caricato da
lucio0091
Trasporto attivo, osmosi e stomi: Appunti di biologia

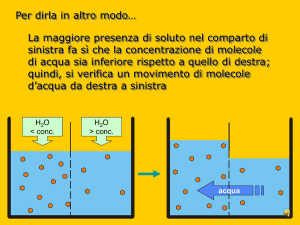
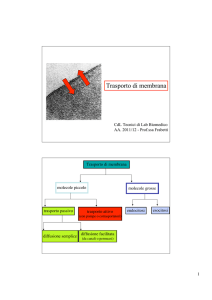
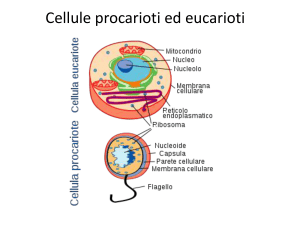
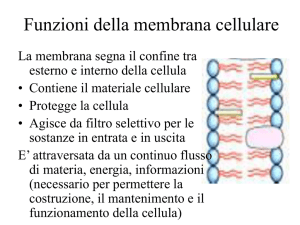
Transporto attivo La catena di trasporto degli elettroni rappresenta la prima fase della fosforilazione ossidativa. In questa fase, gli elettroni dei coenzimi ridotti NADH e FADH2, provenienti dall'ossidazione dei composti organici, vengono ceduti alla catena di trasporto degli elettroni per creare un gradiente protonico (ioni H + ) nello spazio intermembrana del mitocondrio. Come avviene la catena di trasporto degli elettroni La catena di trasporto degli elettroni è un sistema proteico legato alla membrana mitocondriale interna, la quale separa la matrice mitocondriale dallo spazio intermembrana. E' formata da 4 complessi proteici transmembrana e da 2 trasportatori solubili. Il suo compito è quello di accettare gli elettroni ad alta energia provenienti dai coenzimi ridotti NADH e FADH2 e di trasferirli attraverso la catena tramite reazioni di ossido-riduzione a potenziale via via crescente. Ad ogni reazione di ossido-riduzione parte dell'energia degli elettroni viene ceduta al sistema, permettendo di attivare il pompaggio dei protoni nello spazio intermembrana contro il loro gradiente. L'accettore finale degli elettroni è l'ossigeno, che catturando 2 elettroni e 2 protoni viene ridotto ad acqua (H2O). Schema del percorso degli elettroni Complesso I. NADH-Ubichinone-Ossidoreduttasi. Questa proteina di membrana catalizza il trasferimento di 2 elettroni dal coenzima ridotto NADH al trasportatore solubile Ubichinone (o Coenzima Q). La reazione avviene attraverso diversi passaggi. Inizialmente il NADH viene ossidato a NAD + , riducendo il Flavim Mononucleotide a FMNH2 con il trasferimento simultaneo dei 2 elettroni. Dopodiché, ciascun elettrone viene trasferito singolarmente dapprima passando per diversi centri ferro-zolfo e successivamente viene donato all'Ubichinone, che al primo ciclo forma un radicale semichinonico e al secondo ciclo viene completamente ridotto ad Ubichinolo (QH2). Durante il processo vengono trasferiti 4 protoni nello spazio transmembrana. L'esatto meccanismo con cui i protoni vengono traslocati è ad oggi ancora da chiarire. Complesso II. Succinato-CoQ-Reduttasi. La reazione del Complesso II è una via alternativa del Complesso I e catalizza anch'essa il trasferimento di elettroni al Coenzima Q. Il Complesso II è di fatto l'enzima che catalizza la sesta reazione del Ciclo di Krebs trasformando il Succinato in Fumarato, ed è l'unico enzima del Ciclo di Krebs che si trova legato ad una membrana. Durante la reazione viene ridotta una molecola di FADH2, la quale cede gli elettroni ai centri ferro-zolfo del Complesso II ed infine all'Ubichinone. Diversamente dal Complesso I, in questo processo non vengono trasferiti protoni. Complesso III. CoQH2-CitocromoC-Ossidoreduttasi. L'ossidazione dell'Ubichinolo da parte del Complesso III avviene in modo ciclico in un processo chiamato Ciclo Q. I due elettroni trasportati dal Coenzima Q vengono strappati uno alla volta, generando un intermedio semichinonico, e trasferiti sui centri ferro-zolfo presenti sul Complesso III, dove vengono pompati 4 protoni nello spazio intermembrana (ovvero 2 protoni per ogni ciclo di 1 elettrone). Infine, ogni elettrone viene trasferito su una molecola del trasportatore solubile Citocromo C. Complesso IV. CitocromoC-Ossidasi. Qui si svolge l'ultima tappa del lungo percorso degli elettroni. 4 molecole di Citocromo C vengono ossidate dal Complesso IV passando per dei centri contenenti atomi di rame (Cu). Gli elettroni vengono infine donati all'ossigeno molecolare (O2), che funge da accettore finale. I due atomi di ossigeno che compongono la molecola vengono scissi e ridotti dai 4 elettroni, e combinandosi con 4 protoni prelevati dalla matrice mitocondriale si trasformano in 2 molecole di acqua (H2O). Durante il processo vengono inoltre pompati altri 4 protoni nello spazio intermembrana. Il movimento dell’acqua può avvenire per: •DIFFUSIONE (gradiente di concentrazione) E’ dovuta al moto termico casuale delle molecole e si verifica quando si trovano a contatto due soluzioni a differente concentrazione: le molecole si muovono seguendo il gradiente di concentrazione, cioè da zone a più alta concentrazione a zone a più bassa concentrazione. Tale processo è efficiente sulle brevi distanze ed è molto importante per il movimento dell’acqua a livello cellulare •FLUSSO DI MASSA (gradiente di pressione) Il movimento di gruppi di molecole in risposta a gradienti di pressione. È un movimento globale delle molecole di un liquido che si spostano tutte insieme in una direzione e serve in genere per spostare l’acqua ed i soluti da una parte all’altra di un sistema pluricellulare (es xilema e floema) •OSMOSI (gradiente di potenziale idrico) Il termine osmosi indica in chimica e in fisica il fenomeno consistente nel movimento di diffusione di due liquidi miscibili di diversa concentrazione, attraverso un setto poroso o una membrana, semipermeabile . Ogni soluzione possiede una pressione osmotica che è direttamente proporzionale alla sua molalità. Quando sui due lati della membrana si trovano soluzioni a diversa concentrazione, la differenza di pressione osmotica muove le molecole di solvente (le molecule dell’acqua)dalla soluzione più diluita verso la soluzione più concentrata, fino a quando le concentrazioni delle due soluzioni diventano identiche. L’osmosi avviene in risposta ad una forza motrice. Alla forza motrice per il movimento contribuiscono sia il gradiente di concentrazione sia il gradiente di pressione che determinano la direzione e la velocità del flusso. La forza motrice è espressa come il Gradiente del Potenziale Chimico o più comunemente dai fisiologi vegetali come il GRADIENTE DI POTENZIALE IDRICO le differenze tra Osmosi e diffusione La diffusione può avvenire in qualsiasi miscela, inclusa quella che include una membrana semipermeabile, mentre l'osmosi si verifica sempre attraverso una membrana semipermeabile.Quando le persone discutono di osmosi in biologia, si riferisce sempre al movimento dell'acqua. In chimica, è possibile che vengano coinvolti altri solventi. In biologia, questa è una differenza tra i due processi.Una grande differenza tra osmosi e diffusione è che sia le particelle di solvente che quelle di soluto sono libere di muoversi in diffusione, ma quando parliamo di osmosi, solo le molecole di solvente (molecole d'acqua) attraversano la membrana. Assorbimento e conduzione Attraverso i peli radicali la radice assorbe l'acqua dal terreno per osmosi e i sali minerali per trasporto attivo. Le soluzioni fluiscono per diffusione attraverso l'epidermide e la corteccia, quindi giungono all'endoderma dove cellule ispessite impermeabili le costringono a passare attraverso il citoplasma e a entrare nei vasi legnosi: questi, paragonabili a capillari, fanno salire le molecole di acqua, tenute insieme in lunghe catene dalla forza di coesione, per una certa altezza. Contemporaneamente l'evaporazione dell'acqua dalle foglie crea una pressione negativa all'interno dei tessuti legnosi e una forza di suzione che richiama acqua dai vasi sottostanti: si forma così una colonna continua di acqua dalla radice alle foglie, che, regolando la traspirazione mediante gli stomi, regolano anche l'assorbimento a livello radicale. 1)osmosi: fenomeno chimico-fisico di diffusione tra due liquidi miscibili attraverso una membrana di separazione. Se due soluzioni a concentrazione diversa sono separate da una membrana semipermeabile (che lascia passare preferenzialmente le molecole di solvente, in genere acqua, mentre blocca quelle di soluto), si stabilisce un flusso spontaneo di solvente dalla soluzione più diluita verso la soluzione più concentrata, fino a raggiungere un equilibrio in corrispondenza del quale il flusso di solvente è uguale nei due sensi della membrana. 2)trasporto attivo: trasferimento di sostanze (soluti) da un lato all'altro della membrana cellulare; avviene con dispendio di energia, sotto forma di consumo di molecole di ATP. Si realizza grazie a particolari proteine di membrana, specifiche per il trasporto delle diverse classi di sostanze, quali ioni, amminoacidi, zuccheri, ecc. Ognuna di queste proteine è in grado di riconoscere un particolare soluto e di legarsi a esso; utilizzando poi l'energia delle molecole di ATP, modifica la propria conformazione spaziale ruotando in modo tale che la molecola da trasferire, superando la barriera costituita dai fosfolipidi, venga a trovarsi sul lato della membrana opposto a quello in cui si trovava. Il trasporto attivo ha grande importanza poiché permette l'ingresso nella cellula o l'espulsione da essa di ioni e molecole contro gradiente, ossia da zone dove tali sostanze hanno una bassa concentrazione verso zone dove sono più concentrate, opponendosi pertanto al loro spontaneo movimento. Apertura degli stomi Di notte, quando gli stomi sono chiusi (ad eccezione delle CAM), le cellule di guardi sono afflosciate e presentano una bassa pressione interna. L’acqua piò facilmente entrare o uscire da esse. Le cellule di guardai sono in equilibrio idrico con le cellule annesse, per cui l’acqua entra e esce dalle prime con circa lo stesso tasso. Le cellule di guardia inoltre, contrariamente a tutte le altre cellule epidermiche, contengono i cloroplasti e dunque non sono trasparenti. la luce blu (ovvero quella delle prime ore dell’alba) che incide sulle cellule di guardia ha l’effetto diretto sull’apertura degli gli stomi dopo la chiusira notturna: è coinvolto un pigmento, la zeaxantina (famiglia xantofille), che agisce insieme a fotorecettori che percepiscono la luce blu. La luce blu induce l’attivazione di pompe protoniche H+-ATPasi, la sintesi di acido malico e l’idrolisi dell’amido. La zeaxantina regola l’apertura stomatica attivando la pompa protonica ATPasi mediante fosforilazione dei residui di Serine e Threonine del dominio regolatorio della poma protonica (tramite ser/tre chinasi) alla quale si lega anche la proteina 14-3-3! pompaggio all’esterno di H+ stabilendo una gradiente chimico con conseguente iperpolarizzazione della cellula. Questa iperpolarizzazione porta all’apertura dei canali e richiamo all’interno della cellula di ioni K+ attraverso i canali ionici. si tratta di canali di antiporto K+ /H+, e allo stesso tempo anche canali di simporto cloururo-H+. Allo stesso tempo, il cloroplasto produce sia saccarosio che malato; il saccarosio entra nel vacuolo, il malato anche, andando a bilanciare le cariche positive in entrata nel vacuolo. accumulando ioni potassio, rendono l’ambiente interno della cellula ipertonico rispetto all’esterno e provocano il passaggio dell’acqua nella cellula secondo un meccanismo di osmosi. I vacuoli della cellula si riempiono e la rendono turgida: di conseguenza, l’apertura dello stoma aumenta. Questi ioni vengono prelevati dalle cellule annesse, che si trovano adiacenti a quelle di guardia, e che vengono ceduti sotto particolari condizioni e stimoli (ad es.ormonali); insieme agli ioni potassio, carichi positivamente, la cellula, dovendo riequilibrare le cariche all’interno, sarà permeabile anche a sostanze cariche negativamente, come il malato, e ioni cloruro. E’ importante che le cellule di guardia siano in grado di svolgere la fotosintesi attraverso i cloroplasti e la respirazione attraverso i mitocondri per la produzione di ATP, necessaria per l’uscita degli H+. Quando ad esempio una pianta ha poca acqua a disposizione, non si può permettere di perderne, e gli stomi si chiudono. Questa risposta è mediata da un fito-ormone prodotto nelle radici, l’acido abscissico (ABA), che agisce principalmente sulle pompe ioniche delle cellule di guardia, le quali perdono così soluti e acqua, sgonfiandosi. Trasporto passivo Ioni e molecole apolari di piccole dimensioni passano la MP mediante diffusione non è richiesto l'utilizzo di energia biochimica (come ATP). Esso può essere mediato da proteine di membrana Questo tipo di trasporto può essere di due tipi differenti: diffusione semplice, diffusione facilitata avvengono secondo gradiente di concentrazione Diffusione semplice Non è coinvolta nessuna proteine di membrana. Il flusso netto di molecole è sempre a favore di gradiente: da un compartimento ad alta concentrazione a uno a bassa concentrazione Non c'è pertanto una spesa energetica in termini di ATP per spingere la molecola dall'altra parte della membrana. Diffusione facilitata La diffusione facilitata ha, come integrali della membrana, delle proteine canale o delle proteine carrier che facilitano e velocizzano il passaggio di una specifica molecola o di uno specifico ione, attraverso la formazione di legami deboli, necessari per facilitare il rilascio della molecola dall'altra parte della membrana. Osmosi L'osmosi rappresenta un tipo particolare di diffusione semplice in cui sono le molecole di un solvente (e non quelle del soluto) a permeare la membrana attraverso proteine-canale. Quando una membrana non fa passare le molecole di soluto ma solo quelle di solvente (che nel caso delle membrane biologiche è acqua) allora la membrana si dice semipermeabile per quel soluto specifico. La membrana plasmatica, ad esempio, è semipermeabile per molti soluti. Quando esiste un gradiente di concentrazione ai lati di una membrana semipermeabile le molecole di soluto non possono diffondere dall'area a concentrazione maggiore a quella a concentrazione minore. Si assiste, dunque, al fenomeno dell'osmosi, cioè al passaggio delle molecole di solvente dall'area a minor concentrazione di soluto a quella a maggior concentrazione di soluto, in modo da diluire la soluzione più concentrata e annullare il gradiente di concentrazione. In generale, quindi l'osmosi riguarda soprattutto il trasporto facilitato delle molecole d'acqua attraverso proteine-canale, che nella fattispecie sono dette acquaporine (notare che le molecole d'acqua sono polari, ma piccole quantità attraversano lo strato idrofobico della membrana). Trasporto attivo Il trasporto attivo permette il passaggio di soluti contro gradiente di concentrazione. Tale passaggio è mediato da proteine di membrana che richiedono l'utilizzo di energia biochimica (ATP). Questo tipo di trasporto può essere ulteriormente suddiviso in diverse tipologie: trasporto attivo primario e trasporto attivo secondario. NB* REAZIONI CHIMICHE Processo Esoergonico ed Endoergonico ESOERGONICO: processo che presenta, alla fine, un'energia potenziale minore di quella iniziale (processo che libera energia) ENDOERGONICO: processo che presenta alla fine un'energia potenziale maggiore di quella iniziale (processo che richiede energia) La maggior parte delle reazioni chimiche richiede un apporto iniziale di energia per poter prendere l'avvio. Questo vale sia per le reazioni endoergoniche che per quelle esoergoniche. Trasporto attivo primario Il trasporto attivo primario è mediato da proteine di membrana trasportatrici di soluti. Si occupa di trasferire molecole contro un gradiente di concentrazione, richiedendo la presenza di ATP (si verifica quindi una spesa energetica). Il trasporto attivo primario è in grado di generare un gradiente di concentrazione ed un gradiente elettrico a cavallo della membrana cellulare (come avviene, ad esempio, quando vengono traspostati degli ioni). Lo spostamento di un soluto dall'altra parte della membrana, ad opera della proteina di membrana, è strettamente legato alla reazione esoergonica della conversione dell'ATP in ADP e fosfato inorganico.. Trasporto attivo secondario Si parla di trasporto attivo secondario quando il trasporto di una molecola (A) contro gradiente è permesso dal gradiente di un'altra molecola (B) precedentemente creato da un trasporto attivo primario. Infatti è necessario che si verifichi prima il trasporto endoergonico (che cioè richiede energia) di B, affinché si accumuli da un lato della membrana. Successivamente si verifica il trasporto esoergonico di B nella direzione del gradiente, creato precedentemente dal trasporto attivo primario. Tale trasporto esoergonico è solitamente accoppiato al trasporto contro gradiente di A. Tale accoppiamento è definito cotrasporto ed è realizzato da proteine intrinseche della membrana cellulare capaci di riconoscere le molecole A e B. Solitamente B è uno ione, sodio o protone, che viene chiamato ione trainante (driver). Esistono tre tipi di cotrasporto: uniporto, antiporto e simporto. 1. L'uniporto, consente il passaggio di un solo ione o molecola in un'unica direzione. 2. L'antiporto, consente il passaggio contemporaneo ma in direzioni opposte di due ioni e/o molecole differenti. 3. Il simporto, consente il passaggio contemporaneo ma nella stessa direzione di due ioni e/o molecole differenti. Tra i trasporti secondari figurano: Na+/glucosio simporto; Na+/amminoacidi simporto; Na+/Ca2+ antiporto; Na+/H+ antiporto.