MICROBIOLOGIA AGROALIMENTARE
Batteriologia Generale
Prof.ssa Roberta Gaziano
[email protected]
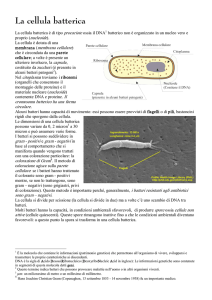
LA CELLULA PROCARIOTICA
Manca:
membrana nucleare
reticolo endoplasmatico
apparato del Golgi
mitocondri
LA CELLULA PROCARIOTICA
Il cromosoma batterico è direttamente immerso
nel citoplasma
Il citoplasma è delimitato esternamente da una
membrana citoplasmatica dalla quale si
dipartono verso l’interno delle invaginazioni che
formano il sistema dei mesosomi
LA CELLULA PROCARIOTICA
Nel citoplasma sono contenuti i ribosomi formati da
RNA e proteine, costituiti da due subunità: 30S e 50S
in base alla costante di sedimentazione
In alcune specie sono presenti flagelli (locomozione)
e pili (adesione e coniugazione)
LA MEMBRANA CITOPLASMATICA
E’ formata da : lipidi (40%)
proteine (60%)
piccole quantità di carboidrati
E’ coinvolta negli scambi metabolici tra citoplasma
ed ambiente esterno
E’ sede di processi biosintetici (sintesi del
peptidoglicano) e degli enzimi della catena
respiratoria, che nelle cellule eucariotiche hanno
sede nei mitocondri
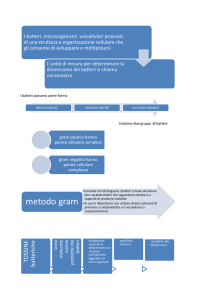
LA PARETE CELLULARE
Esternamente alla membrana c’è la parete
cellulare sulla cui superficie si trova spesso uno
strato polisaccaridico che costituisce la capsula
La parete è presente anche nelle cellule vegetali e
nei miceti ma quella dei batteri è più complessa e
distingue i batteri in Gram+ e Gram –
LA PARETE CELLULARE
E’ costituita da peptidoglicano, un polimero di due zuccheri
che si alternano: N-acetilglucosamina ed acido muramico
Ogni molecola di acido muramico lega un tetrapeptide
costituito nell’ordine da:
L-alanina
Acido D-glutamico
Acido meso-diamino-pimelico o L-lisina
D-alanina
La struttura polimerica del peptodoglicano è responsabile
della rigidità della parete
LA PARETE CELLULARE
STRUTTURA ESTERNA DEI GRAM +
Nei batteri Gram+ la membrana citoplasmatica è
ricoperta da uno spesso strato di peptidoglicano che
costituisce la parete cellulare ed eventualmente dalla
capsula
PARETE DEI BATTERI GRAM +
PARETE DEI BATTERI GRAM +
Ha uno spessore di 200-800 A° ed oltre al
peptidoglicano ha intercalati altri polimeri quali gli
acidi teicoici che possiedono elevata attività
antigenica
Le cariche degli aminozuccheri e degli aminoacidi e i
radicali fosforici degli acidi teicoico, rendono la parete
batterica altamente polare lasciando passare le
molecole idrofile respingendo quelle idrofobe (es. sali
biliari)
Nella colorazione di Gram i batteri Gram+ rimangono
colorati con il primo colorante (Cristalvioletto)
STRUTTURA ESTERNA DEI GRAM Nei batteri Gram- la membrana cellulare è
ricoperta da un sottile strato di
peptidoglicano che costituisce la parete e
da una membrana esterna.
Lo spazio compreso tra la membrana
cellulare e la membrana esterna
costituisce lo spazio periplasmatico
PARETE DEI BATTERI GRAM Ha uno spessore da 10 a 25 volte più sottile di quella
dei Gram+ e non ha intercalati gli acidi teicoici
La membrana esterna alla parete batterica
denominata lipopolisaccaride batterico (LPS)
estremamente tossico per l’uomo e per gli animali,
causa dello shock settico in caso di batteriemia da
batteri GramLPS viene detto anche ENDOTOSSINA
Anche nei Gram- i polisaccaridi dell’LPS rendono la
parete una struttura polarizzata
PARETE DEI BATTERI GRAM Il lipopolisaccaride batterico (LPS) è formato da:
• una porzione lipidica denominata lipide A
• una corta catena di zuccheri che forma il “core”
• una lunga catena polisaccaridica con attività
antigenica (antigene O)
•Antigene O: polisaccaride
lineare formato da 50-100
unita’ saccaridiche, permette
di distinguere tra sierotipi
(ceppi) di una stessa specie
batterica. Fornisce una
notevole compatezza al
batterio
•Il lipide A, formato
da un disaccaride
di glucosammina
fosforilata e acidi
grassi, è
responsabile
dell’attivita’
endotossica
dell’LPS
Lipopolysaccharide
n
O-antigen
Highly variable
Core
• Heptoses
• Ketodeoxyoctonic acid
Lipid A
• Glucosamine disaccharide
• Beta hydroxy fatty acids
•Core: polisaccaride
ramificato essenziale per la
struttura del batterio
PARETE DEI BATTERI GRAM -
CAPSULA
All’esterno della parete cellulare
Ben definita: capsula
Non definita: glicocalice
Generalmente polisaccaridica
Spesso assente in vitro
Presente in vivo e protettiva
CAPSULA
• Formata da mucopolisaccaridi secreti dalla cellula
batterica
• Ha funzione antifagocitaria
• Alcuni batteri possiedono la capsula in vivo ma non la
producono in vitro
• Si evidenzia con inchiostro di china
CAPSULA
FUNZIONI DELLA CAPSULA
-Adesione
-Inibizione fagocitosi e killing intracellulare
Glicocalice
Il glicocalice gioca un ruolo importante nella formazione dei
biofilm batterici.
I biofilm sono costituiti da microcolonie di cellule batteriche
adese ad una superficie incapsulate in una matrice
polisaccaridica (glicocalice) secreta dalle cellule stesse
Struttura del biofilm
La stuttura di un biofilm maturo è molto complessa:
Le microcolonie sono organizzate in agglomerati microbici interrotti da
canali per il passaggio dell’acqua per apporto di nutrienti ed eliminazione
delle scorie
Biofilm di S.epidermidis su corpi estranei
pacemaker
catetere
COLORAZIONI
Semplici
Differenziali
(cristalvioletto, fuxina basica, blu
di metilene)
(Gram, Ziehl-Neelsen)
Per colorare cellule eucariotiche si usano coloranti
basici per evidenziare il nucleo (basofilo) e coloranti
acidi per evidenziare il citoplasma (acidofilo)
Per colorare la cellula batterica si usano solamente
coloranti basici in quanto è uniformemente basofila
Tutti i campioni devono essere fissati (calore,
fissativi chimici) prima di essere colorati
COLORAZIONE DI GRAM
Il campione fissato viene colorato nel seguente modo:
• Coprire il preparato con cristalvioletto (2-3 min)
• Mordenzare* con liquido di Lugol = soluzione di iodio e
ioduro di potassio in acqua (1 min)
• Decolorare con alcool etilico o acetone (1-2 min)
• Coprire il preparato con fucsina o safranina
• Lavare con acqua
*
Mordente viene detta una sostanza che forma un composto insolubile con il
colorante rendendone più stabile l’unione al substrato
COLORAZIONE DI GRAM
Gram+ risultano colorati in violetto
Gram– risultano colorati in rosso
COLORAZIONE DI GRAM
PARETE DEI BATTERI ALCOOLACIDO RESISTENTI
I micobatteri possiedono una parete spessa
che contiene oltre al peptidoglicano e acidi
teicoici anche una grande quantità di cere
Per evidenziarli viene usata la colorazione di
Ziehl-Neelsen
Una volta fatto penetrare il colorante con
l’aiuto di una breve esposizione al calore, non
possono essere decolorato dal trattamento
con alcooli o acidi minerali
Da qui la definizione di batteri alcool-acido
resistenti
FLAGELLI
Sono formati da un unico filamento sprovvisto di
membrana costituito da subunità proteiche di una
peculiare proteina denominata flagellina con
attività antigenica, denominata antigene H
FLAGELLI
Batteri privi di flagelli
atrichi
Batteri con flagelli polari
monotrichi (un flagello ad un polo della
cellula
lofotrichi (un ciuffo di flagelli ad un polo
della cellula)
anfitrichi (un flagello o un ciuffo di
flagelli a entrambi i poli della cellula
Batteri con flagelli attorno
peritrichi
Flagelli
FLAGELLI: strutture a forma di
corda, formati da subunità proteiche
avvolte ad elica (flagellina) ancorati
alle membrana batterica.
PILI
Possiedono lunghezza compresa tra 0,2-2m e sono specifici
organi di ancoraggio
Sono responsabili dell’iniziale processo infettivo e della proprietà
emoagglutinante dei batteri
Sono formate dalla ripetizione di subunità di una o due proteine
(piline) con funzione di adesione (adesine) a particolari substrati
Una classe particolare di pili, di dimensioni maggiori rispetto a
quelli con funzione di adesine, è rappresentato dai pili F coinvolti
nel processo di coniugazione batterica
PILI o FIMBRIE: strutture
proteiche (pilina), lineari, a
centinaia intorno al batterio
che favoriscono l’adesività
(nomi alternativi: lectine,
evasine, aggressine)
E. coli with fimbriae
Pili o fimbriae (frange)
e cilia
Come fattore di adesività
le punte delle fimbrie
contengono delle
proteine (lectine) che
legano specifici zuccheri
(e.g., mannosio) delle
glicoproteine o glicolipidi
espressi sulla membrana
delle cellule epiteliali
Funzione dei pili:
adesione.
WEB PAGE FOR DR. KAISER 'S MICROBIOLOG Y COURSE (BIO L 230) THE COMMUNITY COLLEGE OF BALTIMORE COUNTY,
CATONSVI LLE CAMPUS.
Copyright ゥ1995 -2005 Gar y E. Kaiser All Rights Reserved Updated: Feb. 27, 2006
Adesività di E. coli alle cellule della
mucosa urinaria
Pili F, AFAI e AFAIII e
adesina Dr
ADESIVITA’ MEDIANTE FIMBRIE
Pilo sessuale
o Pilo F
WEB PAGE FOR DR. KAISER 'S MICROBIOLOG Y COURSE (BIO L 230) THE COMMUNITY COLLEGE OF BALTIMORE COUNTY,
CATONSVI LLE CAMPUS.
Copyright ゥ1995 -2005 Gar y E. Kaiser All Rights Reserved Updated: Feb. 27, 2006
IL CROMOSOMA BATTERICO
E’ costituito da DNA, è il depositario dell’informazione
genetica ed è denominato NUCLEOIDE
E’ costituito da un numero di coppie di basi compreso tra
1 e 5 milioni
Ha struttura circolare ed è collegato alla membrana
batterica in corrispondenza di zone caratteristiche
La maggior parte dei batteri possiede anche altre
molecole di DNA circolare denominate PLASMIDI
PLASMIDI
Sia i batteri Gram+ che quelli Gram- possiedono in aggiunta
al cromosoma batterico un elevato numero di elementi
genetici accessori detti plasmidi
I plasmidi sono elementi genetici extracromosomici
formati da DNA bicatenario a struttura circolare costituito da
un numero di coppie di basi variabile da 1000 a 200.000
PLASMIDI
I plasmidi codificano per tutti i materiali necessari e
sufficienti per la duplicazione e ripartizione nelle due
cellule figlie ma possono anche codificare per materiali
richiesti dal batterio per sopravvivere in particolari “nicchie
ecologiche” quali, tossine, pili, adesine ed enzimi in grado
di conferire resistenza all’azione di farmaci antibatterici
detti plasmidi o fattori R
PLASMIDI CONIUGATIVI
Alcuni plasmidi detti plasmidi coniugativi codificano
per una serie di prodotti che vengono richiesti per
trasferire un plasmide da una cellula ad un’altra
attraverso un “ponte coniugativo” permettendo un
trasferimento orizzontale del plasmide (da una cellula
“donatrice” ad una cellula “accettrice”)
GENETICA BATTERICA
Scambio genetico tra batteri:
•Trasformazione
• Trasduzione
• Coniugazione
Trasformazione
E’ il trasferimento di informazioni genetiche per
mezzo di DNA extracellulare.
Morte cellulare --> rilascio di DNA --> DNA che penetra
nella cellula ricevente
• Adsorbimento
• Incorporazione
• Ricombinazione nel cromosoma della cellula ricevente
Acquisizione di nuovi caratteri da parte della cellula
ricevente
TRASFORMAZIONE
Esperimento di Griffith nel 1928
Trasformazione
Sviluppo dello stato di competenza:
La cellula libera nell’ambiente la proteina detta “fattore di
competenza”.
a) Interazione con appositi recettori di superficie della
cellula che produce il fattore.
b) Sintesi di un’ autolisina che digerisce parte della parete
cellulare esponendo la membrana in cui si espongono alcune
proteine come: la proteina DNA- binding ed una nucleasi
specifica
Trasformazione
Frammenti di DNA bicatenario presenti nell’ambiente si
legano alle proteine DNAbinding.
c) Un filamento viene digerito dalla nucleasi specifica.
d) Il filamento residuo, complessato ad alcune proteine
specifiche ed introdotto nella cellula dove per analogia di
basi si integra nel genoma batterico, spiazzando un parte
dei due filamenti della molecola originaria.
Trasduzione
Il trasferimento genetico di DNA tra batteri
mediato da fagi (virus batterici).
I fagi possono essere:
Fagi temperati (Ciclo lisogeno)
Fagi litici (Ciclo litico)
I fagi trasducenti sono
difettivi, perché il pezzo di
DNA batterico ha preso il posto
del pezzo di DNA fagico.
TRASDUZIONE GENERALIZZATA
1. Un fago litico adsorbe ad un
batterio sensibile.
2. Penetrazione del genoma fagico
nel batterio. Utilizzo dei sistemi
metabolici cellulari per la sintesi e
l’assemblaggio delle componenti
virali.
3. In alcuni casi, un frammento di
DNA batterico o un plasmide
possono essere erroneamente
inseriti in alcuni capsidi virali
(particelle trasducenti).
TRASDUZIONE GENERALIZZATA
4. Rilascio dei fagi batterici
in seguito a lisi batterica.
5.Il fago trasduttore adsorbe
ad un batterio sensibile.
6.Penetrazione del DNA
fagico.
7.DNA fagico ricombina
con il DNA della cellula
accettrice.
TRASDUZIONE SPECIALIZZATA
1-2. Un fago temperato
adsorbe al batterio sensibile
e vi inietta il suo genoma.
3.Il genoma fagico
ricombina con il nucleoide
batterico divenendo un
profago.
4.In presenza di stimoli
adeguati, un frammento del
DNA batterico viene escisso
come parte del genoma
fagico
TRASDUZIONE SPECIALIZZATA
5. Durante la replicazione fagica il
DNA batterico viene inserito nel
genoma fagico. Tutti i fagi
veicolano il frammento di DNA
batterico.
6.Il fago adsorbe ad un batterio
sensibile iniettandovi il suo
genoma.
7.Il genoma fagico contenente il
DNA trasdotto ricombina con il
nucleoide della cellula accettrice.
Coniugazione
La Coniugazione è un processo che avviene attraverso una
particolare struttura anatomica (pilo sessuale) posseduto dal
batterio donatore, che unisce le due cellule in accoppiamento.
• La cellula donatrice ha un plasmide che porta l’informazione
genetica che codifica la sintesi del pilo.
CONIUGAZIONE BATTERICA
E’ il trasferimento di materiale genetico da un batterio,
che possiede il plasmide F (F+), ad un altro che non lo
possiede (F-)
La coniugazione batterica avviene tra batteri F+, in cui il
plasmide F codifica anche per la produzione del pilo F,
che serve ad ancorare le cellula, e batteri FCellula Hfr–cellula contenente un plasmide F integrato
nel cromosoma (Hfr –high frequency of recombination)
Coniugazione nei Gram+
Enterococcus faecalis
•Produzione e rilascio di feromoni da parte della cellula
“accettrice” (femminile)
•Feromoni inducono produzione della sostanza
aggregante alla superficie della cellula “donatrice”
(maschile)
•Formazione di aggregati cellulari con trasferimento del
plasmide coniugativo
Elementi Genetici Trasponibili
Sequenze di Inserzione (IS)
Transposoni (Tn)
Gli elementi IS
Le IS sono unità autonome e codificano solo per le
proteine necessarie alla propria trasposizione.
800-2000 pb
Sono costituenti comuni di cromosomi e plasmidi
Transposoni
Definizione: Segmenti di DNA mobili in grado di traslocare
nell’ambito della stessa cellula.
•
•
•
•
•
>2000 pb
Presenti nei cromosomi batterici, fagici, plasmidici.
Frequenza di trasposizione da 1000-10.000.
Possono alterare l’organizzazione dei geni.
Capacità d’inserire un segmento di DNA uguale a se stesso in un
altro sito.
• Integrazione sito-specifica. Le estremità di ogni Tn presentano
sequenze IS.
• I Tn trasportano geni accessori.
TRANSPOSONI
METABOLISMO BATTERICO
E’ l’insieme delle reazioni chimiche che avvengono in un
batterio con produzione di energia (esoergoniche) o con
sottrazione di energia (endoergoniche)
Anabolismo = sintesi di nuova sostanza vivente
Catabolismo = degradazione dei costituenti organici
Classificazione degli organismi viventi in base
alle caratteristiche metaboliche fondamentali
BATTERI
Aerobi obbligati = si sviluppano solo in presenza di ossigeno
Aerobi facoltativi = si sviluppano con e senza ossigeno
Anaerobi obbligati = si sviluppano solo in assenza di ossigeno
Microaerofili = si sviluppano bene a bassa tensione di ossigeno
Metabolismo energetico.
Curva di crescita
A) Fase di latenza : detta anche di
ringiovanimento o fase di
adattamento metabolico .
• Non si hanno atti
moltiplicativi.
• La durata della fase di latenza
dipende dall’età dell’inoculo,
dallo stato fisiologico delle
cellule e dal terreno di coltura.
B) Fase di accelerazione positiva:
fase in cui i batteri iniziano a
dividersi, ma solo il 20% si
divide con un tempo di
generazione più lungo rispetto
al suo standard.
C) Fase esponenziale: o
trofofase o fase logaritmica:
fase in cui il 100% dei batteri
si divide , in maniera
asincrona. Crescita continua
dei batteri in tutti gli stadi di
divisione.
• Il batterio in questa fase
produce metaboliti primari.
D) Fase di accelerazione
negativa: Aumentano i
cataboliti tossici e le sostanze
nutritive si riducono. In
questa fase si dividono l’80%
dei batteri, mentre il 20%
inizia a morire.
E) Fase stazionaria o idiofase:
Questa è una fase dove il 50%
si divide mentre il resto
muore. In questa fase i batteri
producono i metaboliti
secondari ( antibiotici,
tossine, pigmenti,
batteriocine). I batteri
sporigeni durante la fase
stazionaria danno origine alle
spore, per cui il titolo vitale
può restare costante o
diminuire di poco.
F) Fase di declino: La
condizione stazionaria è
seguita dalla fase di
morte. Questa si deve alla
forte riduzione di
sostanze nutritive ed
all’aumento di sostanze
tossiche. I batteri
sporigeni non
raggiungono il declino.
TEMPERATURA
Batteri vengono classificati:
Psicrofili
T. 0 e 20°C
Mesofili
T. 15°C e 45°C
Termofili
T. 45°C e 70°C
Ipertermofili
T. > 70°C
SPORA
Esclusiva dei batteri Gram+ dei generi Bacillus e Clostridium
Le spore batteriche sono endospore, si formano per resistere
a condizioni ambientali avverse e si liberano per rottura della
cellula madre, che prende il nome di sporangio
Nei batteri del genere Bacillus il diametro della spora non
eccede quello della cellula batterica
Nei batteri del genere Clostridium invece il diametro della
spora è maggiore di quello della cellula batterica
Endospore.
Sporogenesi, processo di
formazione dell’endospora
Strutture protettiva in
cui il batterio
sopravvive in uno
stato di dormienza.
Prodotta solo dai
gram positivi.
E’ coinvolta nella
trasmissione di
alcune infezioni
umane
da P.R. Murray, K.S. Rosenthal, G.S. Kobayashi, M.A. Pfaller
Microbiologia
EDISES
STRUTTURA DELLA SPORA
Dall’esterno verso l’interno si trova:
ESOSPORIO = sottile, di natura lipoproteica, non sempre presente
TUNICA = spessa, di natura proteica, con numerosi gruppi S-S
CORTECCIA = spessa, laminare, formata da peptidoglicano ma con
un minore numero di legami trasversali. E’ ricca di acido dipicolinico
che rende la spora resistente al calore
NUCLEO = al centro della spora, contiene il citoplasma delimitato
dalla membrana citoplasmatica, i ribosomi e il nucleoide.
STRUTTURA DELLA SPORA
La spora
CARATTERISTI CHE
DELLA SPORA
Resiste alla penetrazione di
sostanze estranee
Resiste all’essiccamento,
alle radiazioni gamma e
ultraviolette
Sopravvive al calore (anche
> 100°C)
Resiste anche per anni
nell’ambiente esterno
WEB PAGE FOR DR. KAISER 'S MICROBIOLOG Y COURSE (BIO L 230) THE COMMUNITY COLLEGE OF BALTIMORE COUNTY,
CATONSVI LLE CAMPUS.
Copyright ゥ1995 -2005 Gar y E. Kaiser All Rights Reserved Updated: Feb. 27, 2006
GERMINAZIONE DELLA SPORA
In condizioni ambientali favorevoli (acqua,
Germinazione della spora
calore, sostanze nutritive), la spora
germina liberando la cellula batterica
•Clostridium tetani
•Bacillus anthracis
Bacillus anthracis: antrace
Clostridium tetani: tetano
Clostridium botulinum: botulismo
Clostridium perfringens: gangrena gassosa
WEB PAGE FOR DR. KAISER 'S MICROBIOLOG Y COURSE (BIO L 230) THE COMMUNITY COLLEGE OF BALTIMORE COUNTY,
CATONSVI LLE CAMPUS.
Copyright ゥ1995 -2005 Gar y E. Kaiser All Rights Reserved Updated: Feb. 27, 2006
SPORE CLOSTRIDIUM TETANI
Spore tetaniche
RIPRODUZIONE BATTERICA
I batteri si riproducono per
scissione binaria
RIPRODUZIONE BATTERICA
La riproduzione comporta:
•
•
•
•
Duplicazione del cromosoma batterico
Duplicazione del punto di attacco alla membrana
Accrescimento delle membrane batteriche
Allungamento della cellula batterica nella sua
parte centrale
• Allontanamento dei due cromosomi
• Formazione di un setto di separazione a livello
della membrana
• Formazione di un setto a livello della parete
RIPRODUZIONE BATTERICA
PATOGENESI BATTERICA
• Interazione
ospite-parassita:
• Commensalismo
• Simbiosi
• Mutualismo
• Parassitismo
WEB PAGE FOR DR. KAISER 'S MICROBIOLOG Y COURSE (BIO L 230) THE COMMUNITY COLLEGE OF BALTIMORE COUNTY,
CATONSVI LLE CAMPUS.
Copyright ゥ1995 -2005 Gar y E. Kaiser All Rights Reserved Updated: Feb. 27, 2006
Infezione: Sta a significare la moltiplicazione dei microrganismi
all’interno dell’ospite senza danno. Non è sinonimo di malattia.
Malattia: sta a significare la moltiplicazione dei microrganismi
nell’ospite e danno conseguente.
Patogenicità: capacità di un microrganismo di causare danno.
Il livello di patogenicità dipende da due fattori:
Patogenicità
Virulenza: grado di patogenicità, dato dalla
capacità di moltiplicarsi in vivo e dalla invasività
(diffusione del patogeno nell’organismo)
Tossigenicità o produzione di tossine che
danneggiano il normale funzionamento
dell’organismo ospite
DANNO TISSUTALE
• La colonizzazione delle strutture esposte,
• Il superamento delle barriere,
• L’invasione dei tessuti profondi : comportano il danno anatomofunzionale dell’ospite.
• Responsabili del danno possono essere sia i mediatori dell’ospite
(pirogeni, enzimi lisosomiali, mediatori del sistema immune, etc..)
fattori intrinseci di virulenza dei microrganismi tra cui:
LE TOSSINE
che i
Tappe del processo infettivo
Principali fasi del
•Esposizione al patogeno
processo
patogenetico
(aria,acqua,
cibo, contatto
diretto,vettore)
•Adesione alla pelle o alle mucose
ESPOSIZIONE
ADESIONE
•Invasione attraverso l’epitelio.
•Colonizzazione e crescita (e/o
INVASIONE
completamento del ciclo biologico)
•Evasione delle difese immunitarie
dell’ospite
COLONIZZAZIONE
CRESCITA
•Capacità di danneggiare l’ospite
(tossicità).
TOSSICITA’
INVASIVITA’
DANNO AI TESSUTI
Rapporto ospite-microrganismo
La patogenicità è caratterizzata da:
– fattori e meccanismi di virulenza
– carica batterica (numero iniziale di
batteri infettanti)
– stato di salute, in particolare dalla
risposta immunitaria dell’ospite.
ADESIONE
ADESIONE è la conseguenza dell’interazione specifica tra batteri
patogeni e cellule dell’ospite
I batteri che sono dotati di adesine si legano a specifici recettori di
natura glicoproteica o glicolipidica presenti sulla superficie dei
tessuti e non vengono allontanati per esempio dal lavaggio dell’urina
o dalla peristalsi intestinale.
Le adesine sono molecole proteiche presenti all’apice di fimbrie o
pili che si legano a particolari zuccheri presenti sulla superficie dei
tessuti bersaglio.
FATTORI DI ADESIONE BATTERICA (adesine)
Fimbrie o Pili
Strutture filamentose utili ai batteri nell’attacco ad altri
microrganismi oppure a superfici solide
Capsula
Strato di fibre esopolisaccaridiche con un margine esterno
distinto che circonda molte cellule; inibisce la fagocitosi e
agevola l’adesione
Strato S
Strato più esterno, definito anche strato cristallino, costituito
da subunità proteiche a struttura regolare, presente in alcuni
batteri che promuove l’adesione alle superfici
Glicocalice
Acidi teicoici o
lipoteicoici
Pellicola batterica mucopolissacaridica meno compatta di una
capsula e facilmente asportabile
Componenti della parete cellulare dei batteri Gram-positivi che
agevolano l’adesione
INVASIONE
INVASIONE = è il passaggio successivo all’adesione e
consiste nell’internalizzazione del batterio nelle cellule
epiteliali dove i batteri possono essere liberi nel
citoplasma o contenuti all’interno di un vacuolo in cui
sopravvivono e si moltiplicano.
A
B
The bacterium (arrow in A) is beginning to enter the host cell by endocytosis.
Note the endocytic vacuole (arrow in B) containing the bacterium.
WEB PAGE FOR DR. KAISER 'S MICROBIOLOG Y COURSE (BIO L 230) THE COMMUNITY COLLEGE OF BALTIMORE COUNTY,
CATONSVI LLE CAMPUS.
Copyright ゥ1995 -2005 Gar y E. Kaiser All Rights Reserved Updated: Feb. 27, 2006
FATTORI DI VIRULENZA
BATTERICA
FATTORI ANTIFAGOCITARI
COMPETITORI PER IL FERRO (SIDEROFORI)
METABOLITI
ENZIMI DEGRADATIVI
TOSSINE
FATTORI ANTIFAGOCITARI
La capsula è il più importante fattore antifagocitario
I ceppi batterici dotati di capsula sono più virulenti e più resistenti alla
fagocitosi e alla lisi intracellulare dei ceppi non capsulati
Leucocidina - prodotta da alcuni ceppi di Staphylococcus aureus svolge
attività antifagocitaria in quanto capace di uccidere leucociti e macrofagi,
formando su queste cellule dei pori, che ne aumentano la permeabilità ai
cationi.
Enzima coagulasi - prodotta sempre da Staphylococcus aureus,
contribuisce all’attività antifagocitaria, formando uno strato di fibrina
attorno alla cellula batterica che non può venire fagocitata
COMPETITORI PER IL FERRO
(Siderofori)
Il ferro presente nel corpo umano è solo parzialmente
disponibile per il metabolismo batterico in quanto la maggior
parte è legato all’emoglobina negli eritrociti, alla transferrina
nel plasma, alla lattoferrina nelle lacrime, nella saliva e nel
muco bronchiale, nella bile e nel muco intestinale.
I batteri producono siderofori e proteine recettoriali sulla loro
superficie, capaci di riconoscere i siderofori stessi.
In questo modi i batteri possono sottrarre all’ospite ferro da
utilizzare per il loro metabolismo
Siderophores (S)
Receptor
Fe 2+/S
Fe 2+/S
METABOLITI
Molti dei prodotti metabolici derivanti dalla crescita
batterica rappresentano fattori di virulenza, capaci
di causare malattia
Acidi, gas ed altri prodotti del metabolismo batterico
possono risultare tossici per il tessuto
ENZIMI DEGRADATIVI
Sono capaci di dissolvere i tessuti e distruggere le barriere fisiche
che potrebbero limitarne la diffusione
Collagenasi:
degrada il collagene
fosfolipasi:
depolimerizza le membrane cellulari
proteasi:
degrada le IgA presenti sulle mucose
ialuronidasi:
liquefà l’acido ialuronico del connettivo
fibrinolisina:
elimina i coaguli di fibrina che si formano nel
processo infettivo per limitare la diffusione
batterica
lipasi:
degrada lo strato lipidico dell’epidermide
emolisina
lisa le emazie
TOSSINE BATTERICHE
ENDOTOSSINE = sono le componenti tossiche della
porzione
più
esterna della
parete
dei Gram-negativi
(lipopolisaccaridi – LPS)
parete
cellulare
ENDOTOSSI
NE
parete
cellulare
ESOTOSSINE = sono prodotti batterici tossici
che vengono rilasciati all’esterno dal batterio
(proteine) sia dai Gram+ che dai Gram-
ESOTOSSI
NE
Confronto
ESOTOSSINE
Natura proteica
Termolabili
Immunogeni
Neutralizzabili da anticorpi
Profili di azione molto differenziati
Estrema potenza
Alcune attive per ingestione
ENDOTOSSINE
Natura Lipopolisaccaridica
Termostabili
Scarsamente immunogeni
Non neutralizzabili da anticorpi
Profili di azione poco differenziati
Meno potenti
Inattive per ingestione
ESOTOSSINE
Le esotossine sono di natura proteica e rappresentano
importanti fattori di virulenza in malattie, come la
DIFTERITE, IL TETANO, IL BOTULISMO, IL COLERA ETC,
nelle quali la sintomatologia clinica è legata esclusivamente al
danno da esse prodotto.
Fortemente immunogene.
Mediante trattamento con formaldeide perdono il potere
tossico, conservando quello antigenico (anatossine), il che le
rende utilizzabili per la immunizzazione attiva (vaccinazione).
I vaccini a base di anatossina vengono utilizzati nella
profilassi attiva obbligatoria nei confronti di microrganismi
responsabili di gravi patologie nell’uomo, quali la difterite e il
tetano
CLASSIFICAZIONE DELLE ESOTOSSINE IN BASE
AL BERSAGLIO DI AZIONE
A seconda della presenza di specifici recettori cellulari
le tossine si distinguono in:
•
•
•
•
•
Tossine citolitiche o emolisine
Leucocidine
Tossine neurotrope
Enterotossine
Tossine pantrope
TOSSINE
CITOLITICHE
O
EMOLISINE
Danneggiano la
membrana cellulare provocando:
• Formazione di pori o canali che alterano la funzionalità
della cellula
• Distruzione enzimatica della porzione lipidica della
membrana cellulare
Formazione di pori o canali transmembrana
Alcune tossine citolitiche si inseriscono nel
doppio strato lipidico della membrana formando
dei pori o canali che mettono in comunicazione
diretta il citosol con l’ambiente esterno, alterando
la funzionalità della membrana.
Es. emolisine alfa di S. aureus, streptolisina O
di Streptococcus pyogenes = EMOLISI
Leucocidine
Sono tossine prodotte principalmente da Staphylococcus
aureus e Pseudomonas aeruginosa
Hanno azione citotossica esclusivamente nei confronti
dei macrofagi e dei leucociti polimorfonucleati.
Il meccanismo d’azione consiste in una stimolazione
irreversibile della Fosfolipasi C che provoca una massiva
mobilitazione di Ca ++.
A sua volta questo provoca la fosforilazione di una
proteina della membrana lisosomiale che perde la sua
struttura e libera gli enzimi nel citosol, con conseguente
morte cellulare.
NEUROTROPE ATTIVE SULLE CELLULE DEL SISTEMA NERVOSO
Le più importanti sono quella tetanica attiva sulle cellule del SNC, mentre
la botulinica è attiva su cellule del sistema periferico.
ENTEROTOSSICHE
ATTIVE SULLE CELLULE DELLA MUCOSA
INTESTINALE
Citotossiche: che causano danni evidenti alle cellule
Citotoniche, danni non evidenti.
PANTROPE ATTIVE SU TUTTE LE CELLULE, inibiscono la sintesi
proteica,Tossina difterica (prototipo). Inibisce la sintesi proteica e causa
necrosi dell'epitelio, tessuto nervoso, muscolo cardiaco e rene.
Possono essere classificate
in base alla loro attività
Tossine che agiscono a livello extracellulare:
la principale tossina che agisce a livello epidermico è
la tossina epidermolitica prodotta da Staphylococcus
aureus.
Tossine che agiscono sulle membrane cellulari:
sono le tossine citolitiche e le emolisine che portano
alla lisi cellulare attraverso diversi meccanismi.
Penetrano nelle cellule modificandone la regolazione
Penetrano nelle cellule uccidendole
TOSSINE
• Tossina Epidermolitica o Esfoliativa : è responsabile della
Sindrome stafilococcica della cute scottata (scalded skin
staphylococcical syndrome, SSSS)
• Sono proteasi delle serine e causano la rottura dei ponti
cellulari (desmosomi) nello strato granuloso dell’epidermide.
Causa anche shock tossico.
• La tossina induce la produzione di Ac neutralizzanti che
portano alla risoluzione del processo tossico (7-10gg).
Colpisce neonati e bambini di 1-3 anni.
Tossina esfoliativa
Esistono 2 forme della tossina:
ETA termostabile, cromosomica
ETB termolabile, plasmidica
Agisce sui desmosomi (giunzioni cellulari, mantengono la
forma e la consistenza dei tessuti)
ESOTOSSINE: struttura molecolare
.
• Tossine monomeriche costituite da una sola molecola proteica
(alcune).
• Tossine dimeriche formate da due diverse subunità peptidiche A-B
• Tossine multimeriche il componente B è un oligomero formato da
un certo numero di peptidi identici o diversi.
TOSSINE A-B
tossine binarie
Sono tossine che presentano analogie strutturali e
biochimiche.
Sono costituite da due componenti (2 peptidi):
subunità B (binding)/ subunità A (active)
La subunità B (binding) media il legame al recettore
della cellula ospite, favorendo la penetrazione della
subunità A (active), responsabile di attività
enzimatiche che si esplicano nella cellula bersaglio
con conseguente danno.
Subunità o peptide A
rappresenta la tossina vera e propria. L’attività enzimatica
della subunità A si può esprimere con diversi meccanismi che
vanno da un’attività ADP-ribosilante (es.tossina colerica,
pertossica, difterica) ad un’attività proteolitica (tossina
tetanica e botulinica).
Subunità o peptide B è responsabile della penetrazione
intracellulare del peptide A attivato.
B
A
B
A
I due peptidi sono legati da
ponti
disolfurici
che
vengono
scissi
dalle
proteasi dell’ospite. Se i
due
peptidi
vengono
inoculati
singolarmente
sono del tutto innocui .
Attività ADP- ribosilante
Molte tossine hanno attività ADP-ribosilante. Il processo
della ribosilazione consiste nell’aggiunta di un riboso
sulla proteina bersaglio attraverso una molecola di NAD.
È una modificazione post-traduttiva.
TOSSINA COLERICA
• 5 monomeri B circondano un monomero A
• Le parti B, reagendo con i recettori specifici,
modificano allostericamente il complesso,
traslocando attraverso la membrana cellulare il
polipeptide A, che si scinde in una parte
enzimatica attiva, ADP-ribosilante una proteina
G di membrana che controlla l’AMP ciclasi
Tossina Shiga
GB
A
globotriosilceramide
Vescicola
endocitosica
lisosomi
A1
A2
A1: frammento attivo
Legame al ribosoma e distacco di
una adenina a livello dell’RNAr 28S
Blocco sintesi proteica MORTE
TOSSINA DIFTERICA
Tossine (neurotrope) che interferiscono
con il rilascio di neurotrasmettitori:
TOSSINA TETANICA
Meccanismo d’azione della
tossina tetanica: blocco delle
sinapsi inibitorie della
contrazione muscolare
riflessa
TOSSINA BOTULINICA
Meccanismo di azione della
tossina botulinica: blocco della
trasmissione dell’impulso a livello
della giunzione neuromuscolare
ENTEROTOSSINE STAFILOCOCCICHE
Tossiche per ingestione
Intossicazioni alimentari
Resistono al riscaldamento 100°C per 30 min
Provocano febbre, nausea, vomito, diarrea
Ad elevate concentrazioni se rilasciate nel torrente
ematico causano la Sindrome da Shock Tossico (TSS)
ENTEROTOSSINE STAFILOCOCCICHE
Tossine Azione patogena
T A Intossicazione alimentare, stimola la peristalsi,
T B Colite pseudomembranosa stafilococcica,
40% TSS non mestruale
T C Intossicazione da prodotti caseari, TSS non
mestruale
T D Intossicazione alimentare
T E Occasionalmente intossicazione alimentare
T F o TSST-1 Pirogenica, 85% TSS mestruale
Sindrome da shock tossico
Inizialmente da tamponi mestruali colonizzati da
S. aureus produttore di TSS-1
Tamponi chirurgici o infezioni da S. aureus
produttori di varie enterotossine
Di recente da infezioni da S. pyogenes produttore
di Tossine pirogeniche
I superantigeni sono
proteine batteriche ad
azione toxin-like che
attivano le cellule T
CD4+
legandosi
aspecificamente
(senza presenza di Ag
specifico) al T cell
receptor (TCR) ed al
complesso maggiore
di istocompatibilità di
classe II (MHC-II) dei
macrofagi.
L’interazione aspecifica APC-linfocita T mediata dal superantigene si
traduce nella attivazione e proliferazione delle cellule T e
nell’attivazione dei macrofagi con rilascio di elevate dosi di diverse
citochine (IL-1, IL-8, TNF-α, PAF) coinvolte nella patogenesi dello
shock (danno endoteliale, CID, MOFS)
Sindrome da shock tossico
•Ceppi di S. aureus
produttori di tossina TSST-1
•Crescita localizzata nella
vagina (50% infezioni) o in
una ferita e rilascio della
tossina nel torrente
circolatorio
•Nel passato associata all’uso
di tamponi mestruali
iperassorbenti nei quali il
batterio si moltiplicava e
produceva tossina
INIBIZIONE DELLE ESOTOSSINE
Sull’inibizione delle tossine batteriche si basa la
risposta immunitaria attiva dei principali vaccini
antibatterici
Le tossine batteriche sono il principali bersaglio
della sieroprofilassi
ANATOSSINE
Le anatossine o tossoidi sono tossine proteiche
batteriche alle quali è stato artificialmente eliminato
il potere tossico preservando le proprietà antigeniche
Sono i costituenti dei principali vaccini antibatterici
(vaccino antitetanico, antidifterico e antipertosse)
CARATTERISTICHE DELLE
ENDOTOSSINE
ENDOTOSSINE = lipopolisaccaride – LPS
Termostabili
Scarso potere antigene (legato alla porzione polisaccaridica)
Potere tossico ( legato alla porzione lipidica ) non neutralizzato
dall’anticorpo corrispondente
Resistenti all’azione dei succhi gastrici
ENDOTOSSINE
Sono componenti tossiche esclusive della parete dei batteri
GramChimicamente sono lipopolisaccaridi (LPS) costituiti da una
porzione lipidica ed una polisaccaridica
L’endotossina lega specifici recettori presenti su macrofagi, cellule
dendritiche e stimola la produzione ed il rilascio di Citochine di
fase acuta quali IL-1, IL-12, IL-6, prostaglandine e Tumor
Necrosis Factor (TNF-a)
EFFETTI BIOLOGICI DELLE
ENDOTOSSINE
A basse concentrazioni attivano risposte protettive
•
Febbre
•
Vasodilatazione
•
Risposta immunitaria
•
Risposta infiammatoria
EFFETTI BIOLOGICI DELLE
ENDOTOSSINE
Ad alte concentrazioni determina l’insorgenza di fenomeni quali:
•
Febbre alta
•
Petecchie (dovute alla dilatazione dei capillari)
•
Ipotensione
•
Shock
(per
vasodilatazione
capillare)
•
Coagulazione del sangue
•
Morte
ed
aumentata
permeabilità
Le diverse attività
del lipopolisaccaride
LPS attiva quasi tutti
i meccanismi
di difesa del sistema
immunitario ed
anche il sistema della
coagulazione.
DIC = coagulazione
intravascolare
disseminata.
CID: coagulazione
intravascolare
disseminata
ARDS: Sindrome da
insufficienza
respiratoria acuta
MOSF: Sindrome da
insufficienza multipla
degli organi
Le endotossine sono induttori non
specifici dell’infiammazione;
Inducono il rilascio di citochine,
stimolano varie cellule a produrre
delle citochine, incluse IL-1 e TNF.
L’attività tossica è dose-dipendente
Attivano la via alternativa del
complemento.
PATOGENICITA’ DEL
PEPTIDOGLICANO
Il peptidoglicano ha attività per molti versi
simili a quelle dell’endotossina
Tossicità
mediata dal
Peptidoglicano



![5\) batteri [modalità compatibilità]](http://s1.studylibit.com/store/data/000928299_1-28a224c15810efb654d8d9e4b207aa31-300x300.png)