PIANO DI STRUTTURA DELLA PIANTA
Con la germinazione riprende lo sviluppo vegetativo della pianta, interrottosi con la maturazione
del seme.
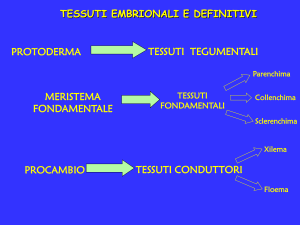
Abbiamo già visto che durante l’embriogenesi si formano dei MERISTEMI PRIMARI (protoderma,
procambio e meristema fondamentale). Man mano che l’embrione matura e l’asse radicegermoglio si accresce, gli apici del germoglio e della radice si distanziano l’uno dall’altro: le cellule
intermedie cessano di dividersi, e le cellule meristematiche restano localizzate agli apici del
germoglio e della radice.
Questo tipo di organizzazione resta anche nella pianta adulta. La presenza di meristemi primari
per tutta la vita conferisce alla pianta la possibilità di un ACCRESCIMENTO INDEFINITO, in
contrasto con quello che avviene negli animali. L’accrescimento indefinito consente alla pianta di
rispondere all’ambiente mediante crescita e differenziamento, che sono il “sostituto immobile” del
comportamento.
Mentre l’intera pianta ha un accrescimento indefinito, alcuni organi, come le foglie, l’ipocotile ecc.
hanno un accrescimento definito (v. grafico).
Le cellule meristematiche sono cellule “indifferenziate” che sono specializzate per una divisione
ordinata, che stabilisce il disegno di sviluppo della pianta (quando la divisione cellulare non è
ordinata, come per esempio in cellule in coltura, si forma un “CALLO”).
I meristemi si possono distinguere in
MERISTEMI APICALI
agli apici del germoglio e agli apici delle radici. Le cellule che li
compongono sono dette cellule iniziali, e hanno divisioni lente, che producono cellule derivate
mitoticamente attive. I meristemi apicali sono in genere già presenti nel seme.
MERISTEMI PRIMARI
protoderma, procambio e meristema fondamentale. Anch’essi sono in
genere già presenti nel seme. Determinano l’accrescimento in lunghezza della pianta, e danno
origine ai tessuti primari e al corpo primario della pianta.
MERISTEMI SECONDARI o LATERALI determinano l’accrescimento in spessore della pianta,
che è coordinato con quello in lunghezza, e danno origine a tessuti secondari e al corpo
secondario della pianta. Derivano da cellule già distese e parzialmente differenziate, o che
comunque avevano sospeso temporaneamente la divisione, le quali riprendono a dividersi.
Le nuove cellule prodotte dai meristemi possono riprendere il ciclo cellulare (cioè restare
meristematiche) o prendere la strada del differenziamento, specializzandosi per svolgere
determinate funzioni, dando quindi origine a diversi tipi di TESSUTI. Queste cellule
progressivamente si distendono e si differenziano, per cui a partire dal meristema avremo un
gradiente di maturazione lungo il quale le cellule più giovani sono quelle più vicine al meristema,
mentre man mano che ci si allontana dal meristema si hanno cellule sempre più differenziate.
Un aspetto importante nello sviluppo della pianta, che è in relazione con la presenza della parete e
l’immobilità della cellula, è dato dal piano di divisione. Considerando l’asse della pianta, una
divisione può essere
TRASVERSALE se il piano di divisione è perpendicolare all’asse dell’organo. Questa divisione
porta ad un aumento in lunghezza;
LONGITUDINALE se il piano di divisione è parallelo all’asse. Una divisione longitudinale può
essere
o PERICLINALE o TANGENZIALE se il piano di divisione è parallelo alla superficie
dell’organo. In questo caso aumenta il numero degli strati di cellule, e l’organo si
accresce in spessore.
o ANTICLINALE o RADIALE se il piano di divisione è perpendicolare alla superficie
dell’organo. In questo caso aumenta il numero di cellule della circonferenza, e si
accresce la superficie dell’organo.
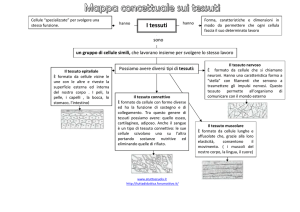
I TESSUTI
La formazione di tessuti specializzati è in relazione con l’emersione delle piante dall’acqua e con la
colonizzazione dell’ambiente subaereo nel corso dell’evoluzione.
L’ambiente subaereo rispetto all’ambiente acquatico consente un miglioramento degli scambi
gassosi e delle condizioni di illuminazione. Tuttavia crea il problema della disidratazione (perché il
potenziale d’acqua nell’aria è per lo più molto più basso che nella pianta), del rifornimento di
acqua, che deve essere sottratta al terreno, del sostegno. Inoltre l’ambiente subaereo è molto più
soggetto a rapide variazioni.
Per risolvere questi problemi le piante hanno dovuto sviluppare degli organi e dei tessuti specifici.
Innanzi tutto hanno dovuto sviluppare una parte che cresca in profondità nel terreno per il
rifornimento di acqua e per l’ancoraggio della pianta stessa (RADICE). La radice, non essendo
esposta alla luce, non può fotosintetizzare e deve essere rifornita di sostanze organiche dagli
organi fotosintetizzanti (FOGLIE), che d’altra parte devono essere rifornite d’acqua dalla radice.
Per questo si forma un organo di conduzione (FUSTO), che serve anche per il sostegno della
pianta. Il corpo della pianta differenziato in radice, fusto e foglie è detto CORMO, e le piante che lo
presentano sono dette CORMOFITE, in contrasto con le TALLOFITE (piante a tallo, come per
esempio alghe e funghi), che hanno il corpo meno organizzato.
A loro volta gli organi sono costituiti da tessuti specializzati: la protezione della pianta dalla
disidratazione e da fattori biotici e abiotici richiede il differenziamento di tessuti TEGUMENTALI; il
sostegno della pianta richiede tessuti MECCANICI; la connessione nei due sensi fra radici e foglie
richiede tessuti CONDUTTORI; i tessuti specializzati per la fotosintesi o per la funzione di riserva
sono detti PARENCHIMATICI.
Un TESSUTO è un aggregato di cellule simili per struttura e funzione, e connesse fra loro.
I tessuti si possono distinguere in
MERISTEMATICI e ADULTI
PRIMARI e SECONDARI (secondo l’origine)
I tessuti adulti possono essere distinti secondo la funzione in:
TEGUMENTALI
PARENCHIMATICI
MECCANICI
CONDUTTORI
SECRETORI
Per che cosa si differenziano i vari tipi di tessuti e come si possono riconoscere e distinguere l’uno
dall’altro al microscopio?
posizione del tessuto nell’organo
forma e dimensioni delle cellule
presenza o meno di spazi intercellulari
presenza o meno del protoplasto
presenza o meno di plasmodesmi e loro dimensioni
spessore e composizione della parete
presenza o meno e grado di sviluppo del vacuolo
sostanze presenti nel vacuolo (pigmenti, inclusi solidi)
tipo di plastidi
MERISTEMI
Meristemi primari
I meristemi primari sono costituiti da cellule di piccole dimensioni (5-15 m), più o meno
isodiametriche o poliedriche, con citoplasma denso, molti ribosomi (intensa sintesi proteica),
nucleo grosso e centrale (alto rapporto nucleo/citoplasma), parete sottile, solo primaria, proplastidi,
vacuoli piccoli o assenti, e sono prive di spazi intercellulari.
Meristemi secondari
Le cellule dei meristemi secondari sono già distese, e quindi rispetto a quelle dei meristemi primari
sono di dimensioni maggiori e vacuolate; il nucleo è generalmente periferico. Hanno anch’esse
parete sottile e sono prive di spazi intercellulari. Riprendono la loro attività meristematica dopo
essersi differenziate o comunque avere cessato di dividersi per un certo tempo.
Le cellule dei meristemi primari derivano direttamente dai meristemi primari dell’embrione. Tuttavia
è possibile che da cellule già distese o in parte differenziate si riformi un meristema primario per
riassorbimento del vacuolo, sintesi di citoplasma, migrazione del nucleo verso il centro della
cellula, ripresa della divisione cellulare. Questo avviene per esempio quando si rigenera un
primordio di radice da una talea di fusto.
TESSUTI TEGUMENTALI
I tessuti tegumentali sono tutti caratterizzati da cellule piuttosto appiattite e prive di spazi
intercellulari, che formano quindi uno o più strati continui.
I tessuti tegumentali si possono suddividere in:
tessuti tegumentali primari: EPIDERMIDE che riveste le foglie e il fusto primario
RIZODERMA che riveste la radice primaria nella zona più giovane
ESODERMA che subentra al rizoderma nella radice
ENDODERMA
tessuti tegumentali secondari: SUGHERO
Epidermide
L’epidermide è per lo più costituita da un unico strato di cellule appiattite e a mutuo contatto. Le
cellule possono avere varie forme, in relazione al tipo di pianta e all’organo che rivestono: sono per
lo più allungate nelle foglie allungate o sul fusto, sono spesso lobate nelle foglie larghe. Le pareti
possono essere più o meno spesse; la parete tangenziale esterna è cutinizzata, e può presentare
anche una cuticola e delle cere. La parete può essere anche silicizzata, come nelle graminacee.
Le cellule dell’epidermide tipicamente non contengono cloroplasti, ma possono contenere pigmenti
(soprattutto antociani e flavoni), come nei petali o nei frutti o in certe foglie di colore violaceo.
L’epidermide presenta anche appendici di vario genere dette TRICOMI o PELI, e STOMI
Tricomi
I peli possono essere uni- o pluri-cellulari, ed essere costituiti da cellule vive o da cellule morte.
I peli morti hanno principalmente funzione di protezione dalla disidratazione, in quanto, costituendo
una specie di feltro sulla superficie della foglia, limitano la traspirazione e riflettono la radiazione
solare. In altri casi i peli morti facilitano il trasporto dei frutti o dei semi, come nel caso del cotone o
del pioppo.
I peli vivi possono avere diverse funzioni: in piante rampicanti come il luppolo o la canapa è
attraverso i peli che il fusto si aggrappa al sostegno; in piante di ambienti tropicali i peli aumentano
la superficie di traspirazione; i peli secretori possono produrre sostanze di vario genere (oli
essenziali, resine, mucillagini, sostanze urticanti (ortica); in alcune piante carnivore i peli
intervengono nella cattura degli insetti e nella secrezione di enzimi digestivi.
Stomi
Gli stomi sono delle aperture regolabili delimitate da due CELLULE DI GUARDIA, che hanno la
funzione di consentire e regolare gli scambi gassosi. Viste di faccia (per esempio in una spellatura
dell’epidermide) le cellule di guardia appaiono reniformi (a clessidra nelle graminacee), attaccate
fra loro alle estremità, e delimitano un’apertura detta RIMA stomatica. A differenza delle altre
cellule dell’epidermide le cellule di guardia hanno cloroplasti. Le caratteristiche di queste cellule
fanno sì che la rima si apra quando le cellule sono turgide. Infatti le pareti ventrali (quelle verso la
rima) sono più ispessite e resistenti delle pareti dorsali; le fibrille di cellulosa nelle pareti hanno una
disposizione radiale. Quando le cellule di guardia, per abbassamento del loro potenziale d’acqua,
richiamano acqua dalle cellule vicine, il vacuolo spinge contro le pareti: le loro pareti dorsali, meno
resistenti, vengono spinte in fuori; e le fibrille di cellulosa, che non possono essere stirate,
trascinano in fuori anche le pareti ventrali, che si inarcano, e la rima si apre.
La formazione dello stoma avviene per divisione ineguale di una cellula dell’epidermide: la cellula
più piccola è la cellula madre dello stoma, che si divide a sua volta in due cellule uguali, che si
differenzieranno in cellule di guardia, modificando le loro pareti, chiudendo i plasmodesmi che le
collegano alle cellule annesse e infine idrolizzando la lamella mediana a livello della rima
stomatica.
Funzioni dell’epidermide
L’epidermide è il tessuto della pianta che è a diretto contatto con l’ambiente esterno, ed è il primo
tessuto che può venire a contatto con altri organismi. Ha quindi funzione di protezione in generale,
e in modo specifico di protezione dalla disidratazione, ma anche di difesa da altri organismi viventi
(parassiti, predatori), o funzione vessillare (nei fiori, nei frutti).
Rizoderma, esoderma ed endoderma saranno trattati nella struttura della radice; il sughero nella
struttura secondaria.
TESSUTI PARENCHIMATICI
Le cellule parenchimatiche possono avere forma e dimensioni varie, hanno la parete generalmente
sottile, spesso limitata alla parete primaria, e sono ricche di spazi intercellulari. Sono nel
complesso poco differenziate e possono riprendere l’attività meristematica (per la rigenerazione o
la cicatrizzazione di ferite, o per dare origine a meristemi secondari).
Possono svolgere diverse funzioni:
parenchima clorofilliano: nelle parti verdi della pianta, ha cloroplasti particolarmente sviluppati
parenchima di riserva: nei tessuti profondi del fusto e della radice, nei semi. Accumula sostanze di
riserva, principalmente in forma di amido nel fusto e nella radice, in forma di amido, lipidi, proteine,
emicellulose nei semi, e anche sotto forma di sostanze solubili nel vacuolo.
parenchima acquifero: nelle piante grasse o succulente; specializzato per trattenere grandi
quantità di acqua, grazie a mucillagini presenti nel vacuolo e nelle pareti.
parenchima aerifero: in piante di palude o acquatiche. Gli spazi intercellulari sono molto grandi,
tanto da formare canali pieni d’aria, per facilitare gli scambi gassosi e il galleggiamento.
parenchima conduttore: si trova nei raggi midollari ed è costituito da cellule allungate e ricche di
punteggiature, per facilitare il trasporto in senso radiale.
cellule transfer: sono cellule specializzate per un trasporto intenso a breve distanza. Sono
caratterizzate da estese invaginazioni della parete, che aumentano di molto la superficie della
membrana plasmatica. Si trovano per esempio nelle nervature delle foglie vicino ai tessuti
conduttori e in strutture riproduttive come la placenta.
TESSUTI MECCANICI
L’insieme delle pareti cellulari ed il turgore costituiscono per la pianta un sistema generale di
sostegno. Ma per resistere alle sollecitazioni meccaniche del peso del tronco, della chioma e dei
frutti, al piegamento, alla trazione, sono necessari tessuti meccanici specializzati
I tessuti meccanici sono caratterizzati dall’ispessimento della parete, e sono di due tipi:
collenchima: costituito da cellule vive, si trova solo nel corpo primario della pianta ed è il tessuto
meccanico tipico delle parti giovani della pianta, ancora in distensione. Le cellule hanno solo
parete primaria, che è ispessita non uniformemente, ma per esempio solo agli angoli (collenchima
angolare) o sulle pareti tangenziali (collenchima lamellare). Si trova in genere subito sotto
l’epidermide, e specialmente nelle costolature o negli spigoli del fusto. E’ in grado di
accompagnare la crescita per distensione della zona di fusto in cui si trova.
sclerenchima: si può trovare sia nella struttura primaria che in quella secondaria. E’ costituito per lo
più da cellule morte, con parete uniformemente e fortemente ispessita, in genere lignificata. Può
essere costituito da FIBRE, cellule allungate e affusolate, o da SCLEREIDI, cellule di varia forma,
non allungate, che si trovano nel tegumento di molti semi, nell’endocarpo duro, nella polpa delle
pere. Le fibre possono essere lunghe anche molti millimetri, e possono venire utilizzate come fibre
tessili: ad esempio, le fibre del lino hanno pareti solo cellulosiche, quelle della canapa hanno pareti
poco lignificate, e quelle della iuta hanno pareti molto lignificate.
TESSUTI SECRETORI
I tessuti secretori producono metaboliti secondari che vengono accumulati nel vacuolo o liberati
all’esterno della cellula. Possono esserci cellule secretrici isolate o strutture formate da più cellule.
Abbiamo già visto i peli secernenti annessi all’epidermide. Esistono poi i CANALI SECRETORI,
cioè delle cavità tappezzate da cellule che secernono sostanze nel lume del canale stesso (Canali
resiniferi nelle conifere, o canali mucipari nelle Liliacee); i LATICIFERI, costituiti da una o più
cellule allungate e sovrapposte, spesso plurinucleate, che contengono nel vacuolo un liquido
lattiginoso detto làtice, che può contenere molti diversi tipi di sostanze; le TASCHE LISIGENE (per
esempio nella buccia degli agrumi), che si formano per lisi delle cellule che producono gli oli
essenziali, i quali si accumulano nella cavità lasciata libera dalle cellule morte.
Le sostanze prodotte dai tessuti secretori hanno una grande importanza nel rapporto della pianta
con altri organismi e per l’utilizzazione da parte dell’uomo.
TESSUTI CONDUTTORI
I tessuti conduttori, per la funzione che svolgono, sono caratterizzati da cellule di forma allungata,
disposte in file longitudinali, e sono in genere accompagnati da cellule parenchimatiche e da
cellule meccaniche.
I tessuti conduttori sono di due tipi:
tessuto vascolare, che trasporta la “linfa grezza” (acqua, sali minerali ed eventualmente altri
soluti) dalle radici alle foglie
tessuto cribroso, che porta la “linfa elaborata” dalle foglie al resto della pianta.
Tessuto vascolare
Il tessuto vascolare è costituito da cellule che a maturità sono morte, quindi della cellula resta solo
la parete cellulare, che è ispessita e lignificata. Il tessuto vascolare può essere costituito da
TRACHEIDI Una tracheide è una singola cellula allungata, del diametro di circa 10-50 m,
nella quale la parete trasversale è spesso obliqua e fortemente punteggiata
TRACHEE o VASI Una trachea è costituita da una successione di cellule disposte una sopra
l’altra, con le pareti trasversali riassorbite in tutto (perforazione semplice) o in parte
(perforazioni multiple). Ogni singola cellula costituisce un ARTICOLO della trachea. Sono in
genere molto più larghe delle tracheidi (diametro 50-500 m)
Le pareti longitudinali di trachee e tracheidi possono avere una lignificazione più o meno estesa. A
seconda dell’aspetto dell’ispessimento della parete secondaria possono essere ANULATE,
SPIRALATE, SCALARIFORMI, RETICOLATE O PUNTEGGIATE (con punteggiature semplici o
areolate).
Il tipo di elementi che costituisce il tessuto vascolare dipende dalla specie, dall’organo in cui il
tessuto si trova, dal momento in cui si differenzia, ed è influenzato dalle condizioni ambientali.
Relazione tra struttura e funzione. La salita della linfa avviene contro gravità e può dover
raggiungere grandi altezze. Essa avviene per aspirazione dalle foglie a seguito della traspirazione
(perdita di vapore acqueo da parte delle foglie). Perciò la linfa deve trovare meno ostacoli possibili
(cellule allungate, eliminazione del protoplasto, perforazioni o assenza delle pareti trasversali) e le
pareti devono essere in grado di resistere alla depressione, che tenderebbe ad aspirarle in dentro,
riducendo il lume del vaso (pareti ispessite e lignificate). Negli elementi anulati e spiralati
l’ispessimento e la lignificazione della parete sono poco estesi, e la parete è poco resistente, ma
può essere stirata ed ha un’ampia superficie che permette l’ingresso o l’uscita dell’acqua: questi
elementi hanno in genere il lume stretto e sono adatti per la conduzione in zone che sono ancora
in distensione; sono infatti i primi elementi che completano il loro differenziamento. Gli elementi
reticolati, scalariformi e punteggiati hanno invece una lignificazione molto estesa, una forte
resistenza alla depressione, ma non possono essere stirati: perciò il lume può essere più ampio, la
conduzione è più efficiente, ma questi elementi devono portare a termine il loro differenziamento in
zone dove la distensione è finita.
Differenziamento del tessuto vascolare
Il differenziamento comporta una crescita per distensione della cellula, soprattutto in lunghezza per
le tracheidi, e anche in larghezza per gli articoli delle trachee. Si differenzia e si ispessisce in certe
zone la parete secondaria, che poi lignifica; il vacuolo si rompe portando all’autolisi del protoplasto
(morte della cellula), e, nel caso delle trachee, si ha anche la lisi delle pareti trasversali.
Tessuto cribroso
Il tessuto cribroso è costituito da cellule ancora vive a maturità, con il plasmalemma integro, ma
generalmente senza nucleo; inoltre il tonoplasto si rompe, provocando un rimescolamento di succo
cellulare e citoplasma; degli organuli cellulari restano solo il reticolo endoplasmico liscio, alcuni
plastidi e mitocondri, che si dispongono alla periferia della cellula; la parete è solo primaria e in
genere sottile, e non è lignificata.
Il tessuto cribroso è caratterizzato dalla presenza di CRIBRI,cioè di zone in cui sono raggruppati i
pori cribrosi, costituiti da plasmodesmi specializzati, che mettono in comunicazione i citoplasmi di
cellule vicine. A livello dei pori nella parete si trova un cilindro di CALLOSIO, un polimero 1-3 del
glucosio. Anche in questo caso ci sono due tipi di elementi: le CELLULE CRIBROSE, che sono
cellule singole, con aree cribrose sparse nelle pareti; i TUBI CRIBROSI, costituiti da una serie di
cellule sovrapposte, comunicanti fra loro per mezzo di PLACCHE CRIBROSE specializzate, con
pori più ampi, sulle pareti trasversali (oltre ad avere aree cribrose sulle pareti longitudinali). Le
placche cribrose possono essere semplici o composte, e i pori hanno in genere un diametro di 0.11 m, ma nelle placche cribrose semplici delle cucurbitacee (ad esempio zucca) il diametro dei
pori può raggiungere i 10-15 m, e i pori sono quindi ben distinguibili al microscopio ottico. Il
callosio, presente nella parete a livello dei pori, può anche essere deposto in maggiore quantità
nella parete fino ad occludere il poro, soprattutto in seguito a ferita. Un’altra sostanza caratteristica
dei tubi cribrosi di alcune piante è la proteina P, che si dispone nel citoplasma lungo le pareti
cellulari, e in seguito a ferite si dispone a formare dei tappi mucillaginosi in prossimità delle placche
cribrose.
I tubi cribrosi sono accompagnati da CELLULE COMPAGNE, cellule di tipo parenchimatico, che
derivano dalla divisione ineguale della stessa cellula madre che darà origine al tubo cribroso, e
costituiscono un supporto vitale al tubo cribroso stesso.
Il contenuto del tubo cribroso è costituito principalmente da acqua, da un’alta concentrazione di
saccarosio, da amminoacidi, ormoni, vitamine.
XILEMA E FLOEMA
Il tessuto vascolare, insieme al tessuto parenchimatico e al tessuto meccanico che lo
accompagnano, costituisce il LEGNO o XILEMA.
Il tessuto cribroso, insieme al tessuto parenchimatico e al tessuto meccanico che lo
accompagnano, costituisce il LIBRO o FLOEMA.
Nella struttura primaria della pianta, xilema e floema sono disposti affiancati, e costituiscono vari
tipi di FASCI CRIBROVASCOLARI.
FASCIO COLLATERALE
legno e libro sono affiancati, il legno verso l’interno e il libro
verso l’esterno. I fasci sono detti CHIUSI se legno e libro sono a diretto contatto, APERTI
se fra legno e libro resta del tessuto indifferenziato che è detto CAMBIO.
FASCIO BICOLLATERALE
come il fascio precedente, ma con libro disposto anche dalla
parte interna del legno
FASCIO RAGGIATO il legno costituisce una stella con numero variabile di punte e il libro è
disposto fra i raggi della stella di legno
FASCIO CONCENTRICO
PERIFLOEMATICO se il legno costituisce un cilindro, che è
circondato da un anello di libro; PERIXILEMATICO se è un cilindro di libro ad essere
circondato da un anello di legno.