pagamento della metà del debito di O2
T/2 (semitempo)
22 secondi
15 minuti
(2 ore per completo smaltimento acido
lattico)