Apoptosi
Copyright (c) by W. H. Freeman and Company
La morte cellulare e la sua regolazione
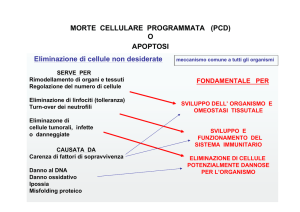
a La morte cellulare programmata è un processo fondamentale che controlla lo
sviluppo degli organismi pluricellulari;
a L’apoptosi porta all’eliminazione di intere strutture (es.: la coda durante lo sviluppo
embrionale dell’uomo), scolpisce tessuti specifici asportando campi di cellule (es.: il
tessuto tra le dita) e regola il numero di neuroni presenti nel sistema nervoso;
a Le interazioni cellulari regolano lo sviluppo in due modi sostanzialmente diversi:
La maggior parte delle cellule degli organismi pluricellulari hanno bisogno di
segnali (fattori trofici) per rimanere in vita e in assenza di tali segnali di
sopravvivenza la cellula attiva un programma di “suicidio”
In alcuni contesti di sviluppo (es.: sistema immunitario) specifici segnali
attivano un programma “assassino” che uccide le cellule
a In entrambi i casi la morte cellulare viene mediata dalla stessa via molecolare
Copyright (c) by W. H. Freeman and Company
La morte cellulare programmata avviene
tramite apoptosi
Cellula normale
La perdita di cellule mediante
morte cellulare programmata è
contraddistinta da una sequenza
ben definita di modificazioni
morfologiche, nel loro insieme
chiamate “apoptosi” (dal greco
“cadere da”).
Le cellule si condensano, e quindi
si suddividono in frammenti che
vengono fagocitati da altre cellule.
Le cellule che muoiono in risposta
ad un danno tissutale (necrosi)
aumentano di dimensioni e
scoppiano, rilasciando il loro
contenuto intracellulare.
Copyright
(c) by W. H. Freeman and Company
Cellula
apoptotica
Le neurotrofine promuovono la sopravvivenza dei
neuroni
Esperimenti di rimozione di tessuto o di
trapianto in embrioni di pollo hanno dimostrato
che il numero dei neuroni che innervano le
regioni periferiche dipende dalle dimensioni del
campo cellulare bersaglio.
I neuroni competono per i fattori di
sopravvivenza prodotti dal tessuto bersaglio.
I sarcomi producono grandi quantità di un
fattore trofico l’NGF.
I fattori trofici, detti neurotrofine sono: NGF,
BDNF (brain-derived growth factor) e la
neurotrofina-3 (NT-3).
Le neurotrofine si legano, attivandoli, ad una
famiglia di recettori tirosina chinasi chiamati Trk
Copyright (c) by W. H. Freeman and Company
(“track”)
Dimostrazione sperimentale che differenti neurotrofine
sono necessarie per la sopravvivenza di classi differenti
di neuroni sensoriali del ganglio della radice dorsale
Negli animali privi di NGF o dei suoi recettori TrkA, i neuroni sensibili al dolore
(nocicettivi) che innervano la pelle sono assenti.
Negli animali privi o della NT-3 o dei suoi recettori TrkC, sono assenti i neuroni
Copyright
(c) by W. H. Freeman and Company
propiocettivi che innervano i fusi
muscolari
Neurotrofine diverse sono necessarie per la
sopravvivenza di differenti classi di neuroni sensoriali
Il BDNF ed i recettori TrkB sono necessari per lo sviluppo dei neuroni
sensoriali dei gangli vestibolari, che innervano gli organi dell’orecchio
interno e che permettono la percezione del moto.
I mutanti per il BDNF presentano difetti dell’equilibrio associati a quelli
del sistema vestibolare.
La perdita dei propiocettori nei mutanti NT-3 si associa ad anomalie
posturali e motorie.
I mutanti per il l’NGF (e per i TrkA) in omozigosi sono letali, ed in
eterozigosi presentano una minore sensibilità al dolore.
Copyright (c) by W. H. Freeman and Company
Studi genetici in C. elegans hanno chiarito i
meccanismi molecolari che regolano l’apoptosi.
Delle 1090 cellule somatiche generate durante lo sviluppo di C. elegans,
131 vanno incontro ad apoptosi.
Mutazioni specifiche hanno portato all’identificazione dei geni coinvolti
nella regolazione dell’apoptosi
Copyright (c) by W. H. Freeman and Company
Studi genetici in C. elegans hanno chiarito i
meccanismi molecolari che regolano l’apoptosi.
a) Larva appena uscita dal
guscio con mutazione del
gene ced-1 che impedisce la
fagocitosi delle cellule morte.
b) Larva appena uscita dal
guscio con mutazione del
gene ced-1 e ced-3.
L’assenza di accumulo di
cellule morte dimostra che
l’apoptosi non avviene. La
proteina CED-3 è necessaria
per l’apoptosi.
Copyright (c) by W. H. Freeman and Company
Tre classi di proteine funzionano nella via apoptotica
Nei vermi che possiedono mutazioni dei geni ced-3 o ced-4, la morte
cellulare non si verifica e tutte le 1090 cellule sopravvivono
Nei vermi mutanti per il gene ced-9, tutte le 1090 cellule muoiono.
Le proteine CED-3 e CED-4 sono necessarie per la morte cellulare;
La proteina CED-9 inibisce l’apoptosi, e la via apoptotica può essere
attivata in tutte le cellule.
Nei doppi mutanti ced-9/ced-3 la morte cellulare non si verifica:
CED-9 agisce a monte di CED-3 inibendo la via apoptotica.
Copyright (c) by W. H. Freeman and Company
Il gene bcl-2 isolato in tumori umani
Il primo gene per l’apoptosi clonato nei mammiferi è stato il gene bcl-2.
Il gene bcl-2 è stato isolato nel linfoma follicolare umano come prodotto
di un riarrangiamento a livello di un punto di rottura.
Il gene bcl-2 funge da oncogene, promuovendo la sopravvivenza delle
cellule piuttosto che la loro moltiplicazione
Le proteine Bcl-2 e CED-9 sono omologhe e fungono da proteine
regolatrici che inibiscono la via apoptotica.
Copyright (c) by W. H. Freeman and Company
L’apoptosi è una via conservata durante l’evoluzione
L’effettore più importante in C. elegans è la proteasi CED-3.
CED-3 è una caspasi (cisteine proteasi che scindono le proteine in
corrispondenza di siti in posizione C-terminale rispetto a residui di aspartato).
Le cellule dei mammiferi contengono molteplici caspasi.
CED-9 e BCL-2 sono proteine di membrana e la CED-4 può legarsi sia alla CED-9
che alla CED-3.
Copyright (c) by W. H. Freeman and Company
Proteine regolatrici pro-apoptotiche promuovono
l’attivazione delle caspasi
La famiglia Bcl-2 comprende sia membri anti-apoptotici (la CED-9 in C.
elegans e Bcl-2 e Bcl-xl nei mammiferi) che membri pro-apoptotici (Bax).
Sono tutte proteine transmembrana, con un unico dominio che attraversa
la membrana. Possono partecipare ad interazioni oligomeriche e sono
localizzate sulle membrane mitocondriale esterna, nucleare e del RE.
Il destino di una data cellula dipende dai membri della famiglia Bcl-2
presenti
I membri della famigli Bcl-2 possono influenzare la distribuzione
subcellulare del citocromo c, che contribuisce all’attivazione delle
caspasi.
Copyright (c) by W. H. Freeman and Company
Proteine regolatrici pro-apoptotiche promuovono
l’attivazione delle caspasi (2)
Nelle cellule normali e sane, il citocromo C è localizzato tra la membrana
mitocondriale esterna e quella interna, mentre nelle cellule che vanno
incontro ad apoptosi viene rilasciato nel citosol.
Nel citosol il legame del citocromo c alla proteina di raccordo Apaf-1 (la
CED-4 dei mammiferi) induce l’attivazione della cascata delle caspasi.
Il rilascio del citocromo c nel citosol è inibito da una espressione elevata
di Bcl-2, e stimolato da una espressione elevata di Bax.
Non è ancora chiaro come, ma gli omodimeri Bax consentono un influsso
di ioni attraverso la membrana mitocondriale esterna che favorisce il
rilascio del citocromo c.
Copyright (c) by W. H. Freeman and Company
Alcuni fattori trofici impediscono l’apoptosi inducendo
l’inattivazione di una proteina regolatrice pro-apoptotica
Copyright (c) by W. H. Freeman and Company
Alcuni fattori trofici impediscono l’apoptosi inducendo
l’inattivazione di una proteina regolatrice pro-apoptotica
Le neurotrofine, come l’NGF, proteggono i neuroni dalla morte cellulare.
In assenza di fattori trofici, la forma non fosforilata della proteina Bad è
associata all’eterodimero Bcl2/Bcl-xl, localizzato a livello della membrana
mitocondriale esterna, e ne inibisce la funzione anti-apoptotica.
La Bad fosforilata non può legarsi all’eterodimero Bcl2/Bcl-xl ed è
localizzata nel citosol, legata alla proteina 14-3-3 con residui di fosfoserina.
Le vie di trasmissione del segnale che portano alla fosforilazione della Bad
trasmettono segnali di sopravvivenza.
Numerosi fattori trofici attivano la PI-3 chinasi, che attiva la chinasi Akt
(protein chinasi B). La Akt fosforila Bad e ne inibisce l’attività proapoptotica.
Copyright (c) by W. H. Freeman and Company
Via di trasduzione del segnale dell’insulina
Ras-indipendente
a L’IRS1 fosforilato si lega anche alla PI-3 chinasi stimolandone l’attività chinasica;
a La proteina chinasi B (protein kinase B, PkB) viene richiamata verso la membrana
grazie alla regione N-terminale che contiene un dominio PH (di omologia alla
Pleckstrina) che si lega ai fosfoinositidi della membrana plasmatica;
a La PKB, posizionata a livello della membrana, viene fosforilata e quindi attivata, da 2
chinasi associate alla membrana;
a La PKB attivata viene rilasciata nel citosol e media molti degli effetti dell’insulina,
compresa la stimolazione dell’assunzione del glucosio e della sintesi del glicogeno.
a La PKB, detta anche Akt, è una componente della via per la trasmissione del segnale
che previene la morte cellulare.
Copyright (c) by W. H. Freeman and Company
La via Ras indipendente attiva la Proteina chinasi B
GSK3 = glicogeno sintetasi chinasi 3
Copyright (c) by W. H. Freeman and Company
Alcuni fattori trofici impediscono l’apoptosi inducendo
l’inattivazione di una proteina regolatrice pro-apoptotica
Copyright (c) by W. H. Freeman and Company
Per vedere questa immagine
occorre QuickTime™ e un
decompressore Animation.
Copyright (c) by W. H. Freeman and Company