Fusi neuromuscolari
Ipotesi su:
Algie correlabili
ai riflessi
innescati dai F.N.M.
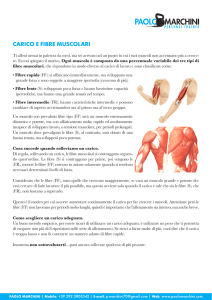
I Fusi Neuromuscolari sono delle strutture altamente specializzate che si trovano diffusamente distribuite
nel parenchima muscolare dei muscoli scheletrici. Sono stati identificati morfologicamente nel 1893 da
Ruffini e kerschner ed inclusi tra i recettori della sensibilità propriocettiva introdotta da Sir Sherrington
nel 1906, inizialmente insieme solo agli Organi tendinei del Golgi e, successivamente, anche a recettori
tattili, pressori, dolorifici ecc… Consistono in fascicoli di fibre muscolari specializzate dai 3 ai 10 mm. di
lunghezza racchiuse in una capsula connettivale affusolata che si espande nella porzione centrale ad
accogliere un fluido di sostanza gelatinosa simile all’umor vitreo del globo oculare. Schematicamente
possono quindi essere suddivisi in 3 zone: una equatoriale sensitiva e due distali motorie (fibre muscolari).
Sono forniti di abbondanti vasi sanguigni e presentano una ricchissima e complessa innervazione (motoria e
sensoriale), infatti si calcola che più della metà delle fibre componenti un nervo muscolare porti segnali dal
fuso al midollo spinale e viceversa. I Fusi neuromuscolari forniscono informazioni basilari per ciò che
riguarda la motricità in quanto si occupano di informare il Sistema di: propriocettvità e posizione, stato di
lunghezza, di tensione e velocità di accorciamento del muscolo; insieme agli organi tendinei del Golgi
situati nel tendine che però sono solo recettori di tensione e non hanno capacità motorie a differenza dei Fusi.
Le fibre che li compongono sono fibre modificate, più sottili di quelle muscolari vere e proprie (extrafusali)
e vengono definite intrafusali; queste si dividono in due gruppi:
1) fibre a catene di nuclei: 8-10 per ogni fuso, in cui i nuclei sono disposti in fila nella porzione mediana.
2) fibre a sacco di nuclei: 3-5 per ogni fuso, le quali presentano nella porzione mediana, che appare rigonfia,
un accumulo di circa 100 nuclei. Queste ultime possono essere suddivise in ulteriori due sottotipi molto
differenti per proprietà strutturali, biochimiche e meccaniche:
1) statiche e
2) dinamiche.
Sia le fibre a catena di nuclei, che quelle a sacco di nuclei statiche sono innervate nelle loro porzioni polari da
fibre motorie provenienti dai motoneuroni gamma delle corna anteriori del midollo spinale: le fibre
gammaII statiche (processi lenti). Quelle a sacco di nuclei dinamiche sono innervate dalle fibre gammaI
dinamiche (processi rapidi). Le fibre sensitive che originano da terminazioni a spirale avvolte sulla porzione
equatoriale di tutte le fibre intrafusali appartengono al gruppo IA di Lloyd, hanno diametro di 12-20 micron,
alta velocità di conduzione e sono definite fibre afferenti primarie. Quelle che provengono
prevalentemente da terminazioni a spirale avvolte sulla porzione paracentrale delle fibre a catena di nuclei
appartengono al gruppo II di Lloyd, hanno diametro e velocità di conduzione inferiore e vengono definite
fibre afferenti secondarie. A queste si aggiungono, distribuite quasi esclusivamente sulle fibre a sacco di
nuclei, le terminazioni a fiorami.
La capsula fusale che, ai poli, è in continuità con l’endomisio, viene a disporsi in parallelo alle fibre
extrafusali, per cui ogni allungamento del muscolo produce un distanziamento delle spire della
terminazione nervosa avvolta sulla porzione centrale delle fibre intrafusali (in cui si trova il fluido
gelatinoso, che permette spostamenti indipendenti tra loro delle varie fibre). Essendo i Fusi recettori di
primo tipo, sono in grado di rispondere a uno stimolo con un Potenziale Generatore avente le caratteristiche
del potenziale locale, il quale, se è sufficientemente ampio da depolarizzare fino al livello di soglia la zona
della fibra corrispondente al primo nodo di Ranvier, può generare una scarica di potenziali d’azione la cui
frequenza sarà proporzionale all’ampiezza dell’area stimolata e all’intensità dello stimolo stesso. Tramite le
variazioni di scarica delle fibre secondarie il S.N.C. riceve informazioni sull’entità dell’allungamento
muscolare e tramite le primarie sulla velocità dell’allungamento. L’innervazione gamma produce una
contrazione localizzata alle estremità polari delle fibre intrafusali, che a sua volta determina uno stiramento
della loro parte equatoriale, dove sono avvolte le terminazioni nervose. In questo modo la detensione del
fuso prodotta dalla contrazione delle fibre extrafusali ad opera dei motoneuroni alfa, può essere
compensata dagli effetti opposti prodotti dalla contemporanea attivazione dei motoneuroni gamma. Così
facendo la sensibilità del recettore può rimanere costante ad ogni livello di contrazione muscolare. Il
controllo di tale sensibilità, operato dal sistema gamma, è in grado di esaltare selettivamente la sensibilità
statica o dinamica del fuso, attraverso i due tipi di fibre gamma che l’innervano. Gli impulsi provenienti dai
Fusi raggiungono il midollo spinale ed eccitano i motoneuroni alfa e quelli dei muscoli agonisti con una
trasmissione monosinaptica.
Quindi il brusco allungamento di un muscolo provoca , mediante l’attivazione dei propriocettori fusali e
tramite fibre afferenti IA di largo calibro, una contrazione riflessa che tende a riportare il muscolo alla sua
lunghezza originaria (riflesso miotatico fasico - feedback di controllo della lunghezza, ad es.: riflesso
patellare, achilleo, ecc..).
Il riflesso miotatico tonico è particolarmente presente nei muscoli che si oppongono alla forza di gravità, la
quale tende ad allungarli, che svolgono un ruolo primario nel mantenimento della postura (stimolo
continuo-risposta tonica). In questi casi è la componente statica del fuso ad essere stimolata: le fibre
afferenti di tipo IA e II attivano interneuroni centrali che scaricano su motoneuroni di piccolo diametro.
Queste unità motorie toniche sono in grado di mantenere per periodi prolungati scariche a bassa frequenza.
Esistono centri sopraspinali di controllo dei gamma motoneuroni tonici, posti nella formazione reticolare e
nel complesso dei nuclei vestibolari, che ottimizzano le funzioni propriocettive dei Fusi. Nel caso di
movimenti volontari la coattivazione alfa-gamma in contemporanea rende i Fusi praticamente insensibili
alla contrazione impedendo che l’attività di scarica diminuisca in funzione dell’accorciamento delle fibre
extrafusali, estendendo le fibre intrafusali e mantenendo costante la scarica. Ne segue che i Fusi
neuromuscolari perdono il ruolo di rilevatori di lunghezza assumendo quello di segnalatori di errore
nell’esecuzione, potendo così “servo-assistere” il movimento volontario.
Definita la composizione ed il funzionamento dei Fusi, cercherò di ipotizzare le cause di fenomenologie
algiche muscolari dovute a situazioni “conflittuali” tra schemi motori, innati e non, ed il controllo muscolare
operato dai Fusi stessi.
Schematicamente: le afferenze della sensibilità propriocettiva (derivanti da Fusi, org.del Golgi e capsule
articolari: vie spino-cerebellari), tattile e pressoria (vie del cordone posteriore e fasci spino-talamici
anteriori), termica e dolorifica (fasci spino-talamici laterali), ascendendo fanno tappa, insieme alle
afferenze vestibolari, uditive, oculari... provenienti da vari distretti della testa, in centri e nuclei superiori in
cui possono innescare nuove risposte riflesse o proseguire verso cervelletto o talamo e corteccia. Quindi il
controllo della motilità somatica si articolerà su diversi livelli che andranno da semplici riflessi a schemi di
movimento involontari fino a volontari complessi. Si avrà la pianificazione quando, presa la decisione
cosciente di effettuare un movimento, l’informazione partirà dalla corteccia, dal lobo frontale, e
raggiungerà le aree associative motorie che a turno la invieranno a cervelletto e nuclei cerebrali.
All’effettuazione del movimento le aree associative manderanno impulsi alla corteccia motoria primaria. Il
feedback proveniente dal cervelletto e dai nuclei modificherà il comando e le efferenze lungo le vie
extrapiramidali (vestibolo-spinali, tetto-spinali, reticolo-spinali, e rubro-spinali) effettueranno degli
aggiustamenti automatici della posizione corporea e del tono muscolare; per cui il controllo dei
motoneuroni avverrà volontariamente attraverso le vie piramidali (cortico-spinali) ed involontariamente,
ma anche legato a schemi motori, attraverso le vie extrapiramidali.
Cercherò di analizzare come le risposte generate da cervelletto e nuclei della base siano sì automatiche ed
involontarie, ma sufficientemente elaborate da entrare in conflitto con movimenti e perturbazioni
dell’organismo “non previsti”. Abbiamo considerato come i Fusi possono controllare più o meno
direttamente il comportamento involontario di muscoli scheletrici agonisti attraverso i riflessi miotatici
fasico e tonico, che coinvolgono fibre sensitive afferenti legate monosinapticamente a motoneuroni
efferenti alfa e gamma a livello del midollo spinale.
Il riflesso miotatico tonico è un sistema di controllo che serve per la regolazione del tono posturale (per
mantenere la postura ideale - Atteggiamento Posturale Primario: posizione anatomica in stazione eretta,
faccia rivolta in avanti arti superiori avvicinati ai fianchi), che cambia a seconda della posizione e tende ad
annullarsi se si è sdraiati in quanto la gravità ha poca influenza in questa posizione. L’A.P.P. può essere
modificato da impulsi squilibrati che possono essere causati da vari fattori: alterazione della sensibilità,
gesti o posture squilibrati ripetuti, atteggiamenti antalgici, limitazioni traumatiche o funzionali… Nel caso
in cui uno squilibrio diventi egemone, nella relazione tra i vari segnali che regolano la postura, ne risulterà
una nuova situazione di equilibrio che comprenderà anche gli adattamenti all’impulso squilibrante e ne
deriverà una nuova postura che verrà comunque considerata ideale pur non essendolo. Fintanto che rimarrà
il segnale di squilibrio proveniente dai recettori, la postura ideale rimarrà quella nuova (adattata). Ne
deriveranno dei compensi dovuti alla ricerca automatica di una situazione antalgica da parte dei muscoli
antigravitari con vocazione posturale (ricchi di Fusi, dal duplice ruolo: mantenimento della posizione e
generazione di movimento). Questi muscoli saranno assoggettati ad un’attività contrattile continua, per cui
la loro fisiopatologia sarà l’ipertonia ed essendo la muscolatura tonica una mucolatura lenta e poco
affaticabile la sua patologia non sarà di debolezza, ma di accorciamento e retrazione (rigidità). Quindi il
protrarsi nel tempo di una postura squilibrata produrrà un’alterazione della viscosità di alcuni gruppi
muscolari responsabili del mantenimento di detta postura anche quando l’impulso squilibrante verrà a
mancare.
Essendo questi muscoli all’interno di catene muscolari toniche, la loro perturbazione o l’accorciamento
genereranno delle nuove zone di compenso statico anche lontano dalla zona dello squilibrio primario, nelle
quali si potranno avere altre manifestazioni di dolore.
Anche l’apparato stomatognatico, in cui sono presenti numerosi ed importanti recettori (dentali,
gengivali...) in grado di registrare anche le minime forze e spostamenti e di trasferirle al S.N.C. tramite il
nervo trigemino, possiede muscoli ricchi di Fusi (elevatori della mandibola..). Ne è dimostrazione il fatto
che, abbassando la mandibola si ha una stimolazione dei Fusi stessi che provoca contrazione attraverso il
riflesso da stiramento. Questo apparato è quindi soggetto a scompensi, proprio a causa di questa notevole
sensibilità, che si possono ripercuotere sulla globalità della postura.
Il riflesso miotatico fasico risponde a bruschi allungamenti del muscolo con una contrazione riflessa
immediata, che nel caso di contemporanee efferenze relative a quadri motori, potrebbero portare a
situazioni di conflitto. Se ad esempio stessimo svolgendo un qualsiasi compito ed alle nostre spalle
sentissimo sbattere una porta, automaticamente potremmo ruotare la testa in quella direzione, producendo
un brusco stiramento dei muscoli estensori, posteriori del collo controlaterali, a cui seguirebbe una
contrazione riflessa, dovuta ai Fusi (riflesso miotatico fasico), non seguita da un effettiva controrotazione
del capo. Avremmo quindi l’attivazione da parte dei Fusi della contrazione delle fibre extrafusali attraverso i
motoneuroni alfa in contrazione eccentrica, non protetta dall’attivazione degli organi tendinei del golgi che
hanno bisogno di tempi di stimolazione più lunghi, con conseguente possibile contrattura, imbrigliamento
più serrato delle fibre di actina e miosina e addensamento di tessuto connettivo. Così come un calcio a vuoto
nella previsione di colpire un pallone, sarebbe una deviazione imprevista da uno schema motorio
precostituito, in cui avremmo un rapido allungamento dei muscoli antagonisti, cui seguirebbe la
contrazione riflessa, ma senza la flessione in risposta… Ricordiamo che l’attività dei Fusi è anche
influenzata da molteplici fattori, uno fra tutti, le efferenze dalla formazione reticolare, responsabile delle
funzioni di attenzione e veglia, per cui lo stato emotivo potrà fare variare l’attività muscolare sia fasica sia
posturale. Un altro fattore è la temperatura, che può modificare il funzionamento dei motoneuroni gamma: il
calore ne riduce l’attività producendo il rilassamento del muscolo, al contrario del freddo che ne aumenta la
rigidità esponendolo a maggior rischio di traumi.
20 settembre 2006
Federico Wang Matricola 178445