Enzimi")
Seconda Università degli Studi di Napoli
DiSTABiF
Anno Accademico 2015-16
Corso di Laurea Magistrale in
SCIENZE DEGLI ALIMENTI E DELLA NUTRIZIONE UMANA
Insegnamento di
BIOCHIMICA e BIOTECNOLOGIE
degli ALIMENTI
Prof. Augusto Parente
Lezione 14
GLI ENZIMI: UN Po’ DI STORIA
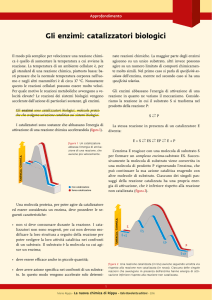
Gli ENZIMI: catalizzatori biologici.
Gli enzimi sono molecole specializzate che presentano una attività
catalitica.
In quanto catalizzatori fanno aumentare la velocità delle
reazioni facendole avvenire in tempi compatibili con le esigenze
cellulari.
Essi non modificano l’equilibrio chimico, ma agiscono
esclusivamente abbassando l’energia di attivazione delle reazioni,
facendo sì che un numero molto più alto di molecole di substrato
superi la barriera energetica per trasformarsi in prodotto.
Aspetti termodinamici
L’energia di attivazione è l’energia necessaria per
arrivare ad un composto intermedio ad alta energia
e bassa stabilità, noto come complesso attivato
Catalizzatori
La natura proteica della gran parte degli enzimi (ad eccezione dei ribozimi) fa sì
che essi abbiano caratteristiche positive rispetto ai catalizzatori inorganici.
Gli enzimi sono EFFICIENTI
Si consideri la reazione
2H2O2
Catalasi
2H2O + O2
La catalasi fa aumentare la velocità della reazione di dismutazione del perossido
d’idrogeno di un fattore pari a 108, mentre un catalizzatore inorganico come il platino la
fa crescere di un fattore pari a 104.
Inoltre le reazioni catalizzate da enzimi non richiedono condizioni drastiche, sono
specifiche e sono regolabili.
=
Idrolisi di una ammide
O
R-C-NH2
(es. legame peptidico)
Catalizzatore
pH
Tempo
Temperatura
Legami
rotti
Nessuno
Neutro
(pH 7,0)
Infinito
Ambiente
Nessuno
H+
< 1,0
110 °C
Tutti
OH-
> 13,0
Giorni
24-72 ore
(1440-4320 min)
Enzima proteolitico Neutro
(pH 7,0)
Minuti
(10-60 min)
37 °C
Alcuni
Rapporto tra il
tempo della catalisi
inorganica
144 (432)
( H+/OH-) e quella
enzimatica.
Gli enzimi sono SPECIFICI
Essi hanno una elevata specificità sia di reazione che di
substrato
La specificità è data dalla presenza del sito attivo
Chimotripsina
SITO ATTIVO
E’ la regione della proteina in cui si lega il substrato (e i cofattori se
esistono).
- Sito di legame o di binding (KM);
- Sito catalitico: costituito dai residui amminoacidici che partecipano
direttamente alla formazione/rottura dei legami interessati nella
catalisi.
1- Il sito attivo è costituito da una tasca (o fenditura ) tridimensionale;
2- il sito attivo occupa una parte relativamente piccola del volume totale di un
enzima;
3- I siti attivi sono dei microambienti unici;
4- I substrati si legano all’enzima mediante un certo numero di interazioni
deboli;
5- La specificità del legame dipende dalla disposizione degli atomi nel sito
attivo.
TRE ENZIMI PROTEOLITICI. Tutti e tre tagliano il legame peptidico, ma su substrati diversi.
Esempio di catalisi covalente.
Sito
catalitico
Ser-195
Sito di
binding
Residui acidi: Asp
Residui relativamente
idrofobici: Gly, Val, Met, Phe
His 57
Asp 102
Ser 195
Residui con catena laterale idrofobica
piccola: Val
Gli enzimi sono REGOLABILI
L’attività enzimatica può essere modulata sia positivamente che negativamente, da
EFFETTORI in funzione delle esigenze cellulari, come nel caso degli enzimi allosterici.
Modelli di legame enzima-substrato
Sono state formulate varie ipotesi sulla maniera con cui il substrato si adatta
perfettamente al sito attivo dell’enzima mediante il riconoscimento
chemio- e stereo-selettivo
Il sito di legame dell’enzima può
già presentare la forma adatta per
sistemare il substrato grazie ad
una complementarità geometrica
(come la chiave si adatta
perfettamente alla serratura)
Il sito di legame sarebbe quindi rigido e ciò contrasta con i suoi cambiamenti
dinamici alla base della catalisi enzimatica
Modelli di legame enzima-substrato
Adattamento indotto
In seguito all’interazione con il substrato il sito di legame dell’enzima subisce
un cambiamento conformazionale per meglio adattarsi alla geometria del
substrato
In tal modo si stabilizza il legame del substrato grazie all’instaurarsi di legami
deboli. La reversibilità di tali legami è alla base della dinamicità
dell’interazione, perché, dopo la sua trasformazione in prodotto, il substrato
deve essere allontanato dal sito di legame.
Adattamento indotto nell’esochinasi
Il legame del substrato al sito attivo dell’esochinasi provoca una
modificazione conformazionale dell’enzima che in tal modo adatta il suo sito
attivo al substrato.
Infatti i reagenti devono entrare in collisione con una corretta orientazione
(fattore probabilistico; costante di Arrhenius A) e con una sufficiente energia
(energia di attivazione E)
- Significativo abbassamento del livello energetico dello stato di transizione;
- Esatto orientamento delle molecole reagenti
Formazione del prodotto
La cinetica enzimatica
- Le reazioni chimiche possono essere monomolecolari, bimolecolari e pseudomolecolari
- A seconda di come la VELOCITA’ viene influenzata dalla concentrazione dei reagenti, una
reazione NON enzimatica è direttamente proporzionale alla concentrazione di S :
- 1° ordine:
- 2° ordine:
S
k
S1+S2
v= -
P
k
P
d[S]
dt
= d[P] =k[S]
dt
v= k[S1]x[S2]
- Ordine 0: rispetto ad S1
se
rimanendo virtualmente costante durante la reazione.
S1 >>S2
Complessivamente la reazione viene definita di pseudo primo ordine
REAZIONE ENZIMATICA
k
S
Substrato
P
Prodotto
Per determinare la velocità della reazione :
V
=
V
=
-
d[S]
dt
(- diminuzione di S)
d[P] (+ formazione di P)
dt
Le unità di velocità sono :
Moli x litro x s: M-1 s-1
oppure
Moli x litro x min
Velocità della reazione (Vo)
Intuitivamente, se aumenta [E], proporzionalmente aumenta vo
Concentrazione di enzima [E]
E così, se aumenta [S], proporzionalmente dovrebbe aumentare vo
Velocità della reazione (Vo)
Concentrazione di substrato [S]
Velocità della reazione (Vo)
Al variare di S, con [E] costante, non si ha la stessa proporzionalità, ma……
Concentrazione del substrato [S]
In realtà sperimentalmente si vede che per
S+E
P+E
La velocità V varia secondo una curva iperbolica. Tende cioè a raggiungere un plateau
all’aumentare di [S].
Nel 1913 Michaelis e Menten proposero un modello
k3
k2
Dove
k4
v= k3 [ES]
K3 = kcat
v= kcat [ES]
Risolvendo, e per [Etot]x kcat = Vmax
v
Equazione di Michaelis e Menten
Modello di Michaelis e Menten
[E]= costante
In una prima fase si forma il complesso
enzima-substrato (ES) ed in una
seconda fase, che generalmente è lo
stadio più lento della reazione, si ha la
formazione del prodotto (P) con
liberazione dell’enzima.
Catalisi cooperativa
Grafico di Lineweaver-Burk
Numero di turnover
Enzima
Vmax= [Etot]x kcat
N. Turnover Reazione
(sec-1)
Acetilcolinesteresi 103
Chimotripsina
102 -103
Tripsina
102 -103
Ribonucleasi
102
Aldolasi
102
Anidrasi carbonica
104 -105
Catalasi
107
HCO3- + H+
2H2O2
+ 2H2O
O2
H2O + CO2
INIBIZIONE
A
Inibizione
IRREVERSIBILE
B
Diisopropilfluorofosfato
Inibitore competitivo
Inibitore non competitivo
Effetto della temperatura
Temperatura ottimale
Un aumento di 1,7-2,5
volte ogni 10°C
Plot di Arrhenius
Denaturazione termica
Effetto del pH
pH ottimale
MODIFICHE COVALENTI: ATTIVAZIONE DEGLI ZIMOGENI
Chimotripsinogeno
Enteropeptidasi
2
1
Tripsinogeno
Chimotripsina
Tripsina
Proelastasi
4
3
Procarbossipeptidasi
Carbossipeptidasi A
1- L’attivazione degli enzimi avviene laddove servono;
2- Il numero di molecole attivate aumenta geometricamente
1:10
10:100
100:1000
Elastasi
Attivazione della chimotripsina
245
Chimotripsinogeno 1
S
S
S
S
Tripsina
Chimotripsina p
1
245
15 16
S
1
S
S
15 16
S
S
Ser14-Arg15
Thr147-Asn148
146 149
S
S
Cambi conformazionali
a-Chimotripsina
S
245
 Enzimi")