Michele Rismondo
[email protected]
Insegnamento di
BIOLOGIA, ANATOMIA E
MORFOLOGIA VEGETALE
Lezione 02: LA CELLULA VEGETALE
I COMPONENTI DELLA CELLULA VEGETALE
Parete cellulare:
Protoplasto:
Lamella mediana
Parete primaria
Parete secondaria
Plasmodesmi
Nucleo
Involucro nucleare
Nucleoplasma
Cromatina
Nucleolo
Citoplasma
Membrana plasmatica
Citosol
Plastidi
Mitocondri
Perossisomi
Vacuoli
Reticolo endoplasmatico
Apparato di Golgi
Vescicole
Citoscheletro
Microtubuli
Filamenti di actina
Ribosomi
Corpi oleosi
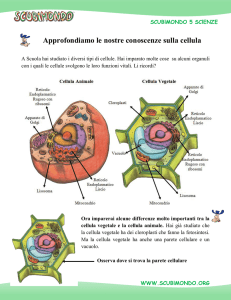
LA CELLULA VEGETALE
LA CELLULA VEGETALE
ORGANULI CITOPLASMATICI
Organuli coinvolti nelle trasformazioni energetiche
• Mitocondri
• Plastidi
– Cloroplasti
– Cromoplasti
– Leucoplasti
ORGANULI CITOPLASMATICI
• Le cellule hanno bisogno di energia per svolgere
le funzioni fondamentali (crescita, riproduzione,
movimento)
• L’energia viene trasformata nei mitocondri
(comuni a tutte le cellule eucariotiche) e
prodotta nei cloroplasti (esclusivi delle piante)
PLASTIDI
• Tutte le cellule vegetali vive hanno i plastidi (organuli presenti nelle cellule delle
piante e dei protisti)
• I plastidi derivano da un proplastidio (plastidio indifferenziato) presente nelle
cellule embrionali.
• Durante il differenziamento cellulare, in base al tessuto che andrà a formare e
alla funzione che andrà a svolgere la cellula, il proplastidio si trasforma in
plastidio adulto:
Cloroplasti
Fotosintesi clorofilliana
Cromoplasti
Pigmentazione
Leucoplasti
Riserva
Tutti i plastidi derivano dallo stesso proplastidio che ha la capacità
di trasformarsi in qualunque tipo di plastidio; questa sua capacità
è condizionata da:
Fattori esterni (luce, temperatura, acqua)
Fattori interni
I fattori interni prevalgono anche se i fattori esterni incidono.
Es. la clorofilla, nei cloroplasti, si forma solo in presenza della luce; i cloroplasti si
trovano solo nelle parti della pianta che sono esposte alla luce.
Foglia sviluppata al buio: i suoi proplastidi non diventano leucoplasti, ma sono
sempre cloroplasti incolori ed anomali
I proplastidi della radice nelle cellule adulte diventeranno leucoplasti (amiloplasti):
se esposta alla luce difficilmente diventeranno cloroplasti
EFFETTO IRREVERSIBILE DELLA DETERMINAZIONE
Il tipo di differenziamento dei proplastidi è già determinato prima che inizi.
Es.: frutto immaturo (rosa, peperone, pomodoro…) è
verde per la presenza cloroplasti; man mano che
matura, sempre in presenza della luce i cloroplasti si
trasformano in cromoplasti (degradazione clorofilla,
scomparsa tilacoidi e accumulo masse di carotenoidi)
CLOROPLASTI
I cloroplasti sono i più conosciuti tra i plastidi, devono il loro
nome alla presenza della clorofilla e all’attività fotosintetica che
rappresenta la più importante sorgente di energia.
Analogamente ai mitocondri sono
delimitati da un duplice sistema di
membrane.
La membrana interna dà origine ad
un complesso reticolo membranoso
la cui struttura varia da organismo a
organismo.
Un cm2 di foglia può contenere fino a
500.000 cloroplasti.
Forma e dimensione dei cloroplasti
Negli organismi più evoluti
(dai muschi in su) i
cloroplasti sono piccoli e
molto numerosi
I cloroplasti si dispongono
parallelamente
o
perpendicolarmente
alla
superficie fogliare in base
alla quantità di luce.
Nelle alghe i cloroplasti possono essere molto grandi e poco
numerosi
Struttura dei cloroplasti
b
a
Cloroplasti al microscopio ottico (a) e elettronico (b)
Ricostruzione tridimensionale della struttura interna
CROMOPLASTI
Presentano una specifica
colorazione sulla base dei
diversi pigmenti contenuti.
Cromoplasto di un petalo
di fiore contenente goccioline
lipidiche in cui si accumulano
i pigmenti carotenoidi
I cromoplasti accumulano al loro interno pigmenti carotenoidi spesso sciolti in
gocce di grasso
I pigmenti carotenoidi sono responsabili del colore giallo,
arancione o rosso di molti fiori, foglie senescenti, frutti e radici.
Radice di carota e
dettaglio
delle
cellule radicali con
cromoplasti
Petali di begonia e dettaglio del
cromoplasto responsabile della
loro colorazione
Non è ancora ben conosciuto il ruolo biochimico di questi organuli nel metabolismo
cellulare, anche se è evidente l’effetto attrattivo del colore verso gli animali
responsabili dell’impollinazione e della disseminazione
LEUCOPLASTI
Sono i plastidi meno differenziati, sono privi di tilacoidi e di
qualunque tipo di pigmento.
In essi vengono accumulate le sostanze di riserva, in genere amido
o, talora, goccioline di oli.
Leucoplasti in cellule della radice di carota (Daucus
carota) al microscopio ottico
I leucoplasti sono quindi costituiti da organuli deputati all’accumulo nella cellula
di sostanze di riserva, come amidi e sostanze lipidiche. Presenti negli organi
sotterranei, nei tessuti profondi non esposti alla luce degli organi aerei (fusti,
frutti, semi).
Prodotti di riserva dei leucoplasti
Cellula parenchimatica del tubero di
patata
contenente
all’interno
numerosi
granuli
di
amido
secondario
(immagine
al
microscopio
elettronico
a
scansione)
I prodotti della fotosintesi (glucosio) vengono condensati in amido all'interno del
cloroplasto; questo viene detto amido primario. L'amido primario viene poi
idrolizzato e trasportato, sotto forma di saccarosio (disaccaride del glucosio) fino ai
tessuti di riserva, dove viene ricondensato come amido secondario all'interno dei
leucoplasti, abbondanti in tali tessuti.
L’amido è la principale sostanza di riserva nelle piante. E’ un polisaccaride
costituito da molecole di alfa-glucosio legate insieme da legami glucosidici 1-4
L’ Amido
L'amido è un carboidrato polisaccaridico che consiste di un gran numero di unità di glucosio unite tra
loro da legame glicosidico. L'amido puro è una polvere bianca, insapore ed inodore, che risulta
insolubile in acqua fredda o in alcol. L'amido è prodotto dalle piante verdi, dove è utilizzato come
riserva nelle cellule vegetali, ed è un'importante fonte alimentare anche per l'uomo. La macromolecola,
quando è disciolta in acqua calda, può anche essere utilizzata in numerosi processi industriali come
agente addensante o collante. È composto da due polimeri: l'amilosio (che ne costituisce circa il 20%) e
l'amilopectina (circa l'80%). In entrambi i casi si tratta di polimeri del glucosio, che si differenziano l'uno
dall'altro per la struttura.
L'amilosio è un polimero lineare che tende ad avvolgersi ad elica, in cui le unità di glucosio sono legate
tra loro con legami glicosidici α(1→4).
L'amilopectina è invece un polimero ramificato che presenta catene di base di struttura simile
all'amilosio che si dispongono a formare una struttura ramificata; L'amilopectina è caratterizzata da
strutture amorfe e strutture cristalline; le prime sono costituite di legami α(1→6), mentre le strutture
cristalline danno origine a tratti di doppia elica. L'amilopectina è disposta verso l'interno in prossimità
del centro dei granuli di amido. Una molecola di amilosio può contenere sino a 1000 residui di glucosio.
L'amilosio lega insieme la struttura più espansa dell'amilopectina.
L’ Amido
Il contenuto in amilopectina dell'amido varia da una specie all'altra: infatti, si passa dal 78%
nel caso della patata, della banana e dei semi di alcuni cereali (come grano, riso e mais) al
99% nei semi di altri cereali.
L’ AMIDO E’ PRESENTE:
- in tutte le piante superiori ad eccezione di alcune composite
- 20-30% nei tuberi di patata
- 36% nei semi di ippocastano
- 52% nei semi di piselli
- 66% nei semi di orzo
- 70% nei semi di frumento
MANCA :
- NEI BATTERI E NELLE ALGHE AZZURRE
- NEI FUNGHI
- NELLE ALGHE (ad eccezione di alcuni gruppi)
L’ Amido
Biosintesi e Funzione :
L'amido è il carboidrato di riserva delle piante, immagazzinato come fonte energetica, sintetizzato per
via enzimatica a partire dal glucosio, a sua volta prodotto dalla fotosintesi clorofilliana
6 CO2 + 6 H2O + luce → C6H12O6 + 6 O2
n C6H12O6 + enzima → H-(C6H10O5)n-OH + n-1 H2O
La formazione dell'amido, cioè l'unione dell'amilosio e dell'amilopectina, è catalizzata da un enzima
chiamato amido sintetasi.
Utilizzi:
L'amido si trova nei frutti, nei semi e nei tuberi delle piante.
Nell'industria alimentare le cinque fonti principali di amido sono il mais, le patate, il riso, la tapioca e il
grano. Anche i legumi come i fagioli ne sono ricchi.
L'amido riveste particolare importanza nell'industria alimentare, quale agente addensante, e nella
produzione della carta, cartone e colle, in forma di salda d'amido, grazie alle sue proprietà collanti. In
questo caso l'amido viene utilizzato direttamente nell'impasto della cellulosa conferendo maggiore
resistenza meccanica alla carta che ne risulta. Altre applicazioni dell'amido riguardano l'industria
farmaceutica dove viene utilizzato come eccipiente.
L’ Amido
Proprietà In natura
l'amido ha disposizione semicristallina nei granuli, il che ne determina la quasi totale
insolubilità in acqua a temperatura ambiente; una parziale solubilizzazione è possibile
attraverso un aumento della temperatura, e porta alla formazione di un gel.
Il processo è influenzato dall'origine botanica dell'amido e dalla quantità di acqua. Un altro
effetto del processo di gelatinizzazione è la variazione della digeribilità dell'amido: la forma
cristallina è difficilmente attaccabile dagli enzimi (quali α- e β-amilasi), mentre quella
gelatinizzata lo è di più in conseguenza della disorganizzazione strutturale avvenuta.
AMIDO PRIMARIO
piccoli granuli di amido primario formato durante le fasi di intensa fotosintesi, quando questa
si attenua viene subito demolito; perciò non assumono mai grandi dimensioni né morfologia
particolare. Gli zuccheri che derivano dall’idrolisi dell’amido primario abbandonano il
plastidio verde e trovano altri impieghi nella stessa cellula o in cellule diverse.
Granuli di AMIDO SECONDARIO
I caratteri morfologici dei granuli di amido sono assai variabili
ILO: Centro di formazione del granulo di amido; attorno ad esso si osservano una
serie di strati concentrici più chiari e più scuri più o meno evidenti.
L’ilo può essere puntiforme (es. patata), lineare e ramificato (fagiolo), a stella
(mais)….
Nella maggior parte delle piante i granuli sono semplici, quindi indipendenti uno
dall’altro; in altre piante sono composti da un numero variabile di granuletti
elementari (avena, riso festuche, pepe….)
Granuli di amido secondario
Aspetto dei diversi granuli di amido
contenuti nei leucoplasti di diverse
piante coltivate
VACUOLI
VACUOLI
I vacuoli sono organuli
pieni di solvente acquoso
con disciolti prodotti
idrosolubili che li hanno
fatti apparire inizialmente
al microscopio elettronico
come
aree
vuote
(vacuoli).
Si tratta di organuli
delimitati da una singola
membrana (tonoplasto).
Cellula vegetale (mesofillo fogliare)
con vacuoli
Le sostanze raccolte nei vacuoli
In molte cellule vegetali si osservano vacuoli
di grandi dimensioni che occupano oltre il
90% del volume cellulare.
Uno dei principali problemi della cellula
vegetale è quello di stoccare una grande
quantità di sottoprodotti metabolici che
possono risultare dannosi se diffusi nel
citoplasma.
Mentre gli animali possiedono sistemi
escretori per eliminare tali sostanze, i vegetali
ne sono sprovvisti e utilizzano i vacuoli per
immagazzinare
le
sostanze
di rifiuto.
Queste
sostanze
possono
essere
immagazzinate anche in quanto tossiche per
gli erbivori e quindi costituire una protezione
per la pianta che risulta in questo modo assai
meno appetibile
Funzioni dei vacuoli
• elevate dimensioni mantenendo una
elevata superficie relativa;
• assorbire acqua dall’esterno e
mantenere il turgore cellulare;
• sostegno meccanico;
• deposito per sostanze superflue o
dannose;
• riserva (zuccheri, grassi e proteine);
• digestiva (enzimi);
• degradazione di macromolecole e
riciclaggio dei loro componenti;
• difesa (sostanze tossiche);
• Sito di accumulo dei pigmenti
(strategia riproduttiva - dispersione dei
semi).
In molte cellule vegetali si osservano vacuoli di
grandi dimensioni che occupano oltre il 90% del
volume cellulare.
IL SUCCO CELLULARE
COMPOSIZIONE:
- ACQUA (90%)
- ACIDI ORGANICI E LORO SALI
- GOMME
- PROTEINE
- OLII
- ALCALOIDI
- RESINE
- INCLUSI SOLIDI
- SALI INORGANICI
- ZUCCHERI
- MUCILLAGINI
- ENZIMI
- PIGMENTI
- TANNINI
- BALSAMI
Queste sostanze sono presenti in SOLUZIONE o sotto forma COLLOIDALE così da
determinare una forte vischiosità.
La composizione del succo cellulare è variabile a seconda della specie, dell’organo, del
tessuto, ecc.
IL SUCCO CELLULARE
GLUCOSIDI E ALCALOIDI
Sono le sostanze contenute nelle piante medicinali o in quelle velenose, nei vacuoli di diversi
organi della pianta: foglie, bulbi, rizomi, frutti e semi.
Gli alcaloidi sono sostanze azotate a carattere basico, poco solubili in acqua, ma capaci di
formare sali solubili in acidi sia minerali che organici quali: COLCHICINE (nel Colchicum
autunnale) e in numerose Dicotiledoni.
TANNINI
Sono diffusi soprattutto nei parenchimi delle cortecce, nel mesofillo delle foglie e nei frutti:
es. nella Quercia (9-12%); negli eucalipti (20-40%).
FUNZIONI:
-sostanze di riserva
-protezione contro l’attacco dei microrganismi.
RESINE
Sono frequenti soprattutto nelle Conifere.
CAUCCIU’
Sono politerpeni diffusi nei canali laticiferi di numerose famiglie (Moracee, Composite,
Euforbiacee).
IL SUCCO CELLULARE
SALI INORGANICI
Si tratta soprattutto di POTASSIO, CALCIO E MAGNESIO. Nel succo delle piante ALOFILE viene
accumulata una grande quantità di sale sotto forma di cloruro.
ACIDI ORGANICI
La loro presenza è la causa del sapore acido di numerose piante.
ACIDO MALICO (MELE)
ACIDO TARTARICO (UVA)
ACIDO CITRICO (LIMONE)
IDRATI DI CARBONIO
MONOSACCARIDI:
- Sorbosio (sorbe)
- Glucosio (uva, susine, ciliegie)
- Fruttosio (pesche)
- Saccarosio (barbabietola, canna da zucchero
POLISACCARIDI:
- Inulina (Composite, Dalia)
LIPIDI E OLII ETEREI
SI TROVANO:
- nei petali delle rose
- nelle radici del rafano senape, giaggiolo e
zenzero
- nelle cortecce: cannella
- negli involucri dei frutti o dei semi: pepe
INCLUSI SOLIDI NEI VACUOLI
GRANULI DI ALEURONE : riserve proteiche che solidificano sotto forma di concrezioni. Si
ritrovano soprattutto nei semi.
Nel ricino essi sono costituiti da una sostanza amorfa inclusa nella quale vi sono un
GLOBOIDE e un CRISTALLOIDE.
Il GLOBOIDE, caratterizzato da una maggiore insolubilità, è il primo a comparire, in seguito
alla disidratazione che accompagna la maturazione del seme. E’ formato di FITINA, SALE DI
Ca e Mg dell’ACIDO INOSITOLOESAFOSFORICO.
Il CRISTALLOIDE deriva dalla solidificazione delle globuline (proteine).
GLOBOIDE
CRISTALLOIDE
INCLUSI SOLIDI NEI VACUOLI
CRISTALLI DI OSSALATO DI CALCIO: Sono costituiti da un sale insolubile che può assumere
forme diverse:
DRUSA: cristallo bipiramidale nel quale, su ogni faccia, si depositano numerosi altri cristalli.
Es. tiglio
RAFIDI: Prismi monoclini allungati così
da assomigliare ad aghi appuntiti riuniti
in fascetti. Es. scilla, fitolacca
STILOIDI: Cristalli prismatici allungati.
Es. cipolla
SABBIA CRISTALLINA: Si ritrova nelle
Solanacee. es.belladonna
INCLUSI SOLIDI NEI VACUOLI
La formazione di cristalli di ossalato di calcio sembra essere collegata alla tossicità dell’acido
ossalico che si forma durante la respirazione: esso verrebbe fatto precipitare nei vacuoli per
renderlo inoffensivo.
La conoscenza delle forme
precipitazione dei cristalli
ossalato di calcio permette
riconoscimento delle piante
cui sono presenti.
di
di
il
in
PARETE
CELLULARE
Parete cellulare
Una parete che avvolge la cellula
esternamente è presente:
• nei procarioti
• nei funghi
• nei vegetali
Funzioni della parete
• Da sostegno e da forma alla cellula, ai tessuti e a tutta la pianta
(stretta relazione tra struttura della parete e funzione della
cellula);
• Controllo dell’espansione cellulare
• Controllo del trasporto intercellulare (presenza di enzimi);
• Protezione del protoplasto contro molti agenti esterni che
potrebbero danneggiarlo (siccità, parassiti, microrganismi
patogeni, ecc.) sia passiva (barriera) che attiva (invio di segnali
chimici in risposta all’invasione di un parassita che si propagano
alla pianta e stimolano meccanismi di difesa come produzione
antibiotici tossici x il patogeno o deposizione lignina che agisce da
barriera);
• Accumulo di sostanze di riserva.
PARETE E MEMBRANA
•La parete ha uno spessore che può arrivare a parecchi micron
•Lo spessore della membrana è circa 1000 volte più piccolo (nm)
•La parete è fatta in gran parte da polisaccaridi
•La membrana da lipidi complessi e proteine
•La parete ha doti di resistenza meccanica, ma vale poco come
barriera chimica (è scarsamente selettiva)
•La membrana non ha molta resistenza meccanica, ma è specializzata
come barriera chimica (è altamente selettiva). La resistenza al flusso
dell’acqua è per il 90% dovuto alla membrana cellulare.
ORIGINE DELLA PARETE CELLULARE
Quando due cellule si separano (mitosi) permane tra di esse un sottile
setto di separazione: PIASTRA C.
LAMELLA MEDIANA.
A questa si appone, sul lato interno una sottile parete
detta PARETE PRIMARIA. Accrescimento centripeto
Successivamente sempre internamente a questa si costruisce la PARETE
SECONDARIA.
LAMELLA MEDIANA
SOSTANZE PECTICHE
PARETE CELLULARE
CELLULOSA
Accrescimento della parete
La costruzione della parete avviene gradualmente, per stadi che corrispondono ad
altrettanti stadi di sviluppo della cellula.
Dato che i costituenti della parete sono sintetizzati dal citoplasma ne consegue
che la parete si accresce dall’esterno verso l’interno in direzione centripeta.
Man mano che la parete diventa più grossa, il lume
cellulare si restringe
Una cellula adulta conserva tutti gli strati di parete tipici della
cellula giovane; l’ultimo strato costruito è il più interno, cioè il più
vicino al citoplasma che ha prodotto tutta la parete.
Spessore della parete
Lo spessore della parete varia nei diversi tipi di cellule in rapporto alla
funzione del tessuto di cui fanno parte.
Lo spessore, quindi, dipende dal ruolo che le cellule giocano nella
struttura della pianta ed in parte dalla loro età.
Lo strato più giovane della parete è quello a contatto col protoplasto.
I primi strati formati costituiscono la PARETE PRIMARIA.
La regione che unisce pareti primarie di cellule differenti è la LAMELLA
MEDIANA
Molte cellule depositano strati aggiuntivi sulla superficie interna della
parete primaria che formano la PARETE SECONDARIA.
Spessore della parete
Le cellule nascono come cellule meristematiche (cellule non ancora
specializzate x una precisa funzione, la cui attività principale è quella
di dividersi frequentemente): esse hanno solo la lamella mediana e
un’esile parete primaria.
Nelle cellule adulte oltre alla parete primaria si ha anche la parete
secondaria.
Differenze tra parete e lamella
mediana
• spessore
• fisiologia
• composizione chimica:
la lamella mediana è costituita essenzialmente da
pectine
la parete primaria da pectine, cellulosa, emicellulose,
proteine
Lamella
mediana
La lamella mediana è comune a due
cellule contigue.
COMPOSIZIONE CHIMICA: costituita da
sostanze
pectiche
(le
pectine,
polisaccaridi non cellulosici), amorfe con
proprietà adesive tra le cellule e
gelificanti.
La lamella mediana è caratteristica dei
tessuti, tutte le cellule di un tessuto sono
unite dalla lamella mediana.
Le
pectine
Sono
polisaccaridi
fortemente idrofili con capacità di
gelificare, di conseguenza conferiscono
alla parete proprietà di elasticità e
flessibilità, permettendone quindi la
distensione
Finito il periodo delle divisioni una cellula passa attraverso due fasi
di sviluppo:
1. DISTENZIONE (le cellule non aumentano di numero ma di
dimensioni)
2. DIFFERENZIAZIONE (le cellule ormai grandi si specializzano per
una determinata funzione a seconda del tessuto cui
appartengono)
PARETE PRIMARIA
Tipica delle cellule in fase di distensione
Ha la capacità di crescere in superficie adattandosi all’aumento di dimensioni
della cellula.
E’ caratterizzata dalla presenza di cellulosa che manca nella lamella mediana
COMPOSIZIONE CHIMICA
• 20-30% cellulosa (che manca nella lamella mediana) in microfibrille a rete
• 25% emicellulosa del tipo degli xiloglucani
• 30% pectina
• Il resto da proteine del tipo dell’estensina
Cellulosa
• È la molecola organica più abbondante sulla terra essendo il
componente principale della parete delle cellule vegetali.
• In varie forme (carta, legno, materie plastiche cellulosiche, fibre
tessili), si trova alla base di numerose attività economiche umane.
• Non è assimilabile dalla maggior parte degli animali, tranne gli
erbivori ruminanti e alcuni gruppi di insetti (tra cui le termiti), che
dispongono degli enzimi adatti a degradarla in carboidrati semplici.
• La cellulosa è una molecola molto stabile difficilmente degradabile
A sinistra microfibrille di cellulosa con
struttura dispersa della parete
primaria
A destra microfibrille di cellulosa con
struttura orientata della parete
secondaria
Cellulosa
Componente principale della parete.
E’ un polisaccaride costituito da unità di glucosio legate tra loro
tramite legami β 1->4.
Le molecole di glucosio sono ruotate di 180° l’una rispetto all’altra per
consentire la formazione del legame; di conseguenza l’unità
fondamentale della cellulosa è un disaccaride detto cellobiosio.
Organizzazione delle fibrille
• Le microfibrille (costituite da numerose catene di cellulosa disposte su più piani)
sono stabilizzate da legami idrogeno intracatena e intercatena.
• Lungo le fibrille si trovano zone (dette micelle) con struttura altamente ordinata,
di tipo cristallino, alternate ad altre zone con aspetto meno organizzato.
• Grazie alla sua organizzazione, la cellulosa presenta straordinarie caratteristiche di
resistenza alla trazione ed agli attacchi enzimatici.
Organizzazione delle fibrille
Parete primaria
Parete secondaria
Macrofibrilla
Microfibrilla
Micella
Molecola di
cellulosa
(Cellobiosio)
Organizzazione delle fibrille
Emicellulose
• Le emicellulose costituiscono il 40-45% delle pareti primarie e il 30-33% delle
pareti secondarie.
• Sostanze estraibili dalla parete mediante trattamento con alcali.
• In base alla composizione chimica si tratta di catene di monosaccaridi, più o
meno ramificate.
• Polimeri di xiloso, xiloso e glucoso, glucoso e mannoso, galattoso e mannoso,
mannoso, glucoso, con vari tipi di legami (α o β, 1->3, 1->4, 1->6). Ramificazioni
ed eterogeneità impediscono l’impacchettamento in fibrille.
• Si tratta di polisaccaridi del tipo degli xiloglucani nelle dicotiledoni e xilani nelle
monocotiledoni.
• Queste emicellulose si legano alle microfibrille di cellulosa tramite legami
idrogeno.
• Hanno un ruolo fondamentale nella regolazione dell’aumento delle
dimensioni della cellula in quanto limitano l’estensibilità della parete tenendo
insieme microfibrille adiacenti.
• Frequentemente svolgono il ruolo di accumulo di sostanze di riserva oppure di
trattenimento dell’acqua.
Proteine
• Esistono proteine strutturali ed enzimatiche.
• Le prime contribuiscono alla formazione e all’organizzazione delle
strutture (Glicoproteine).
• Associate a queste si trovano componenti lipidici.
• Le seconde, dette Proteine enzimatiche, fungono da catalizzatori
per le reazioni biochimiche (i catalizzatori sono acceleratori di
reazioni chimiche).
• Nella parete sono presenti sia proteine strutturali, sia
enzimatiche.
Parete cellulare
riepilogo
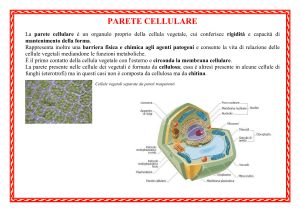
La parete cellulare è un organulo proprio della cellula vegetale, cui conferisce
rigidità e capacità di mantenimento della forma.
IN QUALI CELLULE
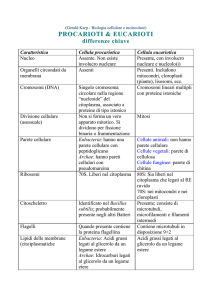
La parete presente nelle cellule dei vegetali è formata da cellulosa, negli Archaea e
nei Batteri è formata da peptidoglicano. Essa è altresì presente in alcune cellule di
funghi (eterotrofi) ma in questi casi non è composta da cellulosa ma da chitina. Gli
altri organismi eterotrofi (come gli animali) ne sono sprovvisti.
DOVE SI TROVA
È il primo contatto della cellula vegetale con l'esterno; aderisce alla lamella
mediana, circonda la membrana cellulare e delimita il protoplasto, termine che
indica la parte cellulare contenuta all'interno della parete.
CHE FUNZIONE SVOLGE
Ha funzione di regolazione verso la pressione osmotica qualora la cellula si trovi in
ambiente non isosmotico e per mantenere la pressione di turgore.
PARETE
SECONDARIA
Una volta raggiunte le dimensioni definitive, la cellula completa il
proprio differenziamento e inizia a funzionare come cellula adulta.
La maggior parte delle cellule del corpo della pianta possiede solo
una parete primaria. Alcuni tipi cellulari, però, completano il
differenziamento deponendo una parete secondaria all’interno
della parete primaria.
La parete secondaria è composta principalmente di fibre di
cellulosa con piccole quantità di emicellulose e pectine pertanto
mostra una flessibilità estremamente ridotta.
Formazione della parete secondaria
Terminata
la
distensione
cellulare
l’accrescimento in spessore prevale su quello
in superficie. L’accrescimento in spessore dà
origine alla parete secondaria.
Il contenuto di pectine e acqua cala
drasticamente.
Aumenta la cellulosa ed aumentano le zone
con struttura cristallina; inoltre i polimeri di
cellulosa sono più lunghi.
Microfibrille tendono ad associarsi e formare
macrofibrille.
Le variazioni di composizione dipendono dalla comparsa di nuove sostanze
(lignina, cere e cutine, suberina e sporopollenina) e dalla variazione di
percentuale di quelle precedentemente presenti.
Caratteristiche della parete
secondaria
La parete secondaria si forma dopo che la cellula ha cessato di
crescere e la parete primaria non si estende più in superficie.
Le pareti secondarie sono importanti nelle cellule specializzate nelle
funzioni di sostegno e di conduzione dell’acqua.
In queste cellule spesso il protoplasto muore dopo aver depositato la
parete secondaria.
E’ costituita per lo più di cellulosa (molto più abbondante della parete
primaria) e possono mancare le sostanze pectiche
La parete secondaria è molto rigida e poco flessibile.
Le microfibrille di cellulosa vengono apposte in lamelle contro la
parete primaria (in direzione centripeta).
La continua apposizione di lamelle dà origine a una disposizione in
strati concentrici, in generale vengono riconosciuti tre strati: S1, S2,
S3.
Sezione trasversale del legno di un fusto di dicotiledone arborea. Si nota la diversa
dimensione e struttura delle cellule che danno origine agli anelli annuali.
Connessioni cellulari
PLASMODESMI
La parete cellulare di una cellula vegetale ha una consistenza ed uno spessore che
garantiscono un adeguato sostegno alle cellule e all’intero organismo vegetale, ma
non permetterebbe alcuno scambio tra cellule contigue.
Questo è garantito da assottigliamenti della parete che consentono il transito di
molecole e soluti da una cellula all’altra attraverso la membrana cellulare.
PLASMODESMI
Le pareti primarie non sono uniformemente ispessite, ma presentano
aree meno ispessite chiamate aree di punteggiature primarie.
I plasmodesmi (canali citoplasmatici) che congiungono cellule
adiacenti sono riuniti generalmente nei campi di punteggiature
primarie.
Figure 34.8 Plasmodesmata (Part 1)
Punteggiature o porocanali
Il
deposito
della
parete
secondaria non avviene in
corrispondenza dei campi di
punteggiature primarie: si hanno
interruzioni
della
parete
secondaria dette punteggiature.
Campo di punteggiatura primaria (parete
primaria)
1 Punteggiatura (parete secondaria)
Le pareti secondarie
lignificate
sono
permeabili all’acqua
solo
in
corrispondenza delle
punteggiature.
La punteggiatura della parete di una cellula si trova
generalmente in corrispondenza della punteggiatura della
cellula con cui è in contatto.
La lamella mediana + le due pareti primarie tra le due punteggiature = membrana della
punteggiatura
Nelle cellule che hanno pareti secondarie si trovano 2
principali tipi di punteggiatura:
Semplice o Areolata (la parete secondaria si inarca sulla
cavità della punteggiatura)
PARETE COME RISERVA D’ACQUA
• Tutti i componenti di una parete normale sono idrofili, cioè
trattengono l’acqua ad eccezione delle pareti modificate in modo
speciale (es. cutinizzate o suberificate).
• La parete può essere immaginata come una spugna intersecata da
innumerevoli canali pieni d’acqua.
• Quest’acqua può attraversare la membrana ed entrare nella cellula
o può essere ceduta all’ambiente esterno (vapore)
• Le cellule di un tessuto sono direttamente o indirettamente unite
tra loro formando un labirinto senza continuità in cui l’acqua può
spostarsi liberamente trascinando con sé molti soluti.
• Questo spazio continuo occupato dalle pareti cellulari in cui
possono circolare liberamente acqua e soluti si chiama APOPLASTO
• L’acqua può percorrere una strada lunghissima prima di essere
assorbita da una cellula
Modificazioni secondarie della parete
• LIGNIFICAZIONE
• SUBERIFICAZIONE
• CUTINIZZAZIONE
• GELIFICAZIONE
• MINERALIZZAZIONE (silicizzazione e calcificazione)
• PIGMENTAZIONE
Esempi di
modificazioni
secondarie della
parete cellulare in
vari organi della
pianta
Lignificazione
foglie
fusto
Cutinizzazione
foglie
fusto
Suberificazione
fusto
LIGNINA
La lignina è un costituente delle pareti di molti tipi di cellule.
La lignina è un polimero organico costituito principalmente da
composti fenolici
Conferisce rigidità e resistenza alla compressione
Si trova nelle pareti delle cellule che hanno una funzione meccanica o
di sostegno
La lignina è idrofoba e si sostituisce all’acqua nella parete cellulare
Dopo la cellulosa la lignina è il polimero organico più abbondante nel
mondo vegetale.
LIGNINA
• La parete cellulare lignificata è idrofoba, resistente e non plastica.
• Principale funzione è quella di conferire forza di compressione e durezza alla
parete cellulare.
• La deposizione lignina ha avuto ruolo importante nell’evoluzione perché la
resistenza delle pareti lignificate ha permesso alle piante di aumentare la loro
altezza e lo sviluppo di ramificazioni imponenti.
• Infatti le pareti non lignificate possono resistere alla tensione, ma non alla
compressione (come quella esercitata dalla forza di gravità)
• Ha una funzione molto importante nel trasporto interno di acqua, nutrienti e
metaboliti.
• La connessione tra le diverse cellule del legno crea un materiale molto resistente
agli urti, alle compressioni e alle flessioni.
• I tessuti lignificati resistono molto bene agli attacchi dei microorganismi, non
permettendo la penetrazione di enzimi distruttivi nella parete cellulare.
LIGNINA
Sviluppi commerciali
• Le lignine sono fortemente ricercate per il legname da
riscaldamento, avendo un notevole potere calorifico. Al
contrario, non sono utilizzate nella fabbricazione della
carta, responsabili della sua colorazione giallastra dopo
l'esposizione al sole. Infine, essendo poco digeribili sono
poco ricercate nella coltura delle piante da foraggio.
• Per queste ragioni, sono in corso numerose ricerche
(essenzialmente genetiche), allo scopo di produrre dei
vegetali meno ricchi di lignina per la fabbricazione della
carta, o al contrario, con un tasso di lignina più elevato per
il legno da riscaldamento.
Castanea sativa
Castanea sativa frutti
“castagno”
Castanea sativa
Castanea sativa
Distribuzione italiana
del castagno
Areale di Castanea sativa
• naturale
° coltivato
Fagus sylvatica
Fagus sylvatica fiori
Fagus sylvatica frutti
“faggio”
Fagus sylvatica
Fagus sylvatica
Distribuzione italiana
del faggio
Areale del Fagus sylvatica
Quercus petraea “rovere”
Quercus petraea
Distribuzione della rovere
in Italia
Areale di Q. petraea
Quercus
robur
“farnia”
Quercus robur
Distribuzione della farnia
in Italia
Areale di Quercus robur
Quercus pubescens “roverella”
Quercus pubescens amenti maschili
Quercus pubescens frutti
Quercus pubescens
Distribusione italiana
della roverella
Areale di Quercus pubescens
Quercus cerris “cerro”
Quercus cerris frutti
Quercus cerris
Distribusione italiana
del cerro
Areale di Quercus cerris
Quercus ilex “Leccio”
Quercus ilex fiori maschili
Quercus ilex frutti
Quercus ilex
Distribusione italiana
del leccio
Areale di Quercus ilex
Suberina e suberificazione
• Impermeabilizza totalmente la cellula.
• Si tratta nuovamente di un poliestere, di acidi grassi, alcoli e
ossiacidi, con componenti fenoliche lignino simili.
• È tipica degli elementi esterni del fusto, ma può essere presente
anche in zone diverse della pianta.
• A differenza della cutina, le placche di suberina vengono deposte
all’interno della parete.
• La suberina è altamente idrofoba e funzione principale è quella di
protezione, che si esercita impedendo all'acqua di penetrare
attraverso i tegumenti esterni.
• Nelle radici la suberina è depositata nelle pareti radiali e trasversali
delle cellule endodermiche. Questa struttura è conosciuta come la
fascia del Caspary.
• Funzione di questa struttura è impedire che l'acqua e le sostanze
nutrienti assorbite dalla radice entrino nel sistema di trasporto
interno senza che la pianta effettui una selezione dei soluti.
IL SUGHERO
E’ il costituente principale
della corteccia di molti alberi.
Nella quercia da sughero (Quercus
suber) si accresce sensibilmente
dando
origine Sughero grezzo
a strati di parecchi cm di spessore.
Sughereta
Dal punto di vista istologico il
sughero è formato da cellule
morte di forma geometrica
disposte in serie regolari e
praticamente impermeabili all’aria
e all’acqua.
Caratteristiche del sughero sono
la
leggerezza,
l’assoluta
impermeabilità ai liquidi e ai gas,
l’ottima coibentazione, l’elasticità
e la resistenza ai parassiti.
Scorzatura del sughero
Prodotti lavorati
Quercus suber “sughera”
Quercus suber
Distribusione italiana della sughera
Areale di Quercus suber
Cutina suberina e cere
CUTINA SUBERINA e CERE sono particolari lipidi che formano barriere contro la
perdita d’acqua.
*Presenza di CUTINA
*impregnazione delle pareti (CUTINIZZAZIONE)
*strato superficiale (CUTICOLA).
*La cutina non è presente nelle pareti epidermiche delle piante sommerse, di luoghi
umidi, degli organi sotterranei (radice).
*La suberina è il principale componente delle pareti del sughero, cioè di quelle
cellule che formano lo strato più esterno della corteccia.
*Le cere sono composti lipidici a lunga catena; sono i composti dei lipidi più
idrorepellenti. Insieme a cutina e suberina formano delle vere e proprie barriere
capaci di impedire la perdita d’acqua e di altre molecole dalle superfici vegetali.
Cutine e cutinizzazione
Strato di cutina
• La parete esterna delle cellule superficiali
(cellule
epidermiche)
subisce
una
modificazione chimica secondaria della sua
composizione, mediante immissione di cutina
e cere nella matrice. Cutina e cere sono lipidi
che formano barriere contro la perdita
d’acqua. CUTINIZZAZIONE.
• La cutina spesso viene escreta all’esterno della
parete a formare uno strato più o meno
consistente di cutina allo stato puro (non
mescolata, cioè, agli altri componenti della
parete) denominato “CUTICOLA” e , in certe
piante, può essere ricoperta da un ulteriore
strato impermeabilizzante costituito da cere.
CUTICOLA
• Quasi totalmente impermeabile a acqua e gas: protezione contro
perdita d’acqua e contro parassiti.
• La cuticola da irrobustimento meccanico all’epidermide
• Lo spessore varia con la necessità di protezione delle piante (è
maggiore nelle piante di luoghi aridi)
• Si trova sull’epidermide di foglie, fusti, fiori, nettari, peli; non è
presente nella radice.
• La cuticola è costituita da cere (catene di idrocarburi a lunga catena
ed esteri di acidi grassi) e cutine (poliestere a maglia
tridimensionale di ossiacidi, acidi grassi e acidi epossidici).
* A volte le cere sono in granuli che formano sull’epidermide una
pruina asportabile (prugne, uva, ecc.)
Mineralizzazione
• Avviene per deposizione
uniforme di sali minerali tra
i vari componenti della
parete.
• E’ caratteristica dei tessuti
dei tegumenti, dove
assume funzione di
protezione (silicizzazione,
calcificazione).
Pareti silicizzate di
equiseto
Pareti silicizzate su foglie di tagliamani
(Ampelodesmos mauritanica)
• Comune nelle foglie delle
graminacee (margine
tagliente) e negli equiseti
(che provocano
microlesioni negli animali
che se ne cibano).
Peli urticanti di ortica
(Urtica urens)
Pareti incrostate di calcare su sassifraga
Gelificazione
• Avviene quando le sostanze pectiche (gomme,
mucillagini) prevalgono sulle altre componenti
che costituiscono l’intelaiatura cellulosica,
accumulandosi all’esterno.
• Come conseguenza della gelificazione si ha
l’acquisizione da parte della parete di una grande
capacità idrica. Fenomeno che si verifica nei
tegumenti dei semi di alcune piante (lino, ecc.)
che si rigonfiano al momento della germinazione
costituendo una notevole riserva d’acqua per la
plantula.
• Il fenomeno interessa anche molte piante
inferiori, alghe azzurre, alghe rosse (agar-agar).
• Processo che avviene anche in seguito a ferite o
danni all’epidermide in seguito alle quali si
formano essudati (gomme vegetali), come nel
ciliegio o la gomma arabica delle acacie.