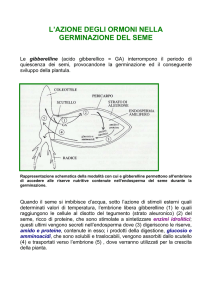
GIBERELLINE
Negli anni ’50 venne caratterizzato il secondo
gruppo di ormoni le GIBBERELLINE
Insieme di composti definiti in base alla loro struttura
chimica e non rispetto alla loro attività biologica (se ne
conoscono più di 136)
BAKANAE
(piantina sciocca)
Malattia del riso diffusa in
Asia causata dal fungo
Gibberella fujikuroi
(Fusarium moniliforme)
faceva crescere in altezza le
piante di riso e ne impediva la
produzione di semi
1930 (Giappone) Isolamento principio attivo
1950 (USA, GB) Struttura acido gibberellico
1950 (Giappone) Struttura GA1, GA2, GA3
1958 Identificazione e purificazione nelle piante (GA1)
ATTUALMENTE SONO NOTE 136 GIBBERELLINE
(12 presenti solo in G. fujikuroi, più di 100 presenti solo nelle
piante, 13 ubiquitarie)
NOMENCLATURA: GAx, a seconda dell’ordine
cronologico della scoperta
PRIMI STUDI
SULL’ATTIVITA’
DELLE GIBBERELLINE
effetto GA sulla crescita dello
stelo floreale del cavolo
effetto GA1 su mais nano
effetto GA sulla fioritura
di piante di carota
effetto GA3 sull’allungamento
del fusto di piante di pisello
STRUTTURA BASE
DELLE GIBBERELLINE
scheletro ent-gibberellanico
a 20 atomi di carbonio
GA12
(C20)
Le gibberelline
contengono 20 o 19 atomi di
carbonio
(perso durante il
metabolismo)
GA9
(C19)
DIFFERENZE NEL NUMERO E NELLA POSIZIONE DI -OH
- idrossilazioni in C3 e in C13 (generalmente sono le forme attive)
l’idrossilazione nella configurazione β del
carbonio 2 abolisce l’attività biologica
Nonostante il gran numero di gibberelline
presenti nelle piante, le analisi genetiche
hanno dimostrato che solo poche hanno
un’azione
ormonale biologicamente attiva.
Tutte le altre agiscono da precursori o
rappresentano forme disattivate.
BIOSINTESI
Le gibberelline costituiscono una grande
famiglia di acidi diterpenici sintetizzati da una
diramazione della via dei terpenoidi
vengono sintetizzate a partire dall’isopentenil
pirofosfato
Esistono due vie:
attraverso l’acido mevalonico (citosol)
attraverso la gliceraldeide 3-fosfato e il
piruvato (plastidi)
Probabilmente viene usata la seconda via
REAZIONI DI CICLIZZAZIONE (plastidi)
Il geranilgeranil pirofosfato è convertito in ent-kaurene
Enzimi coinvolti: CPS, copalil difosfato sintasi e KS,
ent-kaurene sintasi
SERIE DI OSSIDAZIONI (reticolo endoplasmatico)
-un gruppo metilico viene ossidato ad acido carbossilico
-l’anello B si contrae (da 6 a 5 atomi di C)
-enzimi coinvolti= monossigenasi che utilizzano il citocromo P450
Prima gibberellina che si forma in tutte le piante
FORMAZIONE DI TUTTE LE GIBBERELLINE A PARTIRE
DALLA GA12-ALDEIDE (citosol)
Enzimi=
diossigenasi
-ossidazione C7 (da GA12 aldeide a GA12)
-idrossilazioni C13 e C3
-ossidazioni C20
-perdita atomo di C
-idrossilazione C2 (inattivazione)
INIBITORI SINTESI
GIBBERELLINE
Interferiscono con gli enzimi che utilizzano
come substrato α-chetoglutarato
La maggior parte delle gibberelline sono dei
precursori di quelle biologicamente attive
Nella maggior parte delle piante
l’unica gibberellina attiva è la GA1
Altre GAs attive:
GA3
GA4 e GA7
(Arabidopsis, cetriolo)
GA9
EFFETTI DELLE GIBBERELLINE SULLA CRESCITA E SVILUPPO
q stimolano la crescita del fusto in piante nane
e a rosetta
q regolano la transizione dalla fase giovanile ad adulta
q Influiscono sulla formazione dei fiori e sulla
determinazione del sesso
q Promuovono la fruttificazione
q Promuovono la germinazione dei semi
(interruzione della dormienza)
APPLICAZIONI COMMERCIALI
q Produzione di frutti
(aumento lunghezza del picciolo in uva priva di semi;
forma mele Delicious)
q Produzione di malto da orzo
q Aumento rese canna da zucchero
q Allevamento vegetale
(conifere)
+GA
Le gibberelline sono dei regolatori di crescita naturali?
Le piante alte hanno più GAs delle piante nane?
MUTANTI DI PISELLO le
Purificazione GA (HPLC, GC-MS)
Le piante Le hanno un contenuto
maggiore di GA1 rispetto alle
piante le
le
Le
Le piante le rispondono a GA1, ma non a GA20
+GA1
+GA20
Il gene Le conferisce la capacità di
convertire la GA20 in GA1
Le
[13C, 3H] GA20 → *GA1, *GA8, *GA29
le
[13C, 3H] GA20 → *GA29
Esistono dei mutanti di pisello na
(nana) molto più bassi dei mutanti le
Sono incapaci di convertire l’entkaurene in GA12-aldeide
I mutanti Slender(la cry) sono
alti anche quando non hanno GAs
(na la cry)
Questi mutanti contengono elevate
concentrazioni di auxina, e forse un
recettore di “attivazione” dell’acido
giberellico oppure attraverso un processo
successivo che regola la crescita
tessuti di sintesi delle GAs
germogli di pisello
enzimi biosintetici localizzati in
gemme di giovani foglie
internodi superiori
Arabidopsis
enzimi biosintetici localizzati in
gemma apicale giovani foglie
promotore GA20ox::luciferasi
TRASPORTO
floema
vengono trasportati anche i vari metaboliti
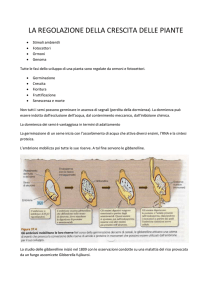
REGOLAZIONE DELLA SINTESI DELLE GIBBERELLINE
CONTROLLO FEEEDBACK
Alte concentrazioni di GAs inibiscono
la produzione di ulteriori molecole di GAs
le GAs inducono la
produzione di un
repressore trascrizionale!
q Fattori ambientali come luce, fotoperiodo e temperatura,
che influenzano lo sviluppo delle piante, alterano
i livelli di gibberelline
q L’effetto è dovuto alla alterazione della trascrizione
di geni codificanti per enzimi della via di biosintesi delle
gibberelline
luce
: regolazione della sintesi di GA1
SEMI FOTOBLASTICI
luce
(germinano al buio per aggiunta di GA1)
↑GA3ox
↑GA1
germinazione
DEEZIOLATURA DEI
GERMOGLI
La crescita è rallentata
meno GA1?
alla luce i livelli di GAs aumentano
si attiva la
GA2ox
FOTOPERIODO
I livelli di GAs ossidrilate in 13
sono molto bassi
Giorni sempre più lunghi
All’aumentare della lunghezza del giorno iniziano ad aumentare
le concentrazioni delle GAs della via di ossidrilazione in 13
TEMPERATURA
Alcune piante richiedono basse temperature per germinare
(stratificazione) e per fiorire (vernalizzazione)
Le gibberelline possono sostituire il trattamento a basse
temperature per produrre questi effetti
fioritura Thalapsi arvense
NO TRATTAMENTO A
BASSE TEMPERATURE
TRATTAMENTO A
BASSE TEMPERATURE
alti livelli acido ent-kaurenoico
alti livelli GAs
(principalmente GA9)
PROMOZIONE CRESCITA DEL FUSTO
allungamento internodo superiore riso
aumento estensibilità parete
cellulare
No acidificazione apoplasto
(auxina)
VARIE IPOTESI
Alterazione distribuzione del calcio
(<[Ca2+] parete, >assunzione Ca2+ cellula)
Attivazione xiloglucano endotransglicosidasi (XET)
STIMOLO DIVISIONE CELLULARE
Incremento mitosi nelle regioni apicali delle piante a rosetta
Nei meristemi le GAs stimolano
la transizione tra la fase G1 e la
fase S del ciclo cellulare
Nel riso le GA aumentano
l’espressione di una CDK (proteina
chinasi ciclina-dipendente) e di una
protein chinasi in grado di attivarla
ALTRI EFFETTI FISIOLOGICI DELLE
GIBBERELLINE
Regolazione della transizione da stato
giovanile a fase adulta
Induzione della germinazione dei semi
Diversi meccanismi
Attivazione della
crescita vegetativa
dell’embrione
Interruzione della dormienza
dei semi (insensibilità alla
temperatura)
Mobilizzazione delle
riserve dell’endosperma
semi dei cereali
embrione
embrione
scutello
endosperma
endosperma aleurone
amilaceo
Ricco di amido
Corpi proteici
I due enzimi responsabili della degradazione dell’amido
sono: α-amilasi e β-amilasi .
MECCANISMO DI AZIONE
Legame al recettore
Attivazione di una o più vie di
trasduzione
Trascrizione dei geni primari
della risposta
Trascrizione dei geni
secondari
RISPOSTA
GA= attive fino
a 10-14 M
In un mutante nano di riso (GA-insensitive dwarf 1) è
stato identificato il recettore per le GA denominato GID1
α-amilasi in protoplasti aleurone
Presenza del recettore delle GA nell’endosperma
amilaceo dei semi
Trasduzione del segnale dal recettore alla PROTEINE G
peptide Mas7 (stimola scambio
GTP/GDP) attiva risposta
In Arabidopsis sono stati identificati 5 geni DELLA:
GA- INSENSITIVE (GAI), REPPRESSOR of ga1-3 (RGA), RGL1,
RGA 2, RGL3
Sottoclasse della famiglia GRAS di regolatori trascrizionali
Meccanismo molecolare GA-GID1-DELLA
Degradazione
Proteasoma 26S