INTRODUZIONE
Il presente modulo del corso di Fisiologia Ambientale non intende affrontare lo sterminato
campo della Fisiologia Comparata ma piuttosto descrivere alcuni aspetti dell'interazione
ambiente-essere vivente, e degli adattamenti che questa interazione ha generato. La prima
parte descrive alcune modalità sensoriali con cui gli organismi analizzano l'ambiente,
soffermandoci principalmente sui meccanismi assenti nell'uomo. In particolare, dopo una
sintetica trattazione dell'evoluzione del Sistema Nervoso, verranno analizzate
l'elettrorecezione, la magnetorecezione, la linea laterale dei pesci e la visione. Nella seconda
parte viene trattato il problema dell'interazione tra organismi e la temperatura ambientale,
analizzando i meccanismi con i quali gli esseri viventi affrontano il problema della
termoregolazione, che risulta altresì connessa con il metabolismo energetico. Questa
trattazione prevede un brevissimo richiamo alla fisiologia comparata del sistema circolatorio e
respiratorio. Vengono infine esaminati alcuni aspetti dell'adattamento all'ambiente marino.
1
COME GLI ANIMALI PERCEPISCONO L'AMBIENTE
Verrano esaminate alcune modalità sensoriali che consentono agli esseri viventi di analizzare
l'ambiente, soffermandoci in particolare su modalità sensoriali assenti nell'uomo. La
trattazione è preceduta da alcuni concetti di base relativi all'evoluzione del sistema nervoso.
Richiami su alcune aspetti del sistema nervoso dell'uomo
Il sistema nervoso dei mammiferi è suddiviso in sistema nervoso centrale e sistema nervoso
periferico. Nell'uomo il sistema nervoso centrale (SNC) è costituito dall'encefalo, racchiuso e
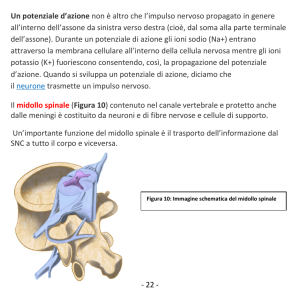
protetto dal cranio, e dal midollo spinale, situato all'interno dei corpi delle vertebre (Fig.1); dal
SNC si dipartono poi i nervi che si distribuiscono a tutto il corpo, costituendo il sistema
nervoso periferico (SNP).
Cervello
Cervello
Tronco
cerebrale
Midollo
spinale
Nervi
periferici
Midollo
spinale
A
B
C
Fig. 1. A e B, visione posteriore del SNC e periferico. C, sezione sagittale del SNC.
L'encefalo comprende il cervello ed, inferiormente, il tronco cerebrale. Questo è costituito
dal mesencefalo, il ponte ed il bulbo. Inferiormente troviamo il midollo spinale.
Il sistema nervoso centrale sembra essere evoluto per aggiunta di parti successive: la parte
più antica è il midollo spinale sopra cui si sono sviluppati il bulbo, il ponte, il mesencefalo ed
infine il cervello, di cui la corteccia rappresenta l'ultimo stadio evolutivo.
Nel SN umano abbiamo circa 1011 neuroni, ciascuno dei quali, in media, ha circa 105
connessioni sinaptiche. L'evoluzione tende a raggruppare insieme, in nuclei ed aree, neuroni
aventi funzione simile (fortunatamente per chi studia l'organizzazione del SN); per esempio,
nella corteccia cerebrale è possibile delimitare delle aree aventi specifiche attività motorie o
sensitive.
Al di sotto dell'encefalo troviamo il midollo spinale, contenuto nei corpi vertebrali della
colonna vertebrale (Fig. 2, A). In fig. 2, B, in alto, sono riportate 2 sezioni trasversali del
2
midollo spinale, effettuate a diversa altezza della colonna vertebrale. All'interno si trova la
sostanza grigia, con la caratteristica forma ad ali di farfalla, contenente i corpi cellulari dei
neuroni. All'esterno troviamo la sostanza bianca (grigia, in figura, per la colorazione
istochimica), costituita da fibre nervose che formano fasci ascendenti o discendenti.
B
A
B
Fig. 2. A, cranio e colonna vertebrale (a sinistra) e 2 vertebre (a destra). B, due sezione
trasversali del midollo spinale, fatte a diversa altezza.
3
Fisiologia sensoriale nell'uomo
Le informazioni sull'ambiente esterno (anche il sangue, l'aria nei polmoni, le cavità gastrointestinali, il liquido prodotto dal rene possono essere considerati "ambiente esterno") sono
valutate dai recettori sensoriali. Ciascun recettore è elettivamente sensibile ad uno specifico
tipo di energia. Abbiamo recettori sensibili agli stimoli meccanici (meccanocettori), chimici
(chemocettori), luminosi (fotorecettori), ma i recettori possono essere stimolati anche da
energie di tipo differente, ma sempre più intense dello stimolo elettivo (Fig. 3).
Fig. 3. Procedendo da sinistra, troviamo due tipi di fotorecettori, i bastoncelli ed i
coni, una cellula ciliata, meccano-recettore del senso dell'udito e dell'equilibrio, i
recettori per l'odorato ed gusto, i cui corpi cellulari sono disposti in periferia, una
terminazione nervosa libera, attivabile, per esempio, dalla temperatura, ed una
terminazione nervosa accompagnata da strutture accessorie, il corpuscolo di Meissner,
che porta informazioni tattili
I segnali diventano percezioni quando raggiungono la corteccia. Ciascuna modalità
sensoriale arriva a specifiche zone o aree della corteccia primaria (Fig. 4). Nella zona
posteriore della corteccia troviamo l'area primaria visiva. Sul lobo temporale troviamo le aree
primarie gustativa ed uditiva. Posteriormente alla scissura di Rolando troviamo infine l'area
primaria somato-estesica, che riceve le afferenze relative al tatto. Le sensazioni che arrivano
alla corteccia primaria vengono poi analizzate dalle are associative connesse all'area primaria.
Specifiche aree sono deputate al riconoscimento della parola scritta od udita.
4
Area premotoria
Corteccia
motoria
Scissura di
Rolando
Corteccia somato-estesica
Area gustativa
Area associativa
somato-estesica
Area associativa visiva
Lobo frontale
Corteccia visiva
Area di Broca.
Area motoria
per la parola
Area uditiva
associativa
Area uditiva
primaria
Area di Wernicke.
Area uditiva per la
parola
Fig. 4. Aree sensitive primarie ed aree associative della corteccia (faccia laterale sinistra).
Sono anche indicate le aree motoria e premotoria, che controllano, rispettivamente, i
movimenti volontari ed i movimenti involontari associati ai movimenti volontari.
Le parti del corpo proiettano in modo ordinato sull'area somato-estesica, per cui, partendo
dalla zona latero-inferiore e procedendo verso l'alto e medialmente, troviamo le aree che
raccolgono informazioni dalla labbra e dalla lingua, dal viso, dalle mani, dalle braccia, dal
tronco, dalle gambe e dai piedi (Fig. 5, A). L'estensione delle aree non è proporzionale alle
dimensioni del corpo, ma alla finezza delle sensazioni tattili, cosicché l'area destinata a labbra,
lingua, viso e mani è molto maggiore di quella destinata a braccia, tronco, gambe e piedi. La
rappresentazione alterata delle proporzioni del corpo umano ha generato l'immaginario
homunculus di Penfield (Fig.5, B).
Come già accennato, le informazioni arrivano dapprima alle aree primarie. Qui vengono
elaborate e poi mandate alle aree associative, dove vengono elaborate ulteriormente. Per
esempio, informazioni visive grezze vengono inviate alla corteccia visiva primaria per una
prima elaborazione. L'output della corteccia visiva primaria viene poi analizzato ed elaborato
dalle aree di associazione visiva, che trasformano una serie di informazioni visive in un
concetto (un uomo, una casa, il mare). Una particolare area interpreta tutte le sensazioni
collegate al linguaggio; lesioni in tale area impediscono di riconoscere che un certo suono è una
parola, o impediscono di riconoscerne il significato. Alcuni tipi di lesione rimuovono la
connotazione emotiva delle sensazioni: per esempio, è possibile perdere la partecipazione
emotiva alla musica.
5
A
B
Fig. 5. A, localizzazione sulla corteccia somato-estesica dei neuroni che ricevono afferenze
dalle varie parti del corpo (da Casella-Taglietti). B, omunculus di Penfield.
6
Caratteristiche anatomo-funzionali del SNC: principi di base
Le operazioni del SNC sono funzione e del comportamento delle singole cellule nervose e di
come le cellule sono interconnesse.
Le singole cellule nervose hanno come specifica caratteristica strutturale quella di poter
possedere sia un lungo (anche 1 m) prolungamento, detto assone o fibra nervosa, sia numerosi
corti prolungamenti, detti dendriti. Hanno caratteristiche funzionali che ritroviamo in tutti i
vari tipi di cellule ma presentano però alcuni aspetti peculiari: la massiccia presenza nelle
fibre nervose di canali ionici al Na+(V), e la presenza di connessioni sinaptiche dove
l'esocitosi è pilotata da canali ionici al Ca2+(V).
I canali al Na+(V) sono i principali responsabili dell'insorgere e del rapido propagarsi del
potenziale d'azione, il meccanismo mediante cui le informazioni si trasmettono lungo le fibre
nervose; i canali al Ca2+(V) nelle terminazioni nervose sono attivati dal potenziale d'azione e
sono responsabili della esocitosi delle terminazioni presinaptiche.
Per quanto riguarda le connessioni tra cellule, nel SN dell'uomo troviamo 1011 neuroni,
ciascuno dei quali, in media, ha circa 105 connessioni sinaptiche, per un totale di 1016
connessioni. Il SNC dell'uomo differisce da quello del topo non tanto qualitativamente (le
singole cellule funzionano nello stesso modo) ma quantitativamente: per numero di neuroni e
per la complessità delle connessioni.
I canali ionici
I canali ionici sono presenti in tutte gli organismi, inclusi gli Archaea ed i batteri. Gran parte
delle classi funzionali dei canali ionici, compresi quelli che possono sembrare specifici del
sistema nervoso, hanno origine già dai procarioti. Il sequenziamento genico ci indica che molti
geni che noi associamo al movimento, alla comunicazione cellulare ed al SN nei metazoi si
sono evoluti dai procariotiin queste cellule, dove troviamo, per esempio, canali al Na+, al K+
ed al Ca2+ voltaggio-dipendenti. Esistono però differenze strutturali: per esempio, i canali al
Na+ ed al Ca2+ diventano tetramerici solo nei procarioti. I Coanoflagellati, molto affini ai
coanociti presenti in Porifera, mostrano già un'iniziale diversificazione genica dei canali al
Na+ ed al Ca2+ verso i canali dei metazoi.
7
Fig. 6. Rappresentazione schematica dei canali ionici dei metazoi. La topologia
transmembranaria è raffigurata da rettangoli che rappresentano domini transmembranari (il
dominio extracellulare è in alto). Le singole subunità peptidiche hanno differente colore.
color. (A) Kir (inward rectifier): la subunità ha 2 domini TM e nei metazoi è un tetramero,
un monomero nei procarioti.
Il recettore-canale per il glutammato ha un motivo Kir invertito con attaccato un TM (in
grigio), e costituisce un tetramero. (B) i canali cationi voltaggio-dipendenti hanno 4 domini
TM quali sensori di Vm (grigio) attaccati al motivo di Kir. Nel caso dei canali Na+(V) e
Ca2+(V) le 4 subunità sono unite a formare un monomero.
I canali ionici attivati dallo stimolo meccanico (stretch channel, non descritti in Fig. )
rappresentano probabilmente la più antica forma di trasduzione sensoriale. Lo stimolo agisce
stirando la membrana plasmatica che a sua volta deforma il canale, causandone l'apertura: il
canale è permeabile al Ca2+, un secondo messaggero chimicamente semplice ed ampiamente
disponibile, universalmente utilizzato per attivare la risposta meccanica di evitamento, la più
primitiva, ma altresì fondamentale, risposta degli organismi all'ambiente ( è "pericoloso"? Mi
allontano).
Possiamo portare quale esempio il protozoo Paramecium, nelle cui ciglia è presente un canale
attivato dallo stimolo meccanico permeabile al Ca2+: quando le ciglia toccano un ostacolo il
canale si apre e le ciglia cominciano a battere in modo da allontanarsi dall'ostacolo (vedi Film
in http://www.youtube.com/watch?v=fmwN_mD7TvY)
Il tipo di stimolo (interazione a contatto), il meccanismo di trasduzione (uno ione inorganico),
e la risposta motoria( una semplice retromarcia) sono tutti aspetti particolarmente "primitivi".
Nelle forme di vita attuali i canali ionici attivati dallo stimolo meccanico convertono stimoli
meccanici in segnali intracellulari che modulano il turgore dei batteri e delle piante, la
funzione tubulare e renale, la motilità del tubo digerente, la pressione sanguigna, e trasducono
gli stimoli sonori e, ovviamente, tattili.
Il segnale chimico rappresenta una seconda forma di stimolazione e costituisce la più ampia
classe di segnali in grado di influenzare un organismo. E' più evoluto dello stimolo meccanico,
sia perché lo stimolo ha un'energia di gran lunga inferiore sia perché l'organismo può
riconoscere differenti stimoli chimici ed uno stesso stimolo può evocare risposte le più
diverse.
8
Il protozoo Tetrahymena thermophila (Fig. 7) mostra il fenomeno della chemiotassi: nuota
verso molecole "attraenti" e cerca di evitare molecole "pericolose". Le proteine sono
chemoattrattive, mentre molecole secrete da predatori o prodotte dalla morte di organismi
sono repellenti.
Fig. 7. Tetrahymena thermophila
In questo caso è il contatto con singole molecole (incapaci di stimolare meccanicamente la
cellula) ad evocare una risposta motoria, che può qui essere duplice (mi avvicino o mi
allontano). La risposta è generata ancora una volta dal semplice ingresso di Ca2+,
probabilmente tramite canali aperti dalle molecole.
L'evoluzione ha generato numerosi meccanismi di trasduzione attivati da una immensa varietà
di molecole.
9
Le interazioni tra cellule e l'evoluzione del Sistema Nervoso
Meccanismi di interazione tra cellule sono presenti già nelle spugne, gli organismi più
semplici tra i metazoi (Fig. 8).
Fig. 8. Suddivisioni del regno dei Metazoi (Animalia). Nell'inserto, una colonia di
coanoflagellati.
Le spugne (Fig. 9) non solo sono dotate di canali ionici con strutture simili a quelle di tutti i
metazoi ma sembrano presentare, contrariamente a quanto generalmente ritenuto, sia un
abbozzo di coordinata interazione tra cellule sia un abbozzo di tessuto epiteliale.
Di fatto, le spugne sono sì costituite da alcuni tipi differenziati di cellule organizzate intorno
ad un sistema di canali, ma erano ritenute prive di epitelio e di un'attività multicellulare
coordinata. Recentemente, tuttavia, è stato visto che i coanociti delle spugne sono connesse da
tight junction. Inoltre stimoli esterni, come il contatto con piccoli crostacei o con detriti attiva
un'onda di contrazione che si propaga a tutto l'organismo. Questo movimento favorisce il
movimento dell'acqua e l'espulsione di detriti dai canali. La velocità di contrazione è molto
bassa (circa 0.5 mm/s) e sembra essere innescata da l'apertura di stretch channel che
consentono l'ingresso di Ca2+, inescando la contrazione della cellula. L'onda si propaga o
mediante una catena di stimolazione meccanica (una cellula tira l'altra) o mediante la
secrezione di mediatori chimici, quali l'ATP, che attivano la contrazione nella cellula vicina.
Questo meccanismo sembra essere l'antenato dei movimenti peristaltici presenti nel sistema
circolatorio ed intestinale.
10
Fig. 9. Struttura delle spugne. Giallo: pinacociti; rosso: coanociti; grigio: mesoilo o
mesoglia. La freccia indica la direzione del flusso di acqua. Nel mesoilo sono presenti altri
tipi di cellule.
Cnidaria, sebbene non abbiano un sistema nervoso centrale, presentano una rete di cellule
nervose connesse tra di loro, che consentono un'attività coordinata mediante una rapida (anche
di soli pochi ms) comunicazione tra cellule (Fig. 10). E' questo sicuramente un fondamentale
avanzamento dei processi evolutivi.
Rete
nervosa
Fig. 10. Rappresentazione schematica della rete nervosa di Hydra
La rete nervosa forma altresì alcuni plessi nervosi (addensamenti di fibre nervose) forse in
contatto diretto mediante ponti citoplasmatici. I neuroni di Cnidaria hanno spesso carattere
misto (sensitivi, motori, interneuroni e neurosecretori), ma presentano caratteristiche proprie
del sistema nervoso: potenziale d'azione, sinapsi e trasmissione chimica . Il sistema nervoso di
Cnidaria è fortemente peptidergico: i peptidi isolati, presenti anche in strutture presinaptiche,
possono inibire o attivare le rispettive fibre muscolari. Alcune specie posseggono porzioni di
sistema nervoso specificamente dedicate al controllo della nutrizione e del movimento ed
alcune meduse posseggono cellule sensibili alla luce ed alla gravità. Cnidaria possiedono
11
dunque i meccanismi di base del funzionamento del SN ed una, seppur primitiva,
organizzazione anatomo-strutturale.
Con il procedere del processo evolutivo, i meccanismi di base diventeranno più numerosi, ma
è soprattutto la complessità strutturale ad aumentare enormemente, portando ad un aumento
del numero dei neuroni e delle connessioni.
A partire dai Platelminti inizia la centralizzazione del SN (Fig. 11). Questa ha un duplice
vantaggio: rende la comunicazione tra le cellule centralizzate più rapida e meglio protegge la
struttura nervosa centralizzata. I rapporti con la periferia vengono velocizzati utilizzando
assoni giganti (come nei calamari) o progettando fibre dotate di mielina e di nodi di Ranvier.
Ganglio anteriore,
o cervello
Nervo
trasversale
Cordone nervoso laterale
Fig. 11. Sistema nervoso di planaria (Platelminti)
Gli anellidi hanno un cervello bilobato, 2 cordoni dorsali con gangli segmentali e neuroni
motori e sensitivi distinti. Sono spesso presenti assoni giganti, che assicurano una veloce
propagazione del PdA.
Gli artropodi hanno un SN organizzato in modo analogo a quello degli anellidi, ma i gangli
sono di maggiori dimensioni e gli organi di senso e l'attività motoria sono molto più sviluppati
(Fig. 12, A). Notevolmente complesso è il comportamento sociale.
Nei cefalopodi i gangli hanno dato origine ad un complesso sistema nervoso centrale e
periferico, con organi di senso molto sviluppati e capacità comportamentali sofisticate.
Infine nei vertebrati troviamo un SNC costituito da un cervello e da un midollo spinale, ed un
SNP costituito da gangli e nervi (Fig. 12, B).
Gangli
segmentali
Cervello
Cervello
Midollo
spinale
A
Gangli
sensoriali
B
Fig. 12. Sistema nervoso di una cavalletta (A) e di una salamandra (B).
12