1
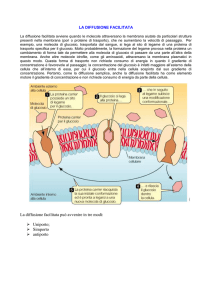
Glicazione non enzimatica dell’emoglobina
La glicazione è un processo post-traduzionale per mezzo del quale uno zucchero interagisce con una
proteina con il risultato di ottenere una proteina modificata con uno zucchero legato covalentemente a
uno o più amminoacidi della proteina stessa . La struttura secondaria della proteina è perciò alterata
(rispetto alla proteina non glicata ) spesso dando origine ad un cambiamento della funzione della proteina
glicata . Sebbene avvengano modeste glicazioni in tutte le persone in salute , ad alti livelli di glicazione
possono verificarsi cambiamenti fisiologici e patologici . Come esempio un individuo iperglicemico ha un
alto grado di glicazione e spesso manifesta complicazioni nel diabete mellito tipo 2 . In aggiunta , il legame
covalente del glucosio all’estremità N-terminale della valina della emoglobina è servito come marker
della concentrazione di glucosio nel sangue lungo un periodo esteso (2-3 mesi ). Poiché la glicazione delle
proteine ha implicazioni sullo stato di salute , la comprensione del processo di glicazione ha grande valore.
L’obbiettivo di questa lezione è quello di comprendere meglio il primo stadio della glicazione proteica , la
quale consiste nell’iniziale interazione tra lo zucchero e la proteina . In particolare il glucosio sarà lo
zucchero polare che funge da substrato e la forma adulta dell’emoglobina umana (HbA) sarà la proteina
modello in questa lezione .
1. Le proteine possono interagire con i substrati in modo tale che l’interazione proteina/substrato può
essere di natura covalente o non covalente .
Il Riquadro A rappresenta una interazione proteina / substrato covalente o non covalente ?
Non covalente
Il Riquadro B rappresenta una interazione proteina/ substrato covalente o non covalente ?
Covalente
2. Quale è l’interazione non covalente prevalente tra un substrato e una proteina ?
Un suggerimento: Nel Riquadro A l’interazione predominante non covalente è raffigurata con una
linea tratteggiata ( tra il paio di elettroni solitario sull’azoto del residuo amminoacidico e il protone
sul gruppo – OH del substrato , in questo caso , uno zucchero )
2
Legame ad Idrogeno
3. Nel legame ad idrogeno ci sono donatori di legami-H e accettori di legami -H .
a. Nel Riquadro A , il residuo amminoacidico è accettore del legame a idrogeno o donatore del legame
a idrogeno ?
Accettore di legame-H
b. Nel Riquadro B , il residuo amminoacidico è accettore del legame a idrogeno o donatore del legame a
idrogeno ?
Donatore di legame -H
4. Nel nostro studio sul processo di glicazione , il substrato in questione è il glucosio . Il glucosio non
è una singola specie , ma sono invece cinque strutture che si interconvertono ( il processo di
interconversione di queste specie è chiamato mutarotazione ) in soluzione acquosa .
3
Basandoci sulla abbondanza percentuale della forma ad Anello – Aperto (AA) di glucosio all’equilibrio , la
forma AA è più o meno disponibile ad interagire con una proteina rispetto alle quattro forme ad Anello Chiuso (AC) del glucosio ?
Meno disponibile
5. I ricercatori concordano nel ritenere che la forma AA del glucosio è l‘unica forma che interagisce
con le proteine nel processo di glicazione . Questo assunto è illustrato nello schema seguente :
Basandoci sulla Fig. 1 , la struttura del glucosio , che è indicata come necessaria per la prosecuzione del
processo , è evidenziata dalla intestazione “Non covalent interactions : Stage 1 “. E’ la forma AA o AC del
glucosio?
La forma ad Anello aperto
6. Poiché il processo è chiamato “non covalent “ , quale è la più probabile interazione tra la forma AA
del glucosio e l’amminoacido terminale di HbA (vedi domanda 2 e 3 ) ?
Legame ad Idrogeno
4
7. Il complessivo processo di glicazione è diviso in quattro stadi ( vedi Fig.1) . Da notare che lo stadio 1
è il processo non covalente e gli stadi 2-4 coinvolgono interazioni covalenti . Si conosce molto degli
stadi 2-4 della glicazione e perciò non saranno al centro della nostra attenzione . Invece , il nostro
interesse è nel processo non covalente , l’iniziale legame tra HbA e il glucosio.
Sapendo che il 99.998% del glucosio disponibile in soluzione acquosa non è nella forma AA (vedi domanda
4) , e che la forma di glucosio che si suppone continui nel processo covalente è la forma AA , dobbiamo
allora porci una domanda . Come possiamo avere una significativa glicazione nonostante sia così piccola la
quantità disponibile , per reagire , di glucosio nella forma AA ?
Per aiutare a dare una risposta a questa domanda considereremo un meccanismo semplificato “ chiave e
serratura “ per un substrato che si lega ad una proteina .
a. E’ possibile che la forma AA si adatti al sito di legame meglio delle altre quattro forme AC di
glucosio?
Si
b. Se la forma AA si adatta al sito di legame meglio delle altre forme , è concepibile che AA sia il solo
che si leghi ?
Si
8. Non esiste un metodo sperimentale diretto , accessibile per determinare se la forma AA lega meglio
delle altre quattro forme ad anello chiuso (AC) .Tuttavia è possibile risolvere questa questione con
l’ausilio del computer. Si può costruire un modello di substrato e legare questo substrato alla
proteina , sotto definite condizioni , attraverso un modello compiutazionale . Attraverso questo
metodo è possibile misurare le caratteristiche esoergoniche del legame di qualsiasi substrato a una
proteina ( si può calcolare un teorico ΔG per ciascun substrato che venga legato )
L’energia libera di Gibbs per una reazione (ΔG = ΔH – TΔS ) prende in considerazione sia l’entalpia ΔH che
l’entropia ΔS . Se ΔG è negativo , la reazione (o processo ) sarà spontaneo e sarà chiamato esoergonico .
Data il problema che abbiamo se la forma AA del glucosio si lega meglio ad HbA rispetto alle quattro
forme AC del glucosio , viene progettato un approccio computazionale per rispondere alla domanda 7 . In
modo specifico quale substrato potrebbe essere costruito ? Quale proteina potrebbe essere costruita ? .
Che cosa si fa con la struttura una volta costruita e che cosa si misura ?
5
Si potrebbe costruire la proteina HbA , nonché i 5 isomeri di glucosio (anello aperto, forme piranosiche e
forme furanosiche ). Quindi si potrebbe far ineragire ciascuno degli isomeri ad HbA per determinare la
esorgonicità dell'associazione per ogni isomero , dandoci un modo per confrontare la capacità di ogni
isomero di legarsi ad HbA.
9.
Se si fosse trovato che tra i cinque isomeri del glucosio la forma AA è la sola che si lega in modo
esoergonico ( mentre le altre si legano in modo endoergoniche ) , o la forma AA si lega in modo più
esoergonico rispetto alle altre forme AC , come possiamo rispondere alla seguente questione …..
La forma AA si lega selettivamente ad HbA in presenza delle quattro forme di AC ?
Si
10. Se la esoergonicità del legame dei cinque isomeri del glucosio è circa la stessa , o se la forma AA in
realtà si lega meno bene di quanto facciano le forme AC ( o al meno una delle forme AC ) , come
risponderesti alla seguente domanda …
La forma AA si lega in modo selettivo ad HbA in presenza delle quattro forme AC ?
No
Esperimento 1 : Furono eseguiti una serie di calcoli dopo che erano stati costruiti ciascuno dei cinque
isomeri e ciascuno indipendentemente legato alla catena β di HbA . La esorgonicità del legame in ciascun
esperimento è riportata in Tabella 1 .
11. Basandoci sui dati dell’esperimento 1 (Table 1 ) la forma AA ( Ring – opened glucose ) si lega con
una esorgonicità più grande rispetto a tutte le altre forme di AC ?
No
12. E’ più facile che la forma AA ( Ring – opened glucose ) del glucosio lega selettivamente HbA rispetto
alle quattro forme di AC?
No
13. Perciò , dato che la forma AA non è quella che si lega esclusivamente ( in presenza delle quattro
forme di AC ) , il meccanismo della chiave e serratura descritto nella domanda 7 si può applicare
qui ?
No
6
14. Se la forma AA non si lega selettivamente , allora sorge una domanda . Come finisce per legarsi la
forma AA ad HbA ? La domanda è importante perché la forma AA del glucosio è la sola forma che
precede la glicazione (Figura 1) , però è soltanto lo 0.002% rispetto alle forme AC (vedi domanda
4) e inoltre la forma AA non si lega selettivamente . Così di nuovo …. Come la forma AA del glucosio
finisce per legarsi ad HbA ? Per rispondere a questo quesito , consideriamo e rispondiamo a una
serie di relative domande di fondo .
a. Basandoci sui dati della Tabella 1 quale forma di glucosio si lega meglio ?
β -piranosio
b. 1.0 – 1.2 kcal/mole di differenza nella esorganicità del legame non è sufficiente per
aspettarci che una specie si legherà, in presenza di un’altra , in modo esclusivo . Perciò
quale isomero potrebbe legarsi insieme al β – piranosio ?
α-piranosio
c. Se α- e β- piranosio si legano come l’isomero AA ( anzi meglio ) e la concentrazione di αe β- piranosio è circa 50.000 volte più alta rispetto all’isomero AA , quale isomero del
glucosio quasi esclusivamente si lega inizialmente a HbA ?
α-piranosio e
β -piranosio
15. Poiché ci sono 50.000 α- e β- piranosio molecole di glucosio AC per 1 AA glucosio disponibile a
legarsi e poiché le strutture di glucosio α- e β- piranosio si legano meglio rispetto all’isomero AA , è
più facile che il legame non covalente del glucosio AA alla proteina (come mostrato in Figura 1 )
nasca dal legame diretto del glucosio AA ?
No
Perciò , poiché la formazione di un legame di AA è necessario perché si vada verso il processo di
glicazione , e esso non nasce da un legame diretto , noi ritorniamo alla nostra precedente
domanda ancora senza risposta ….
Come la forma AA del glucosio finisce per legarsi ad HbA ? Ma ….. ancora non possiamo
rispondere a questa domanda senza rivedere un altro concetto specifico , la mutarotazione del
glucosio .
16. Nella figura sotto , nota le frecce degli equilibri . Se si parte con α- piranosio , quale è il solo
isomero al quale possiamo direttamente arrivare ?
Il glucosio ad anello
aperto
7
17. Considerate le frecce dell’equilibrio , può l’isomero AA tornare indietro ad α- piranosio ?
Si
18. Una volta che siamo all’isomero AA , l’isomero AA è limitato a ritornare solo all’ α- piranosio ?
No
19. Quanti composti si ottengono partendo dall’isomero AA ( incluso il ritorno all’ α- piranosio ) ?
Quattro
20. Ancora , osservando le frecce degli equilibri , in particolare in termini delle loro relative dimensioni
e verso, che partono dall’isomero AA , sarà l’isomero AA ad avere una lunga vita o il suo anello si
chiuderà rapidamente in una delle quattro forme AC
L’anello si chiuderà rapidamente
21. Dopo aver riesaminato il processo completo di mutarotazione , possiamo ritornare al legame del
glucosio e al processo di glicazione . Se l’isomero AA è la sola specie reattiva che conduce in
direzione della glicazione e, come abbiamo già dimostrato , la forma AA non esiste in grande
concentrazione e non si lega selettivamente ---- Come possiamo arrivare alla specie AA legata ?
Prima di rispondere , ricorda che AC α- e β-piranosio inizialmente si legano ad HbA .
Consideriamo lo schema dell’equilibrio della mutarotazione del glucosio , domanda 16. In questo
schema , la interconversione tra i quattro isomeri AC e l’isomero AA a vita breve si svolge in
soluzione acquosa . Per il nostro scopo , il mezzo acquoso potrebbe essere il citosol dell’eritrocita
(che contiene la proteina ) . Tuttavia , perché l’isomero del glucosio si interconverta in acqua
( processo di mutarotazione ) è ragionevole supporre che un simile processo di mutarotazione può
avvenire una volta che un isomero AC è legato in modo non covalente ad una proteina ? Se è così ,
è ragionevole asserire che il glucosio AC si lega e poi , mentre è legato , mutoruota e genera un
isomero intermedio AA ?
Si
Se è così , un isomero AA può diventare non legato covalentememte attraverso la mutarotazione
di un isomero AC che si era legato precedentemente alla proteina . Questo forma AA può poi
8
muoversi attraverso il resto del processo di glicazione . Per illustrare questo , lo schema seguente
rappresenterebbe il legame non covalente di una molecola di AC β- piranosio a una proteina nella
fase della muratotazione per diventare l’isomero AA ( senza mostrare nessun stadio del
meccanismo )
Il riquadro 1 rappresenta la condizione di pre-legame e il riquadro 2 l’iniziale legame non
covalente del β- piranosio alla proteina . Per aiutare la visualizzazione del processo , completa lo
schema disegnando il glucosio AA legato alla proteina nel terzo riquadro …..
Con lo schema precedente in vista possiamo riassumere dove siamo …
Riassunto delle ipotesi per la formazione del legame del Glucosio AA :
a. Le strutture di glucosio AC α- e β- piranosio sono gli unici isomeri che inizialmente si legano ad
HbA non covalentemente .
E’ questo elemento della nostra ipotesi convalidato dai dati ?
Si
Se così , da dove traiamo i dati a sostegno ?
Tabella 1
b. Una volta legatosi , le strutture del glucosio α- and β-piranosio possono aprire l’anello per dare
l’isomero AA reattivo così che questi prosegue verso il processo di glicazione ( questo è descritto
sopra nel disegno che hai completato )
Le nostre ipotesi complessive per la formazione del glucosio AA reattivo legato non fanno
riferimento alla natura della reattività del legame del glucosio AA , perciò dobbiamo domandarci
come il glucosio AA legato reagisce per procedere nel processo di glicazione .
22. Per gli artefatti , andiamo a rivedere la Figura 1.
9
La transizione dallo Stadio 1 allo Stadio 2 richiede che si formi un legame covalente . Il legame covalente
formatosi deve essere il risultato dell’attacco di un nucleofilo verso un elettrofilo .
Nelle due illustrazioni seguenti , lo zucchero AA agisce come nucleofilo o elettrofilo ?
Elettrofilo
Nelle due illustrazioni seguenti , il doppietto solitario di NH2 nel residuo amminoacidico agisce come
nucleofilo o elettrofilo ?
Nucleofilo
Se il residuo amminoacidico nella seguente illustrazione è protonato ( così che è NH3+ ) la reazione non
procederebbe . Perché è necessario che il residuo amminoacidico sia deprotonato per agire da nucleofilo ?
Se il residuo amminoacidico è protonato , non è più disponibile la coppia di elettroni solitaria sull’azoto
perché questi possa agire come nucleofilo .
10
23. Lo scopo dei due precedenti schemi ( domanda 22 ) è di stabilire che un residuo amminoacidico (Nterminale o gruppo R- ) deve agire come nucleofilo per formare un legame covalente guidando verso la
glicazione . Così … quale amminoacido interno , in generale , può agire come un nucleofilo ?
Suggerimento: solo quelli il cui gruppo R ha un atomo che possiede una coppia di elettroni ….
Nel segmento di proteina mostrato sotto , quale residuo amminoacidico ( rispondi a fianco all’elenco degli
amminoacidi si o no ) può servire come nucleofilo ?
si o no
No
No
No
Si
Tra gli amminoacidi precedenti , quale residuo amminoacidico in una proteina come HbA potrebbe essere
glicosilato ? Spiega .
Lisina . Tra i residui amminoacidici precedenti , la lisina è il solo residuo amminoacidico che ha un
gruppo funzionale che porta una coppia solitaria di elettroni , capace di agire come nucleofilo e
formare un legame covalente che porti alla reazione di glicazione
Nota : I soli residui amminoacidici interni glicati in HbA noti sono residui di lisina , che è coerente con la
nostra ipotesi .
Data la conoscenza acquisita con le ultime due domande , quale è poi il ruolo del glucosio AA legato nel
processo di glicazione ----- è nucleofilo o elettrofilo ?
Elettrofilo
24. Il disegno seguente differisce dal segmento di proteina precedente in quanto tale segmento non è
un segmento interno ma è piuttosto l’estremità N-terminale di una proteina .
Basandoci su questo disegno e su cosa conosciamo del meccanismo di glicazione , può la valina terminale
agire come nucleofilo ?
Si
Che cosa accadrebbe alla reattività della valina terminale se l’ NH2 terminale fosse protonato a NH3+ ?
( aumenterebbe o diminuirebbe )
Diminuirebbe
11
25. I ricercatori hanno sostenuto storicamente che l’isomero AA del glucosio è la sola forma del glucosio
reattivo che procede verso il processo di glicazione per produrre la base aldimmina /Schiff ( transizione
Stadio 1 – Stadio 2 ) . Basandoci sulle nostre nuove dimostrazioni che il glucosio legato è l’elettrofilo , quale
è il miglior elettrofilo , l’iniziale anello chiuso del glucosio o il glucosio come AA che si genera dalla
mutarotazione dell’iniziale glucosio ad anello chiuso ? Perché ? ( vedi la figura sotto )
Il glucosio ad anello aperto che si genera per mutarotazione potrebbe essere l’elettrofilo migliore perché
il suo gruppo carbonio anomerico contiene un gruppo carbonile
26. Fino ad ora noi sappiamo che :
a. La specie legata inizialmente è un anello chiuso α- o β- piranosio
b. La struttura legata inizialmente del glucosio α- o β-pyranosio, mentre si lega ,
genera l’isomero del glucosio AA .
c. La sola forma reattiva del glucosio legato che procede verso la glicazione è
l’isomero AA del glucosio .
d. L’isomero AA del glucosio agisce come elettrofilo ed è attaccato da un residuo
amminoacidico nucleofilo di lisina ( o il terminale N dell’amminoacido ) della
proteina per formare la base aldimmina /Schiff ( transizione Stadio 1 – Stadio ),
così …
L’iniziale isomero AC del glucosio apre l’anello prima o dopo l’attacco nucleofilo ?
Prima
27. Come si apre l’anello del glucosio legato α- o β-pyranosio mentre si sta legando?
Per rispondere a questo è necessario rivedere alcuni principi .
Basandoci sulla mutarotazione in soluzione , il meccanismo dell’apertura dell’anello del glucosio è
teoricamente acido o base catalizzato .
a. Nel meccanismo seguente , rappresentato sotto , chi agisce come un acido ( l’α-piranosio o il
fosfato inorganico ) ?
α-piranosio
b. Chi agisce come una base ?
Il fosfato inorganico
12
c. Il meccanismo porta alla apertura dell’anello ?
Si
d. Il meccanismo di apertura dell’anello è acido catalizzata o base catalizzata (riferito al reagente non
zucchero ) ?
Base – catalizzata
28. Una seconda alternativa è un meccanismo acido catalizzato .
a. In questo caso ( mostrato sotto ) quale è l’acido ?
Lo ione idrogeno
b. Quale è la base ?
α - piranosio
c. Il risultato è l’apertura dell’anello?
Si
13
29. Abbiamo bisogno di progettare un esperimento ( o esperimenti ) per stabilire se l’anello che si sta
aprendo del glucosio legato alla proteina è catalizzato da un acido o una base . Questo è molto difficile da
realizzare con una proteina realmente coinvolta . Così , una serie semplificata di esperimenti può essere
compiuta per aiutarci a rispondere alla domanda . Consideriamo la serie di sperimenti seguenti ….
Esperimento 2 : Verifica per la Catalisi Acida .
Se si dovesse condurre la reazione dell’apertura dell’anello dell’α-piranosio in funzione del tempo in D2O
con l’aggiunta di acido acetico glaciale ( vedi struttura seguente ) fino a un pH di circa 4.5 , sarà disponibile
più acido in questa reazione che non era disponibile in una reazione a pH = 7 ( la reazione standard in
condizioni fisiologiche ) ?
( Si o no )
Si
Acido acetico
Se si eseguivano altri due esperimenti ---- uno dove l’acido acetico è addizionato fino a pH circa 3 e l’altro
dove l’acido acetico è addizionato fino a pH circa 1 , avremmo tre serie di dati con una concentrazione
sempre maggiore di acido disponibile . Perciò , potresti confrontare la velocità di apertura dell’anello in
funzione del’ aumento della concentrazione di acido ?
Se l’apertura dell’anello dell’ α- piranosio può essere catalizzata da acidi , la relativa velocità dovrebbe
aumentare o diminuire se l’acidità aumenta ? Spiega .
Aumenta . La velocità relativa dovrebbe aumentare come l’acidità aumenta . Se l’apertura dell’anello
dell’α-piranosio è catalizzata dagli acidi allora la cinetica di reazione dipenderà dalla concentrazione
dell’acido .
Se l’apertura dell’anello dell’ α- piranosio non può essere catalizzata da acidi , che cosa potrebbe accadere
alla velocità relativa se l’acidità aumenta ? Spiega .
La velocità relativa non dovrebbe variare se aumenta l’acidità . Se l’apertura dell’anello non è
catalizzata dagli acidi , allora la cinetica della reazione non dipenderà dalla concentrazione dell’acido .
14
Basandoci sui dati della Tabella 2 , può l’apertura dell ‘ anello di α-piranosio essere catalizzata da acidi ?
( Si o no )
Si
Esperimento 3 : Verifica per la Catalisi Basica
Se noi misuriamo la velocità dell’ apertura dell’anello di α- piranosio in funzione della forza della base
aggiunta , possiamo valutare se l’apertura dell’anello può essere catalizzata dalla base . Specificamente ,
tre basi fisiologiche sono esaminate : fosfato , bicarbonato , e lattato ( i pKa per queste basi sono circa 12.1,
10.3 , e 3.9 rispettivamente ) .
Se l’apertura dell’anello dell’ α-piranosio può essere catalizzata da una base , la relativa velocità dovrebbe
aumentare o diminuire se cresce la forza della base ?
Aumentare
Se l’apertura dell’anello dell’ α-piranosio non può essere catalizzata da una base , che cosa dovrebbe
accadere alla relativa velocità se la forza della base cresce ?
Non variare
Basandoci sui dati della Tabella 3 , può l’apertura dell’anello dell’ α-piranosio essere catalizzata da una
base? ( Si o no )
Si
30. Basandoci sui dati della Tabella 2 e 3 , l’apertura dell’anello del glucosio può essere catalizzata sia da un
acido che una base . Così cosa è meglio , la catalisi acida o basica ? Per aiutare in questa risposta ,
consideriamo che la reazione standard in entrambe le Tabelle 2 e 3 sono le stesse , cioè , la prima
15
annotazione in ciascuna tabella riflette la stessa velocità . Perciò , i numeri delle Tabelle 2 e 3 possono
essere direttamente confrontati .
a.
Perciò , addizionando un acido , la velocità di apertura dell’anello del glucosio è accelerata più o
meno rispetto all’aggiunta di una base ?
Meno rispetto alla aggiunta di una base . La relativa velocità di apertura dell’anello aumentando
l’acidità accellera di un fattore di 3 , mentra la velocità può essere accellerata dal bosfato di un
fattore di 25 .
b. Perciò , cosa catalizza meglio un acido o una base ?
Base
c. Quali condizioni sono responsabili per l’apertura più veloce dell’anello ? ( Acidità o Basicità )
Basicità
31. Sebbene la β- catena di HbA ha molti residui di lisina , i residui glicati sono Lis 17 , 59 , 61, 66, e 82 .
Perché solo questi ? Perché non tutti i residui di Lisina contenuti nell’intera proteina ?
Precedentemente abbiamo stabilito che l’apertura dell’anello è facilitata dalla base ( vedi domande 27 ,
30). Se in presenza di una base aumenta la velocità di apertura dell’anello di uno zucchero legato , la
presenza di un amminoacido basico ( in aggiunta alla presenza di un nucleofilo ) dovrebbe giocare un ruolo
nella velocità .
a. Potrebbe la presenza di un amminoacido basico far aumentare o diminuire la concentrazione di
uno zucchero AA legato ?
Aumentare
b. Potrebbe la presenza di un amminoacido basico far aumentare o diminuire la concentrazione di un
elettrofilo legato nella reazione ?
Aumentare
c. Se la concentrazione dell’elettrofilo legato è aumentata , che cosa potrebbe accadere alla velocità
della glicazione ?
Aumentare
Nel segmento di proteina mostrato sotto , quale residuo amminoacidico ( catene laterali nelle caselle ) può
comportarsi da base ?
Si / No
No
No
No
Si
16
32. In HbA , la sole tasche della proteina che legano il glucosio che hanno sia una Lisina ( che agisce come
un nucleofilo ) che una istidina basica ( che catalizza l’apertura dell’anello ) sono queste associate con Lisina
17, 59 , 61 , 66 , e 82 . Da notare , questi sono i residui prevalenti di Lisina che sono conosciuti nella
reazione di glicazione .
Ci sono molte altri residui di Lisina in HbA che legano il glucosio , ma non portano alla glicazione . Perché ?
Per procedere allo stadio covalente del processo di glicazione , una tasca della proteina deve :
a. Legare α- o β-piranosio
b. Possedere un lisina localmente ( o l’N-terminale di un amminoacido come la valina ) che possa agire
come nucleofilo
c. Possedere una base localmente come una istidina per legare α- o β-piranosio per formare
l’isomero AA (anello aperto ) del glucosio che diventa un elettrofilo reattivo .
Se la tasca di una proteina lega il glucosio e ha una lisina localmente ma non una istidina , non potrà
procedere allo stadio covalente di glicazione perché quale dei seguenti requisiti mancano :
La base per generare AA elettrofilo o il nucleofilo ?
AA elettrofilo
Se la tasca di una proteina lega il glucosio e ha una istidina localmente ma non una lisina sempre
localmente , non può procedere lo stadio covalente della glicazione perché è mancante quale dei seguenti :
La base per generare AA elettrofilo o il nucleofilo ?
Il nucleofilo
Perché sono necessari sia una istidina che una lisina nella stessa tasca che lega il glucosio in modo che
avvenga la glicazione ?
La presenza della Istidina è necessaria perché attraverso la catalisi basica genera l’apertura dell’anello
dello zucchero . E’ necessaria la presenza della Lisina per avere l’attacco nucleofilo allo zucchero dopo
l’apertura dell’anello e generare la aldimmina /base di Schiff per proseguire nel processo di glicazione .
33. Sommario del Meccanismo : Il meccanismo seguente è una potenziale descrizione del processo di
glicazione . Rispondete alle domande che si basano su questo meccanismo .
17
Nel riquadro A:
Lo zucchero è legato covalentemente o non covalentemente ?
L’iniziale legame dell’ anello dello zucchero è nella forma aperta o chiusa ?
Nell’iniziale legame dello zucchero c’è un carbonio carbonilico ?
Nell’iniziale legame lo zucchero è un buon elettrofilo tale che la lisina può
attaccare come nucleofilo e procedere nel processo della glicazione ?
Quale ruolo gioca l’istidina quando interagisce con lo zucchero ?
( acido o base )
Quale ruolo gioca l’acido aspartico ( indicato come asp ) ?
Suggerimento : L’acido aspartico rende l’istidina più o meno basica ?
Non legato covalentemente
Anello chiuso
No
No
Base
Rende l’Istidina più basica
Nel riquadro B :
Lo zucchero è legato in modo covalente o non covalente ?
Lo zucchero legato è AA o AC ?
Non legato covalentemente
Anello aperto AA
18
Nel legame iniziale lo zucchero è un buon elettrofilo tale che
la Lisina può attaccare come un nucleofilo e procedere alla glicazione ?
Si
C’è un buon nucleofilo vicino ?
Si
Se così , che cosa può agire da nucleofilo ?
Un residuo di Lisina
Nel riquadro C :
Lo zucchero è legato in modo covalente o non covalente ?
Sta avvenendo un attacco nucleofilo ?
Chi sta agendo come nucleofilo ?
Chi sta agendo come un elettrofilo ?
Non legato covalentemente
Si
Un residuo di Lisina
Il glucosio ad anello aperto AO
Nel riquadro D :
Quale nuove specie si sono formate dall’ attacco nucleofilo ?
Aldimmina / base di Schiff
Lo zucchero è legato in modo covalente o non covalente ?
Legato in modo covalente