Adrian T. Sumner/Science Photo Library/Photo Researchers, Inc.
Cromosomi appaiati durante la metafase della prima divisione meiotica,
il meccanismo che produce i gameti, quali uova e spermatozoi (SEM colorata).
11
Piano di studio
11.1
I meccanismi della meiosi
La meiosi si basa sulle interazioni e sulla distribuzione
delle coppie di cromosomi omologhi.
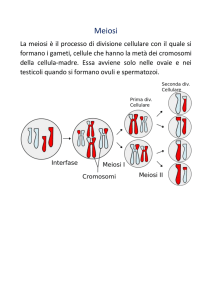
Il ciclo cellulare meiotico produce quattro cellule figlie geneticamente diverse, ciascuna con la metà dei cromosomi della cellula progenitrice.
11.2
I meccanismi che producono variabilità genetica
La variabilità prodotta dalla ricombinazione è conseguente all’appaiamento cromosomico e agli scambi fisici
che avvengono tra cromatidi omologhi.
La segregazione casuale dei cromosomi materni e paterni
costituisce la seconda fonte principale di variabilità genetica durante la meiosi.
L’unione casuale dei gameti maschile e femminile alla fecondazione produce ulteriore variabilità genetica.
11.3
Momento e sede della meiosi nel ciclo vitale degli organismi
Negli animali predomina la fase diploide, mentre la fase
aploide è ridotta e la formazione dei gameti segue direttamente la meiosi.
Nella maggior parte delle piante e dei funghi si ha alternanza di generazioni, entrambe pluricellulari, in fase aploide e diploide.
In alcuni funghi e in altri organismi predomina la fase
aploide, mentre la fase diploide è ridotta ad una sola cellula.
Meiosi: le basi cellulari
della riproduzione
sessuale
PERCHÉ È IMPORTANTE
Una coppia dimostra interesse reciproco. Prima egli la accarezza con un
braccio, poi con un secondo, poi con un altro e un altro ancora. Lei ricambia e questa interazione va avanti per ore; un abbraccio qui, una stretta là.
Al momento giusto il maschio raggiunge il suo mantello, prende un pacchetto di spermatozoi e lo introduce sotto il mantello della femmina. Da
ciascuno degli spermatozoi che riesce a portare a termine la sua funzione, si
originerà uno zigote che darà origine ad un nuovo polpo.
Per il polpo, il sesso è un evento occasionale che viene preceduto da
un rituale di corteggiamento effettuato tramite un intreccio di tentacoli.
Per un altro animale marino, la patella, è un’attività di gruppo che dura
tutta la vita. Le patelle sono parenti stretti delle chiocciole. Al pari di molti
altri animali, la patella attraversa un periodo di immaturità allo stato libero prima di trasformarsi in adulto. Quando arriva il momento di diventare adulta, si fissa ad una roccia o a una qualsiasi superficie fissa. Se si
fissa isolata ad un substrato diventa femmina, se invece si fissa su una
femmina diventa maschio. Se un’altra patella si fissa sul maschio, diviene
essa stessa maschio. Le patelle adulte vivono solitamente così impilate
l’una sull’altra e quella sul fondo è sempre una femmina. Tutti i maschi
producono continuamente spermatozoi che fecondano le uova deposte
221
dalla femmina. Se la femmina muore, il maschio che rimane in fondo alla pila si trasforma in femmina, e così la
riproduzione continua.
Questi polpi e patelle hanno una riproduzione sessuale, cioè generano figli per mezzo della fusione di gameti maschili e femminili, uova e spermatozoi negli animali. La riproduzione sessuale dipende quindi dalla
meiosi, un processo di divisione cellulare specializzato
nella produzione di gameti. La meiosi porta alla riduzione
del numero di cromosomi, cioè produce gameti che hanno la metà dei cromosomi presenti nelle cellule somatiche
di una specie. L’etimologia del termine meiosi (meioun =
diminuire) si riferisce proprio a tale riduzione. Alla fecondazione i nuclei dell’uovo e dello spermatozoo si fondono
e formano una cellula chiamata zigote, nella quale viene
ripristinato il numero diploide di cromosomi tipico della
specie. Senza il dimezzamento del numero di cromosomi
che avviene con la meiosi, la fecondazione porterebbe al
raddoppiamento del numero di cromosomi ad ogni successiva generazione.
La meiosi e la fecondazione hanno anche la funzione di
mescolare le informazioni genetiche e produrre nuove
combinazioni in modo che nessuno dei figli di una coppia
sia geneticamente identico. Al contrario, la riproduzione
asessuale produce figli identici perché essi vengono prodotti attraverso divisioni mitotiche (la riproduzione ases-
Adulti diploidi (2n)
46 cromosomi
Zigote diploide (2n)
46 cromosomi
FECONDAZIONE
Fase diploide (2n) del ciclo vitale
MEIOSI
Fase aploide (n) del ciclo vitale
Uovo aploide (n)
23 cromosomi
Spermatozoo aploide (n)
23 cromosomi
Figura 11.1
Il ciclo della meiosi e della fecondazione. La meiosi riduce il numero dei cromosomi dall’assetto
diploide (due membri per ogni cromosoma) a quello aploide (un membro per ogni cromosoma).
La fecondazione ripristina il corredo diploide.
222
UNITÀ DUE
GENETICA
suale è illustrata nel Capitolo 10). La riproduzione sessuale
produce pertanto la variabilità che è alla base della maggior
parte delle differenze ereditarie tra gli organismi che si riproducono per via sessuale. Questa variabilità è alla base dei
processi evolutivi.
Il dimezzamento del numero di cromosomi della specie e il mescolamento delle informazioni genetiche in
nuove combinazioni – entrambi prodotti dalla meiosi – e
il ripristino del corredo cromosomico diploide alla fecondazione costituiscono gli aspetti biologici fondamentali
della riproduzione sessuale. Lo scambio di tentacoli nei
polpi e la comunità sessuale nelle patelle, così come il corteggiamento e i rituali di accoppiamento nell’uomo, non
sono altro che variazioni nelle modalità usate per raggiungere lo scopo fondamentale della procreazione, la fecondazione.
11.1 I meccanismi della meiosi
La meiosi avviene solo negli eucarioti che si riproducono
sessualmente e solo negli organismi che sono almeno diploidi, cioè organismi che posseggono almeno due copie di
ciascun cromosoma.
La meiosi si basa sulle interazioni
e sulla distribuzione delle coppie
di cromosomi omologhi
Per poter seguire bene le tappe della meiosi, bisogna avere
le idee chiare su come sono costituite le coppie di cromosomi negli organismi diploidi. Come già illustrato nella
Sezione 10.1, i due membri di uno stesso cromosoma in
una cellula diploide formano una coppia di omologhi, ovvero hanno gli stessi geni (sono cioè costituiti dallo stesso
DNA), che sono distribuiti con lo stesso ordine sul cromosoma. Un cromosoma di ciascun paio, il cromosoma
paterno, viene ereditato dal padre, l’altro, il cromosoma
materno, dalla madre.
Anche se i due cromosomi omologhi di una coppia
contengono gli stessi geni distribuiti con lo stesso ordine,
forme diverse di ogni gene, dette alleli, possono essere presenti nell’uno o nell’altro cromosoma della coppia. Gli alleli dello stesso gene hanno sequenze diverse di DNA e codificano per forme diverse della stessa proteina, che possono avere differente struttura o composizione chimica, o
sia l’una sia l’altra.
Per esempio, l’uomo ha normalmente 46 cromosomi
nelle sue cellule, cioè 23 coppie di omologhi (vedi Figura
10.7). Tuttavia, ciascun individuo (eccetto i gemelli identici) possiede una combinazione unica di alleli nei due
cromosomi omologhi che formano una coppia. Il diverso
complesso di alleli che si origina a seguito dei meccanismi di meiosi e fecondazione determina in ogni individuo di sesso maschile o femminile la presenza di una
combinazione unica di caratteri ereditari, quali ad esempio l’altezza, il colore degli occhi, la suscettibilità a certe
malattie e persino vari aspetti della personalità e dell’intelligenza.
La meiosi separa gli omologhi di ogni coppia, riducendo così il corredo cromosomico diploide (2n) ad
aploide (n) (Figura 11.1). Ogni gamete prodotto tramite la
meiosi riceve solo un membro di ogni coppia di omologhi.
Per esempio, un uovo o uno spermatozoo umano conten-
gono 23 cromosomi, uno per ognuna delle coppie. Quando
l’uovo e lo spermatozoo si uniscono nella riproduzione sessuale e producono lo zigote – la prima cellula del nuovo organismo – si ricostituisce il corredo diploide di 46 cromosomi (23 coppie). Il processo di replicazione del DNA e la
divisione cellulare per mitosi assicurano che il numero diploide di cromosomi venga mantenuto nelle cellule somatiche che si formano a partire dallo zigote.
Il ciclo cellulare meiotico produce quattro
cellule figlie geneticamente diverse,
ciascuna con la metà dei cromosomi
della cellula progenitrice
(meiosi II), i cromatidi fratelli si separano a loro volta e si distribuiscono (segregano) in cellule diverse. Alla fine dell’intero processo meiotico si producono quattro cellule, ciascuna con un corredo cromosomico aploide.
Per convenienza i biologi suddividono la divisione
meiotica negli stessi stadi della mitosi: profase, prometafase,
metafase, anafase e telofase. Gli stadi, a seconda della divisione alla quale appartengono, vengono identificati con la
simbologia I o II (ad esempio, profase I o profase II). Le due
divisioni meiotiche sono intervallate da una breve interfase
detta intercinesi, durante la quale peraltro è bene ricordare
che non si ha replicazione del DNA.
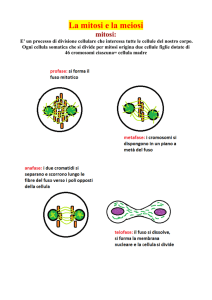
Profase I. All’inizio della profase I, i cromosomi raddop-
La meiosi è un processo di divisione cellulare che avviene in due tappe (dette meiosi I e meiosi II) negli organismi che si riproducono per via sessuale. Durante la
meiosi, i cromosomi raddoppiati della cellula progenitrice vengono distribuiti in quattro cellule figlie, ciascuna delle quali avrà pertanto la metà dei cromosomi
della cellula progenitrice. Nella mitosi, invece, ad ogni
raddoppiamento dei cromosomi segue una divisione
cellulare, perciò il numero dei cromosomi rimane costante da una generazione all’altra.
Il ciclo cellulare meiotico inizia con una interfase
premeiotica quando il DNA si replica di pari passo con
il raddoppiamento delle proteine cromosomiche.
(Questa parte della interfase comprende gli stadi G1, S e
G2, al pari della interfase premitotica). Come nella interfase premitotica, il risultato di questa duplicazione è
la formazione di due cromatidi fratelli identici per ciascun cromosoma (Figura 11.2). Dopo questa fase le cellule iniziano le due divisioni meiotiche, meiosi I e
meiosi II. I cromosomi omologhi della stessa coppia si
appaiano e possono scambiarsi segmenti cromosomici
nel corso della prima divisione; questo processo è detto
ricombinazione. Dopo la ricombinazione i cromosomi
omologhi si separano e la cellula prosegue la meiosi I, al
termine della quale si ottengono due cellule, ciascuna
con un corredo cromosomico aploide nel quale ogni
cromosoma è costituito da due cromatidi (è cioè bicromatidico). Nel corso della seconda divisione meiotica
piati, costituiti ciascuno da due cromatidi fratelli, incominciano a condensarsi ed avvolgersi in strutture filamentose
bastoncellari all’interno del nucleo (Figura 11.3, stadio 1).
Coppia di cromosomi
omologhi
Cellula diploide con i due
cromosomi omologhi
di una coppia.
Duplicazione dei cromosomi
durante la interfase premeiotica
Cromatidi fratelli
Il risultato della duplicazione
è che ciascun cromosoma è
costituito da due cromatidi fratelli.
Appaiamento dei cromosomi omologhi
durante la profase meiotica I
Mentre sono appaiati, i cromosomi omologhi
(o meglio i cromatidi omologhi) possono
scambiarsi segmenti e andare così incontro
al fenomeno della ricombinazione genetica.
Figura 11.2
Produzione di quattro nuclei aploidi
come conseguenza delle due divisioni
meiotiche. (Per semplicità è stata rappresentata una sola coppia di cromosomi omologhi con i due membri di colore diverso a seconda della provenienza materna o paterna).
Prima divisione
meiotica
La prima divisione meiotica consiste
nella separazione degli omologhi, che
passano ciascuno in una delle due
cellule figlie prodotte dalla divisione.
Tali cellule contengono un numero
aploide di cromosomi, anche se ciascuno
di essi è tuttora composto da due cromatidi.
Seconda divisione
meiotica
Alla seconda divisione
meiotica i cromatidi fratelli
si separano e migrano
ciascuno in una delle cellule
derivate da questa divisione.
CAPITOLO 11
MEIOSI: LE BASI CELLULARI DELLA RIPRODUZIONE SESSUALE
223
Prima divisione meiotica
Profase I
Membrama
plasmatica
Centrioli
duplicati
Cromosomi
omologhi
Involucro
nucleare
Tetrade
Due cromatidi
fratelli
Condensazione dei cromosomi
1 All’inizio della profase I i cromosomi
incominciano a condensarsi e a formare
delle strutture bastoncellari. Ciascun
cromosoma è costituito da due
cromatidi fratelli a seguito della
precedente replicazione del DNA
che avviene nel corso della interfase
premeiotica. Sono rappresentate due
coppie di cromosomi omologhi, una
lunga e una corta.
Sinapsi
2 I cromosomi omologhi
si avvicinano e si appaiano.
Ricombinazione
3 Mentre sono appaiati, i cromatidi
appartenenti a cromosomi omologhi
possono scambiarsi segmenti e andare
incontro a ricombinazione genetica.
Il cerchio indica un sito di scambio.
(Vedi anche la freccia).
Prometafase I
4 Nella prometafase I l’involucro nucleare si
frammenta e il fuso va a collocarsi dove prima
era situato il nucleo. I microtubuli del fuso si
legano al cinetocore dei cromosomi con un
preciso e specifico processo: i microtubuli che
si dirigono verso un polo del fuso si attaccano
ad entrambi i cromatidi fratelli di un cromosoma
e li trasportano a quel polo, mentre i microtubuli
del fuso che si dirigono al polo opposto si
uniscono ad entrambi i cromatidi fratelli dell’altro
omologo e li trasportano uniti all’altro polo.
Seconda divisione meiotica
Figura 11.3
Le divisioni meiotiche. Il disegno rappresenta la sequenza degli eventi che
avvengono in una cellula germinale maschile (sono rappresentate due coppie di
cromosomi omologhi).
224
UNITÀ DUE
GENETICA
Profase II
Prometafase II
8 I cromosomi di ricondensano
e si forma un nuovo fuso.
9 L’involucro nucleare si frammenta
ancora, il fuso si colloca nell’area dove
prima era situato il nucleo e i microtubuli
provenienti da (o diretti verso) i poli
opposti si attaccano al cinetocore di
ciascun cromosoma.
Piastra
metafasica
Coppia
di omologhi
Intercinesi:
non si ha
alcuna
duplicazione
del DNA tra
la prima
e la seconda
divisione
Metafase I
5 I movimenti dei
microtubuli del fuso
determinano l’allineamento
delle tetradi sul piano
equatoriale – detto piastra
metafasica – collocato tra
i due poli del fuso.
Metafase II
10 I movimenti dei
microtubuli del fuso
determinano l’allineamento
dei cromosomi sulla piastra
metafasica.
Anafase I
Profase II
della
seconda
divisione
meiotica
Telofase I
6 I microtubuli del fuso sono
responsabili della separazione dei due
cromosomi omologhi di ciascuna
coppia e determinano la loro
migrazione verso i poli opposti
del fuso. A questo punto ogni polo
contiene un numero aploide di
cromosomi, ciascuno dei quali è
comunque composto da due cromatidi.
7 In questo stadio i cromosomi
non subiscono praticamente alcun
cambiamento, se non una limitata
decondensazione e distensione in
alcune specie. Il fuso della prima
divisione meiotica scompare e si
formano due nuovi fusi in vista
della seconda divisione.
Anafase II
Telofase II
11 I microtubuli del fuso
provocano la separazione
dei due cromatidi di ciascun
cromosoma e li trasportano
ai poli opposti del fuso.
12 I cromosomi iniziano a
decondensarsi (distendersi),
le fibre del fuso si
disassemblano e si forma un
nuovo involucro nucleare.
CAPITOLO 11
MEIOSI: LE BASI CELLULARI DELLA RIPRODUZIONE SESSUALE
225
SPUNTI DALLA RIVOLUZIONE MOLECOLARE
Regioni di fertilità nel cromosoma Y umano
Una parte del cromosoma Y umano contiene geni attivi e si appaia con la regione
omologa del cromosoma X durante la
meiosi. La parte rimanente del cromosoma Y, che non si appaia con il cromosoma X, è stata a lungo ritenuta inerte,
priva di geni funzionali. Ma gli scienziati
hanno individuato in questa regione la
presenza di vari geni. Uno di essi è il gene
SRY, che regola la formazione dei testicoli
negli embrioni maschili. Altri, scoperti recentemente, comprendono un gruppo
(cluster) di otto geni “housekeeping”,
cioè geni che codificano per proteine
fondamentali in processi essenziali, quale
la sintesi proteica, che avvengono in ogni
cellula. Era già noto che i geni “housekeeping” erano presenti sul cromosoma X e
su alcuni autosomi, ma è stato sorprendente trovarli in una regione del cromosoma Y che non ha omologia con alcuna
regione dell’X. Questa scoperta inattesa
ha portato due ricercatori del Massachusetts Institute of Technology, Bruce T.
Lahn e David C. Page, a chiedersi se altri
geni potessero essere presenti nella regione dell’Y che non si appaia con l’X. Un
indizio che vi potrebbero essere localiz-
zati altri geni è fornito dal fatto che delezioni in questa regione possono portare
nell’uomo a sterilità e alla comparsa di tumori testicolari, il che ha suggerito che il
cromosoma Y possa contenere sequenze
di DNA di fondamentale importanza per
la fertilità maschile.
Per individuare eventuali altri geni,
Lahn e Page hanno usato preliminarmente tecniche di analisi genetica per
eliminare le regioni del cromosoma Y
non codificanti, cioè prive di geni, così
come quelle che contenevano geni già
noti. In seguito all’uso di queste tecniche
sono stati evidenziati alcuni frammenti di
cromosoma contenenti geni al momento sconosciuti. L’analisi molecolare
condotta su questi frammenti ha dimostrato la presenza di dodici geni, tutti
nella regione dell’Y che non si appaia
con il cromosoma X. Lahn e Page hanno
usato un programma computerizzato
per confrontare le sequenze di aminoacidi codificate dai 12 geni con la banca
dati delle sequenze proteiche note. Il
confronto ha dimostrato che cinque di
questi geni sono “housekeeping” (uno,
per esempio, codifica per una proteina
Quindi i due cromosomi omologhi di ciascuna coppia si
uniscono allineandosi punto per punto a formare una
struttura simile ad una cerniera; questo processo è detto appaiamento o sinapsi (Figura 11.3, stadio 2). Quando sono totalmente appaiati per tutta la loro lunghezza, i cromosomi
di una coppia formano la cosiddetta tetrade, in quanto ciascun membro è costituito da due cromatidi, quattro in totale. Nella mitosi non avviene un simile appaiamento dei
cromosomi omologhi.
Mentre sono appaiati, i cromatidi appartenenti a cromosomi omologhi si scambiano fisicamente dei segmenti
(Figura 11.3, stadio 3). Questo scambio fisico, detto ricombinazione, costituisce il meccanismo mediante il quale gli alleli localizzati sui cromosomi omologhi assumono nuove
combinazioni, contribuendo ad incrementare la variabilità
nella riproduzione sessuale (vedi più avanti in questo stesso
capitolo i dettagli sul meccanismo della ricombinazione genica).
Al termine della profase I nel citoplasma si forma un
fuso, con le stesse modalità già illustrate nella Sezione
10.3.
Prometafase I. Nella prometafase I, l’involucro nucleare si
frammenta e il fuso si porta nell’area dove si trovava il nucleo (Figura 11.3, stadio 4). I due cromosomi di ciascuna
coppia si attaccano mediante il cinetocore ai microtubuli
che raggiungono il polo opposto del fuso. Cioè entrambi i
cromatidi fratelli di un cromosoma omologo si attaccano ai
microtubuli che raggiungono un polo del fuso, mentre entrambi i cromatidi dell’altro cromosoma omologo si attac226
UNITÀ DUE
GENETICA
ribosomale). Tutti e cinque questi geni
sono localizzati anche sul cromosoma X.
Le proteine codificate dagli altri 7 geni
non mostrano alcuna relazione con proteine già note. Tuttavia, la maggior parte
contiene una sequenza di aminoacidi caratteristica di proteine che si legano a
sequenze di DNA o di RNA o alla cromatina (le fibre cromosomiche di DNAproteine). Queste caratteristiche suggeriscono che le proteine codificate da
questi geni possono regolare altri geni o
stabilizzare il DNA, l’RNA o la cromatina; pertanto, essi potrebbero avere un
ruolo importante nei processi che portano alla produzione di spermatozoi maturi.
Le ricerche di Lahn e Page dimostrano che le regioni del cromosoma Y
una volta considerate deserti in termini
genetici in effetti contengono geni attivi,
alcuni dei quali potrebbero essere necessari per la normale fertilità maschile.
L’identificazione delle funzioni di questi
geni potrebbe portare a trattamenti con
effetti benefici nella cura della infertilità
maschile causata dal loro funzionamento
difettoso.
cano ai microtubuli che si dirigono verso l’altro polo del
fuso.
Metafase I e anafase I. Alla prima metafase i movimenti dei
microtubuli del fuso determinano la collocazione delle tetradi sul piano equatoriale – la piastra metafasica – collocato
tra i due poli del fuso (Figura 11.3, stadio 5). Quindi i due
cromosomi omologhi di ciascuna coppia si separano e migrano verso i poli opposti del fuso, verso i quali sono trasportati dai microtubuli del fuso, che si contraggono durante l’anafase I (Figura 11.3, stadio 6). Questi movimenti
portano alla separazione (segregazione) degli omologhi e
determinano la migrazione a ciascun polo del fuso di un
corredo aploide di cromosomi. Comunque, tutti i cromosomi sono ancora costituiti in questo stadio da due cromatidi fratelli.
Raramente la segregazione cromosomica va incontro
ad errori. Per esempio, entrambi i cromosomi omologhi
di una coppia possono migrare allo stesso polo del fuso all’anafase I. Ne risulta una cosiddetta non-disgiunzione,
nella quale il fuso non riesce a separare i cromosomi omologhi. Tutta la tetrade migra allo stesso polo, che in tal
modo riceve entrambi i cromosomi di una coppia di omologhi, mentre all’altro polo non giunge alcun membro di
quel cromosoma. Lo zigote che ne risulta dopo la fecondazione avrà pertanto tre copie di un cromosoma invece
delle normali due, a causa della non-disgiunzione. Nell’uomo la maggior parte di questo tipo di zigoti non conclude lo sviluppo e quindi non origina un nuovo organismo. Un’eccezione è costituita dalla sindrome di Down,
che è caratterizzata dalla presenza di tre copie del cromosoma 21. La sindrome di Down comporta modificazioni
caratteristiche dell’aspetto del viso e del corpo, ritardo
mentale e una significativa riduzione della fertilità (vedi
Capitolo 13 per una descrizione più dettagliata della sindrome di Down).
Telofase I e intercinesi. La telofase è uno stadio breve,
transitorio, nel quale avvengono ben pochi cambiamenti a
carico dei cromosomi (Figura 11.3, stadio 7). L’involucro
nucleare si ricostituisce in alcune specie, ma non in altre.
La telofase è seguita da una intercinesi durante la quale il
fuso della prima divisione meiotica scompare e i microtubuli si riassemblano a formare due nuovi fusi per la seconda divisione.
Profase II, prometafase II e metafase II. La seconda divisione meiotica o meiosi II è un processo simile alla divisione mitotica. Durante la profase II, i cromosomi si condensano e si forma il fuso (Figura 11.3, stadio 8). Durante la
prometafase II, l’involucro nucleare si frammenta, il fuso si
porta nell’area precedentemente occupata dal nucleo e i microtubuli del fuso, che si dirigono verso i poli opposti del
fuso stesso, si attaccano ai due cinetocori di ciascun cromosoma bicromatidico (Figura 11.3, stadio 9). Alla metafase II,
i movimenti dei microtubuli del fuso determinano l’allineamento dei cromosomi sulla piastra equatoriale o metafasica
(Figura 11.3, stadio 10).
Anafase II e telofase II. L’anafase II inizia con la separazione
dei due cromatidi di ciascun cromosoma, che vengono trasportati ai poli opposti del fuso dai microtubuli (Figura 11.3,
stadio 11). Al termine della anafase II i cromatidi, a questo
punto di nuovo chiamati cromosomi, sono distribuiti ai
due poli. Durante la telofase II, si decondensano per acquisire la forma distesa tipica della interfase, il fuso si dissocia e
scompare e un nuovo involucro nucleare si forma attorno
alla massa di cromatina (Figura 11.3, stadio 12). Il risultato è
che si formano quattro cellule aploidi, ciascuna con un
nucleo contenente la metà del numero di cromosomi presente nella cellula progenitrice allo stadio G1 dell’interfase
iniziale.
Cromosomi sessuali e meiosi. In molti eucarioti, compresa
la maggior parte degli animali, una o più coppie di cromosomi, detti cromosomi sessuali o eterocromosomi, sono
diversi nei maschi e nelle femmine della stessa specie. Per
esempio, nell’uomo le cellule delle femmine hanno una
coppia di cromosomi XX (i cromosomi sessuali sono rappresentati nella Figura 10.7). I maschi contengono invece
una coppia di cromosomi sessuali costituita da un cromosoma X e da un cromosoma più piccolo detto Y. I due cromosomi X delle femmine sono completamente omologhi,
mentre l’X e l’Y del maschio sono omologhi solo per un
corto segmento. Comunque, un cromosoma X della madre
può appaiarsi sia con l’X che con l’Y paterno e le due diverse
coppie che possono formarsi si comportano durante le divisioni meiotiche come le altre coppie di cromosomi omologhi (autosomi).
Al termine della meiosi il gamete che si origina nelle
femmine contiene comunque uno dei due cromosomi X. Il
gamete maschile può invece contenere o il cromosoma X o
quello Y. (L’inserto Spunti dalla rivoluzione molecolare illustra in dettaglio gli studi che hanno portato alla scoperta del
gene essenziale per la determinazione del sesso sul cromosoma Y dei mammiferi).
La sequenza degli stadi e degli eventi delle due divisioni
meiotiche permette la realizzazione dei risultati più significativi della meiosi: la produzione di variabilità genetica e la
riduzione del numero di cromosomi. (La Figura 11.4 riassume le due divisioni meiotiche e le confronta con la singola divisione mitotica).
INTERVALLO
1. Quali sono le principali differenze tra i prodotti ottenuti dalla meiosi e dalla mitosi?
2. In cosa consiste la ricombinazione e in quale stadio
della meiosi avviene?
3. Quale delle due divisioni meiotiche è simile alla mitosi?
11.2 I meccanismi che producono
variabilità genetica
La produzione di variabilità genetica è un vantaggio fondamentale della riproduzione sessuale. La variabilità aumenta
la probabilità che almeno alcuni neonati sopravvivano e
raggiungano con successo la maturità riproduttiva se cambiano le condizioni ambientali.
La variabilità prodotta dalla riproduzione sessuale è
ben visibile e nota a tutti noi, specie se consideriamo il genere umano. A parte i gemelli identici, non esistono due
persone che siano uguali, si comportino allo stesso modo
o abbiano identiche caratteristiche biochimiche e fisiologiche, neppure se appartengono alla stessa famiglia. Allo
stesso modo, altre specie che si riproducono per via sessuale
presentano una variabilità simile che si origina dai processi
meiotici.
Nel corso della meiosi e della fecondazione la variabilità genetica è prodotta in particolare da tre meccanismi: (1)
la ricombinazione, (2) le diverse combinazioni di cromosomi di origine materna e paterna che segregano ai poli del
fuso durante la anafase I, (3) i particolari assetti cromosomici dei gameti maschili e femminili che si uniscono alla fecondazione. I tre meccanismi agiscono insieme e producono una variabilità complessiva così elevata che due gameti prodotti dallo stesso individuo, o da individui diversi,
o due zigoti prodotti dall’unione dei gameti non hanno alcuna probabilità di avere la stessa costituzione genetica.
Queste tre fonti di variabilità sono trattate in dettaglio nei
prossimi paragrafi.
La variabilità prodotta dalla ricombinazione
è conseguente all’appaiamento cromosomico
e agli scambi fisici che avvengono
tra cromatidi omologhi
La ricombinazione è l’evento chiave della profase I e inizia
con l’appaiamento dei cromosomi omologhi (Figura 11.5,
stadio 1). Quando si appaiano, i cromosomi omologhi
sono tenuti insieme da una struttura chiamata complesso
sinaptinemale (Figura 11.6). Grazie a questa struttura i segmenti omologhi dei cromatidi materno e paterno si scambiano dei tratti, producendo così nuove combinazioni di
CAPITOLO 11
MEIOSI: LE BASI CELLULARI DELLA RIPRODUZIONE SESSUALE
227
alleli (Figura 11.5, stadio 2). Il processo di scambio è molto
preciso e coinvolge la rottura e il ricongiungimento di porzioni di DNA operati da specifici enzimi. Ogni evento di
ricombinazione coinvolge due dei quattro cromatidi che
costituiscono la tetrade, mentre gli altri due cromatidi non
sono impegnati in quell’evento. Al termine della meiosi,
ciascuno dei quattro nuclei prodotti dalle due successive
divisioni riceve uno solo dei quattro cromatidi di ciascun
cromosoma (Figura 11.5, stadio 3); due ricevono cromatidi
non modificati, e gli altri due ricevono cromatidi con nuove combinazioni di alleli a seguito dello scambio. Quando
gli eventi di ricombinazione sono terminati, verso la fine
della profase I, il complesso sinaptinemale si dissocia e
scompare.
I siti dove è avvenuto lo scambio sono visibili più tardi
nella profase I, quando la progressiva condensazione dei
cromosomi determina un loro ispessimento e li rende visibili al microscopio ottico (vedi Figura 11.3, stadi 3 e 4). Questi siti, chiamati crossover o chiasmi (al singolare chiasma),
mostrano chiaramente che due dei quattro cromatidi si
sono scambiati dei segmenti nel processo di ricombinazione, che è detto anche crossing-over.
La ricombinazione può avvenire casualmente in quasi
ogni punto dei cromosomi, tra due qualsiasi dei quattro
cromatidi, a due a due omologhi. La stessa coppia di cromosomi omologhi può subire altri (uno o più) eventi di ricombinazione, che possono coinvolgere gli stessi due cromatidi che hanno subito scambio nel primo evento oppure
cromatidi diversi. Nella maggior parte delle specie si possono avere in media da due a tre scambi per ogni corredo di
cromosomi omologhi.
La segregazione casuale dei cromosomi
materni e paterni costituisce la seconda fonte
principale di variabilità genetica
durante la meiosi
La segregazione casuale dei cromosomi di origine materna
e paterna rappresenta la seconda importante fonte di variabilità genetica prodotta dalla meiosi. È opportuno ricordare
che la metafase I è lo stadio della meiosi nel quale le coppie
di cromosomi omologhi si attaccano alle fibre del fuso. I
cromosomi materni e paterni di ciascuna coppia contengono tipicamente alleli diversi di molti dei geni localizzati
su quel cromosoma. Un membro di ogni coppia di cromosomi omologhi viene trasportato dalle fibre del fuso ad un
polo, l’altro al polo opposto. Durante l’aggancio alle fibre
del fuso tutti i cromosomi materni possono migrare ad un
polo e tutti quelli paterni al polo opposto. O, come è ovvio,
si può avere con la stessa probabilità ogni tipo di possibile
combinazione di cromosomi materni e paterni ad ogni polo
(Figura 11.7). La seconda divisione meiotica non fa altro che
distribuire nei nuclei dei gameti queste diverse combinazioni di cromosomi.
Il numero totale di possibili combinazioni di cromosomi materni e paterni dipende dal numero di coppie di
cromosomi di ogni specie, cioè dal suo corredo aploide. Per
esempio, nell’uomo, che ha 23 coppie di cromosomi, si
possono avere complessivamente 223 diverse combinazioni
di cromosomi materni e paterni che migrano ai poli del
fuso e che portano alla possibilità di produrre 8.388.608 tipi
di gameti geneticamente diversi, solo in virtù di questa fonte di variabilità.
Figura 11.4
Confronto tra gli stadi fondamentali della meiosi e della mitosi.
In entrambi i disegni sono rappresentati esempi di cellule animali.
I cromosomi materni sono rossi, quelli paterni blu.
Meiosi I
Cellula diploide (2n)
Siti di ricombinazione
Profase I/prometafase I
Metafase I
I cromosomi duplicati si condensano,
si appaiano e si scambiano
tratti omologhi, quindi si attaccano
alle fibre del fuso.
Ciascun cromosoma materno
(costituito da due cromatidi fratelli),
al pari del suo omologo di origine
paterna, si dispone casualmente
nella regione mediana del polo.
228
UNITÀ DUE
GENETICA
Anafase I
I cromosomi omologhi (ciascuno
costituito da due cromatidi fratelli)
si separano e migrano verso i poli
opposti del polo (segregano).
Telofase I
Si formano due nuclei aploidi (n).
L’unione casuale dei gameti maschile
e femminile alla fecondazione produce
ulteriore variabilità genetica
I gameti maschili e femminili prodotti dalla meiosi sono tra
loro geneticamente diversi. L’unione dei due gameti alla fecondazione è un fenomeno del tutto casuale e questo amplifica ulteriormente la variabilità prodotta dalla riproduzione sessuale. Solo considerando la variabilità che si ottiene dalla segregazione casuale dei cromosomi omologhi
alla prima profase meiotica e quella prodotta dall’unione
casuale dei gameti alla fecondazione, la possibilità che due
figli della stessa coppia possano ricevere le stesse combina-
zioni di cromosomi materni e paterni è uguale a 1/(223)2 ,
cioè uno su 70.368.744.000.000 (circa 70mila miliardi), un
numero ben più alto di tutta popolazione umana mondiale.
L’ulteriore variabilità dovuta al fenomeno della ricombinazione rende praticamente impossibile per l’uomo e per gli
altri organismi che si riproducono sessualmente la produzione di gameti e figli geneticamente identici. L’unica eccezione è quella dei gemelli monozigoti o identici, che peraltro non derivano dall’unione di gameti identici alla fecondazione, ma dalla separazione di cellule embrionali (blastomeri) ottenute a seguito di divisioni mitotiche dell’unica
cellula uovo fecondata (zigote), che sviluppandosi separatamente danno origine a individui geneticamente uguali.
Mitosi
Cellula diploide (2n)
Cromatidi fratelli
Cromosomi
omologhi
Profase/prometafase
Metafase
I cromosomi duplicati (costituiti
da cromatidi fratelli) diventano visibili
in seguito alla condensazione e si
attaccano alle fibre del fuso.
I cromosomi si allineano
individualmente nella regione
mediana del fuso (piastra metafasica).
Anafase
I cromatidi fratelli di ciascun
cromosoma migrano verso i poli
opposti del fuso.
Telofase
Si formano due nuclei diploidi (2n)
e la citodieresi produce due cellule
figlie diploidi.
Meiosi II
Profase II/prometafase II
Ogni coppia di cromatidi fratelli si
attacca alle nuove fibre del fuso.
Metafase II
Anafase II
Telofase II
I cromosomi si allineano individualmente
nella regione mediana del fuso.
I cromatidi fratelli di ciascun cromosoma
migrano verso i poli opposti.
Si formano quattro nuclei aploidi (n)
e la citodieresi produce quattro cellule
aploidi.
CAPITOLO 11
MEIOSI: LE BASI CELLULARI DELLA RIPRODUZIONE SESSUALE
229
Figura 11.5
Coppia di omologhi
Effetti dello scambio tra due cromatidi
che determina la ricombinazione genetica. Le lettere indicano due alleli, A e a, di
un gene e due alleli, B e b, di un altro gene.
I genitori presentano gli alleli nelle combinazioni A-B e a-b (dette forme cis); a seguito della ricombinazione due dei cromatidi, detti ricombinanti, presentano le
nuove combinazioni alleliche a-B e A-b
(dette forme trans).
Cromatidi
fratelli
A
A
Cromatidi
fratelli
a
a
A
A
a
a
A
a
A
a
B
B
b
b
B
B
b
b
Crossover
(chiasma)
B
B
b
b
1 Coppia di
cromosomi omologhi.
2 I cromatidi omologhi si
scambiano dei tratti.
3 I cromosomi omologhi si separano
alla prima divisione meiotica.
Cromatidi fratelli di un membro di
una coppia di cromosomi omologhi
11.3 Momento e sede
della meiosi nel ciclo vitale
degli organismi
Per gent. conc. di Diter von Wettstein
Complesso
sinaptinemale
Cromatidi fratelli dell’altro membro di
una coppia di cromosomi omologhi
Fibre di cromatina
dei cromatidi
Figura 11.6
Il complesso sinaptinemale di una cellula in meiosi del fungo Neotiella. È illustrata la relazione tra il complesso sinaptinemale e le fibre cromosomiche dei cromosomi omologhi appaiati.
INTERVALLO
1. Quali sono i tre meccanismi attraverso i quali la riproduzione sessuale produce variabilità genetica?
2. Prendi in considerazione un animale con sei coppie di
cromosomi; un corredo aploide di 6 cromosomi deriva dal padre, l’altro corredo aploide omologo di 6
cromosomi dalla madre. Quanti gameti (in percentuale) prodotti da questo animale potranno contenere
cromosomi tutti di origine materna?
230
UNITÀ DUE
GENETICA
Il momento e la sede in cui avviene la meiosi durante il
ciclo vitale degli eucarioti seguono tre modelli principali (Figura 11.8). Le differenze riguardano: a) i periodi
del ciclo vitale trascorsi dagli organismi nella fase
aploide o diploide; b) se tra la meiosi e la formazione
dei gameti si hanno divisioni mitotiche.
Negli animali predomina la fase diploide,
mentre la fase aploide è ridotta
e la formazione dei gameti
segue direttamente la meiosi
Gli animali seguono il modello nel quale la fase diploide predomina nel ciclo vitale (Figura 11.8a), la fase
aploide è ridotta e la meiosi è immediatamente seguita
dalla formazione dei gameti. Nei maschi degli animali
ciascuno dei quattro nuclei prodotti dalla meiosi è contenuto in una cellula a seguito della divisione citoplasmatica (citodieresi) e ciascuna della quattro cellule si
differenzia in uno spermatozoo funzionale. Nelle femmine solo uno dei quattro nuclei diventa funzionale
all’interno dell’unica cellula uovo che si forma al termine della meiosi.
La fecondazione ripristina la fase diploide del ciclo vitale. Pertanto, gli animali hanno un corredo
aploide di cromosomi solo nei loro gameti maturi e le
loro cellule aploidi non vanno incontro a divisioni
mitotiche.