Il trasporto del glucosio
• Il glucosio non può diffondere direttamente
nelle cellule
• Esso entra nelle cellule mediante due
meccanismi di trasporto:
a) diffusione facilitata Na+ indipendente
b) co-trasporto Na+ -monosaccaride
Diffusione facilitata Na+ indipendente
• Avviene secondo il gradiente di concentrazione, per
cui non richiede energia!
• E’ un processo mediato da una famiglia di proteine
comprendente almeno 14 trasportatori (GLUT1…..GLUT-14)
• L’espressione è tessuto specifica (Glut-1 G. Rossi ed
encefalo, Glut-2 fegato, rene...)
• Questi trasportatori si trovano nella membrana
cellulare
• Il glucosio extracellulare si lega al trasportatore,
esso modifica la propria conformazione e così
trasporta il glucosio attraverso la membrana
Co-trasporto Na+monosaccaride
• Richiede energia !
• Trasporta il glucosio contro il suo gradiente di
concentrazione
• E’ mediato da un trasportatore
• Il trasporto del glucosio è accoppiato al
gradiente di concentrazione del Na+
(contemporaneamente trasportato dentro la
cellula)
• Questo trasporto avviene nelle cellule epiteliali
dell’intestino, dei tubuli renali e del plesso
corioideo
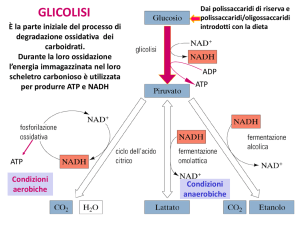
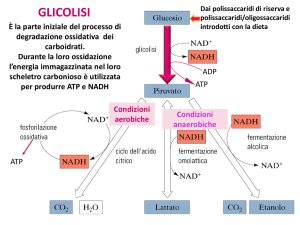
Le reazioni della glicolisi (2)
• (1) Fosforilazione del glucosio
• Intrappola lo zucchero nel citosol (gli
zuccheri fosforilati non attraversano le
membrane cellulari)
• è irreversibile
• è catalizzata dall’enzima esochinasi (uno dei 3
enzimi regolatori della glicolisi, insieme a
fosfofruttochinasi e piruvato chinasi)
• Nel fegato e nel pancreas è catalizzata dalla
glucochinasi
L’ esochinasi
• L’esochinasi catalizza la fosforilazione del glucosio
nella maggior parte dei tessuti
• Ha un’ampia specificità per il substrato (fosforila
anche altri esosi)
• è inibita dal prodotto della reazione (glucosio 6fosfato)
• ha una Km bassa per il glucosio (e quindi un’affinità
alta)
• ha una Vmax bassa, per cui non possono essere
fosforilati più zuccheri di quanti la cellula non ne
possa utilizzare.
La glucochinasi
• E’ il principale enzima responsabile della fosforilazione
del glucosio nel fegato e nelle cellule delle isole
pancreatiche.
• Nel fegato facilita la fosforilazione del glucosio
durante l’iperglicemia.
• Differisce dalla esochinasi perché:
1) ha una Km >> esochinasi (cioè richiede una
[ glucosio] più elevata per raggiungere 1/2 Vmax) perciò
funziona soltanto quando la [glucosio] negli epatociti è
alta (p.es. dopo un pasto ricco di carboidrati)
2) ha una Vmax elevata (permette al fegato di eliminare
con efficienza dal sangue portale il glucosio, anche dopo
un pasto ricco di carboidrati, minimizzando
l’iperglicemia durante il periodo dell’assorbimento)
REGOLAZIONE DELLA GLUCOCHINASI
INSULINA
Negli epatociti, l’attività della glucochinasi.
• FRUTTOSIO 6-FOSFATO e GLUCOSIO
Nel nucleo c’è una proteina regolatrice della
glucochinasi:
1) in presenza di fruttosio 6-fosfato (che si trova in
equilibrio con il glucosio 6-fosfato), l’enzima si
trasferisce nel nucleo, si lega alla proteina regolatrice e
diventa inattivo.
2) un di glucosio attiva l’enzima poiché provoca il
distacco della glucochinasi dalla proteina regolatrice.
L’enzima migra nel citosol e fosforila il glucosio a
glucosio 6-fosfato
(2) L’isomerizzazione del glucosio 6-fosfato in
fruttosio 6-fosfato
• E’ catalizzata dalla fosfoglucosio-isomerasi
• E’ reversibile
(3) Fosforilazione del fruttosio 6-fosfato a
fruttosio 1,6 bisfosfato
• è irreversibile
• è catalizzata dalla fosfofruttochinasi-1 (PFK-1)
• E’ il punto di controllo più importante della glicolisi ed è
la tappa che limita la velocità della via
• La PFK-1 è inibita da: ATP, citrato
• La PFK-1 è attivata da: AMP, fruttosio 2,6bisfosfato.
Regolazione della
fosfofruttochinasi-1 (PFK-1)
• La PFK-1 è INIBITA in modo allosterico da:
[ATP] (indicano abbondanza di composti
ad alta energia)
[citrato]
• La PFK-1 è ATTIVATA in modo allosterico
da:
[AMP] (indicano un impoverimento della
riserva energetica della cellula)
• fruttosio 2,6-bisfosfato
• Il più potente ATTIVATORE della PFK-1
è il fruttosio 2,6-bisfosfato
• esso è sintetizzato in tutti i tessuti
• la sua sintesi è (+) dall’insulina e (-) dal
glucagone
• funge da segnale intracellulare,
indicando un’abbondanza di glucosio
• La sua azione contemporanea sulla
glicolisi [(+) fosfofruttochinasi-1] e sulla
gluconeogenesi [(-) fruttosio 1,6
bisfosfatasi] fa sì che le due vie non
siano attive nello stesso momento,
evitando un ciclo futile.
• Durante lo stato alimentato nel fegato
[fruttosio 2,6 bisfosfato] con della
glicolisi.
• Durante il digiuno nel fegato si ha una
[fruttosio 2,6 bisfosfato ] con
conseguente della gluconeogenesi.
Regolazione della piruvato chinasi
(a livello epatico)
• Nel fegato, la piruvato chinasi è attivata da un
[fruttosio 1,6-bisfosfato] (il prodotto della
reazione della fosfofruttochinasi-1)
• Questa regolazione collega l’attività delle due
chinasi:
attività fosfofruttochinasi (PFK-1)
[fruttosio 1,6-bisfosfato]
(+) piruvato chinasi
• La fosforilazione della piruvato chinasi (ad opera di
una proteina chinasi cAMP dipendente) determina la
sua inattivazione
1) Quando la [glucosio] è bassa:
[glucagone] [cAMP] intracellulare
fosforilazione della piruvato chinasi (-) piruvato
chinasi
2) Il fosfoenolpiruvato (PEP) non può continuare la via
glicolitica e sarà utilizzato nella via gluconeogenica
• La defosforilazione della piruvato chinasi (ad opera
di una fosfatasi) riattiva l’enzima
REGOLAZIONE DELLA GLICOLISI
• La regolazione della glicolisi è affidata a tre passaggi di
importanza strategica:
1) fosforilazione del glucosio (esochinasi e glucochinasi);
2) la formazione di fruttosio 1,6-bisfosfato (PFK-1);
3) la formazione di piruvato (piruvato chinasi).
• Nei tessuti, la glicolisi è un processo molto inibito cioè
funziona ad una velocità molto bassa rispetto alla Vmax .
• In tal modo le cellule possono modulare velocemente
questo processo, senza ricorrere alla produzione di
nuove proteine ed enzimi.
• La regolazione della glicolisi è affidata ad enzimi
regolatori che, in condizioni basali, sono molto inibiti.
• Il controllo di questi enzimi avviene ad opera di
effettori allosterici (piccole molecole che si legano ad
un sito diverso dal sito attivo) o per mezzo di
modificazioni covalenti (fosforilazione di una proteina).
REGOLAZIONE ORMONALE
DELLA GLICOLISI
• La sintesi dei 3 enzimi chiave della glicolisi
(glucochinasi, fosfofruttochinasi e piruvato
chinasi) è:
dall’ insulina
dal glucagone
• Un pasto ricco di carboidrati o la
somministrazione di insulina la sintesi dei
3 enzimi (è favorita la conversione del
glucosio in piruvato)
• Nel digiuno e nel diabete, la [glucagone] è
e la sintesi dei 3 enzimi è
Champe et al., Le basi della biochimica, Ed. Zanichelli
Destini alternativi del piruvato
1) Decarbossilazione ossidativa
• via importante nei tessuti con elevata capacità
ossidativa (cuore)
• catalizzata dalla piruvato deidrogenasi
• converte irreversibilmente il piruvato in acetil CoA
(utilizzato nel Ciclo di Krebs o per la sintesi degli
acidi grassi)
2) Carbossilazione ad ossalacetato
• catalizzata dalla piruvato carbossilasi
• rigenera gli intermedi del ciclo di Krebs e fornisce
substrati alla gluconeogenesi
3) Riduzione del piruvato ad etanolo (F. alcolica)
• avviene nei lieviti e in alcuni batteri