Regolazione dell’espressione genica
nei batteriofagi:
il ciclo litico ed il ciclo lisogeno del
fago lambda
Argomento di vaste proporzioni che non si può esaurire in una singola lezione. L’argomento di questa
lezione
sono i fagi temperati ed in particolare il fago lambda ed il meccanismo con cui essi sono capaci di
scegliere tra due possibili
strategie di moltiplicazione.
Una lezione di questo tipo prevede perciò una premessa: la preventiva conoscenza da parte degli
studenti della struttura di un batteriofago
dei fagi virulenti che dovrebbe, tra l’altro, aver portato alla consapevolezza che un virus è molto di più che
una semplice associazione
tra un acido nucleico e proteine e che pressioni selettive molto forti devono essere state responsabili
della compattazione
del genoma e della sua organizazione spaziale e temporale del genoma virale.
E ciò non può essere visto come una cosa a sé stante, svincolato dall’ospite. Anzi, in realtà la capacità
dei virus di utilizzare il macchinario cellulare
per far avvenire le complesse reazioni necessarie per la propria moltiplicazione implica un alto grado di
specializzazione,
il raggiungimento drel quale deve aver richiesto una lunga evoluzione nel corso della quale forti pressioni
selettive hanno agito
sia sui virus che sulle cellule da essi infettate.
Questop, se possibile, è ancor più vero nel caso dei fagi simbionti o temperati (fagi non virulenti).
Che cosa succede allora quando un fago temperato, come il fago lambda, infetta una cellula ospite?
Esso può escegliere tra due
strategie si “sopravvivenza” alternative.
Due domande:
1) quali siti sono implicati nella specifictà della
terminazione
2) come funzionano gli antiterminatori?
La stretta relazione esistente tra un batterio e un fago
temperato è
evidente nel caso del fago lambda
⇓
nello stato di profago non si ritrova in siti casuali del
cromosoma batterico, ma in un sito preciso e specifico,
al contrario, ad esempio del fattore F, che può integrarsi
in punti diversi del cromosoma
⇓
attB e attP
⇓
devono avere subito una coevoluzione
Voi sapete che il virus lisa la cellula solamente
alla fine
⇓
sarebbe folle che esprimere i geni della lisi
prima di essersi e prima di aver
sintetizzato testa, coda etc….
⇓
vi deve essere una perfetta sintonia
nella espressione temporale di questi geni
⇓
una sintonia temporale che è associata
ad una sintonia spaziale
Come si può realizzare
la sintonia spazio-temporale?
Mutazioni nel gene N
⇓
ciò significa che in qualche modo N
permette la trascrizione dei geni precoci ritardati
⇓
La trascrizione dei geni N e cro si ferma
ai terminaroti Rho-dipendenti
Q è un antiterminatore diverso da N che
permette la trascrizione dei geni tardivi!
Ciò significa che N e Q hanno una diversa
specificità
Principio generale
L’identificazione del siyto sul DNA in cui una
proteina esercita il suo effetto non
necessariamente corrisponde con la sequenza
che viene riconosciuta
⇓
i due siti possono essere separati
È plausibile supporre che N si leghi da
qualche parte sul DNA del fago labmda. Dove?
Mediante mappatura è stato possibile identificare
i siti di riconoscimento di N
⇓
dove sono localizzati?
Per capire come funziona questo interruttore
genetico dobbiamo prendere in
considerazione la mappa genetica del fago lambda
che può
essere rappresentata in forma circolare anche
se in realtà nella forma vegetativa è lineare
Vi è una espressione genica a cascata
⇓
ha una sua corrispondenza nella organizzazione
di questi geni
⇓
dove è che si ha necessità di un
controllo spazio-temporale dell’espressione
dei geni?
⇓
nei virus!!
CICLO DEL FAGO λ
VIA LISOGENA
VIA LITICA
VIA LISOGENA
VIA LITICA
PROFAGO
Immunità
Induzione
U.V.
Perché la
LISOGENIA?
Vantaggi ?
La lisogenia favorisce la persistenza e la
diffusione di un virus più efficacemente
della virulenza
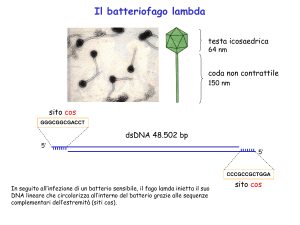
FAGO λ
DNA a doppia elica
lineare
48514 bp
Escherichia coli
12 basi
VIRUS
3’
5’
5’
3’
12 basi
Infezione
Ligasi
12 basi
Ligasi
BATTERIO
REGOLAZIONE
Geni PRECOCI
IMMEDIATI
cro
N
inibisce
antiterminatore
cII e cIII
Geni PRECOCI
RITARDATI
2 geni REPLICAZIONE
10 geni TESTA
11 geni CODA
2 geni LISI
attivano
7 geni RICOMBINAZIONE
Q antiterminatore
Geni TARDIVI
cI
repressore
Mutanti cI
Placche chiare
via litica
Mutanti cro
Placche torbide
via lisogena
cI
necessario per l’instaurarsi della
via lisogena
cro
necessario per l’instaurarsi della
via litica
crescita del batterio
Terreno povero di nutrienti
cI
Terreno ricco di nutrienti
cro
Via LISOGENA
cro
cI
Via LITICA
REPRESSORE E CRO
COOH
COOH
COOH
COOH
COOH
NH
2
NH
2
NH
2
NH 2
DNA
REPRESSORE
CRO
DNA
NH 2
Repressore
TL1
cIII
TR1
N
cI
cro
PLOL
cII
PROR
cro
REGIONE REGOLATIVA
N
TL1
cIII
TR1
N
cI
cro
PLOL
cII
PROR
cro
INFEZIONE
Mutanti del fago λ nel gene N
Trasfezione dei soli
geni precoci
immediati
non lisano le cellule di
E. coli
Meccanismo
pN riconosce nut
Azione sulla RNA-polimerasi
NON risponde al terminatore
Poiché la localizzazione di nut è VARIABILE
né inizio né terminazione della
TRASCRIZIONE
pN agisce sulla RNA-pol durante il suo
movimento
Probabilmente pN riconosce nut
dell’RNA, ma NON si LEGA né al
DNA né all’RNA
pN si lega alla
RNA-POLIMERASI
cIII
N
TL1
cIII
TR1
PM
N
cI
cro
PLOL
PROR
PRE
cro
?
cII
cII
Repressore
RNA-pol
cII
TL1
cIII
N
cI
cro
PLOL
PROR
cII
PRE
TL1
cIII
N
cI
cro
PLOL
PROR
cII
PRE
FUNZIONE
cII
Prende contatto con la
regione -35 del promotore
pRE
RNA-pol può iniziare la trascrizione a
partire da pRE
INTEGRASI
RNA-pol
cII
TL1
cIII
int
PI
N
cI
PLOL
cro
PROR
cII
PRE
Integrazione del DNA virale nel genoma
batterico
attP
bio
gal
attB
Integrazione
(Int)
gal
Excisione
(Int e Xis)
bio
TL1
cIII
N
cI
PLOL
cro
PROR
cII
PRE
U.V.
TL1
cIII
N
cI
PLOL
cro
PROR
cII
PRE
TL1
cIII
N
cI
PLOL
cro
PROR
cII
PRE
cro
ESPRESSIONE GENICA nella VIA LITICA
MUTAZIONI CHE AUMENTANO LA STABILITÀ DI CII
AUMENTANO LA FREQUENZA DELLA LISOGENIA
IL LIVELLO DI CII CHE DETERMINA
L’ESITO DELL’INFEZIONE.
INSTABILITA’ DI CII
λ
CIII
hflA hflB
E. coli
Mutazioni in:
cIII
Inattivazione di cII
cIII protegge cII dalla degradazione ad opera di una
proteasi codificata dai geni
hflA
Stabilizzazione di cII
hflB
cAMP
(+)
(-)
Proteasi
Hfl
(-)
cII
(+)
PI
cIII
(-)
PRE
AZIONE DEL REPRESSORE
PM
O R3
O R2
O R1
cI
cro
Promotore
per cI
Promotore
per cro
cI
cro
RNA-pol
cI
cro
cI
cro
RNA-pol
mRNA
INDUZIONE
RNA-pol
cI
cro
mRNA
U.V.
cI
RecA
cro
O R3
O R2
O R1
cI
cro
RNA-pol
cI
cro
RNA-pol
cI
cro
cro
Repressore LexA
Ad altri geni controllati da LexA
proteina
RNA messaggero
lexA
recA
uvrA
uvrB
dimero
Proteasi RecA
Repressore LexA scisso
Repressore LexA
proteina
lexA
RNA messaggero
recA
uvrA
uvrB
TL1
cIII
N
cI
PLOL
cro
PROR
cII
PRE
U.V.
TL1
cIII
N
cI
PLOL
cro
PROR
cII
PRE
TL1
cIII
N
cI
PLOL
cro
PROR
cII
PRE
cro