caricato da
giorgia91indellicati
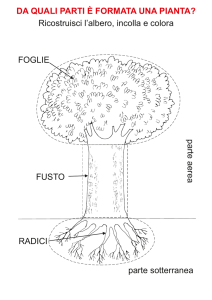
Biologia Vegetale: Cellula, Parete Cellulare, Plastidi

BIOLOGIA VEGETALE Procarioti→No nucleo, cell più semplici: Archea, Bacteria Eucarioti→Nucleo _animali _piante _funghi _protisti CELLULA VEGETALE La forma è determinata dalla parete cell., (cell deprivata di parete cell→protoplasto). Citosol→ soluzione colloidale contenuta nella m.p., costituita di acqua, ioni, lipidi, proteine enzimatiche e strutturali, zuccheri, nucleotidi, ormoni, RNA, ribosomi ede elementi del citoscheletro, al suo interno si svolgono reazioni e processi metabolici. Citoplasma→insieme del citosol e degli organuli (esclusi nucleo e vacuoli). Organuli caratteristici della cell veg: .plastidi→conferiscono il colore verde .vacuolo→zona priva di organuli e colore, nella cell adulta occupa gran parte del volume totale. Membrana plasmatica o plasmalemma: Costituita da doppio strato lipidico, Circonda la cell, costituisce l’interfaccia con l’ambientre extracell, è composta da lipidi, proteine e carboidrati, interagisce con la parete cellulare. Osmosi: Trasporto per diffusione semplice. Fenomeno che si manifesta quando una membrana semipermeabile (selettivamente permeabile) separa due soluzioni a diversa concentrazione, solo le molecole d’acqua possono diffondere, mentre i soluti no. Acqua da soluzione più diluita (+ acqua)→a soluzione più concentrata (+ soluti); il flusso dell’acqua verso la soluzione più concentrata produce una pressione detta pressione osmotica. Nella cell vegetale la parete cellulare rappresenta un sistema che si contrappone alla pressione osmotica, impedendo alla cell di espandersi fino a scoppiare (lisi cellulare), questo fenomeno è detto pressione di turgore. Il citoplasma è ricco di Sali, zuccheri ecc, di conseguenza se la cell viene immersa in soluzione ipotonica l’acqua passa attraverso la membrana dall’esterno all’interno della cell; se invece la cell viene posta in soluzione ipertonica l’acqua dall’interno della cell passerà all’esterno. Se la soluzione è isotonica (concentrazione di soluti uguale interno-esterni) non avviene osmosi. Mitocondri: Organuli ad attività metabolica: consumo cell di ossigeno e conservazione di energia (ATP). Forma variabile, numero variabile in fuzione della tipologia di cell e dello stadio di sviluppo di questa. Doppia membrana: esterna e interna, separate da spazio intermembrana. M. interna→enzimi per metabolismo e respirazione cell, forma le creste (per aumentare superfice cell e quindi efficienza). Matrice→ambiente interno, conprende ribosomi ed enzimi. Perossisomi: organuli con membrana singola (non doppia), contengono enzimi, sono compartimenti metabolici con funzione ossidativa, degradano molecole dannose con lo scopo di prevenire l’intossicazione cellulare. 1 Ribosomi: Sede della sintesi proteica, formati da 3 molecle di rRNA e proteine che si associano a formare le 2 subunità. Cell veg 3 tipi di ribosomi: citoplasmatici, mitocondriali e plastidiali. PARETE CELLULARE Struttura esterna alla membrana plasmatica, tipica delle cellule vegetali (ma anche nelle cellule di funghi e batteri) con spessore da 0,1 a 100 μm (=micrometri), che aumenta a livello degli angoli della cell. Conferisce forma alla cellula, da cui dipende la forma della pianta. E’ un compartimento utile nella conduzione di acqua e soluti e nella comunicazione cellulare, nel riconosciento self (cell stessa pianta) e non-self (es: patogeni). La parete varia a seconda di molto fattori come tipologia della cell, età, condizioni ambientali. Architettura parete cell: .Lamella mediana→in tutte le cell, porzione più esterna, sottile, responsabile dell’adesione cell-cell. Nelle cell con parete secondaria viene incrostata da lignina e scompare. .Parete primaria →in tutte le cell, è plastica, contiene il protoplasto, solitamente sottile, più spessa in alcuni punti. .Parete secondaria→solo in alcune cell specializzate, è formata da 3 strati apposti all’interno della parete primaria: esterno, mediano, interno, ricchi di cellulosa, spesso contengono lignina, struttura lamellare resistente, frequente la lignificazione. Costituenti della parete cellulare: .Polisaccaridi (polimeri di zuccheri) .Proteine strutturali ed enzimi .Fenilpropani (lignina=polimero di natura fenolica, idrofobico, rigido, poco comprimibile; conferisce alla parete idrofobicità e compattezza) .Componente fibrillare→cellulosa. Le singole unità di glucosio nella sua forma piranosica (cioè formante un anello composto da 6 unità di cui 5C e 1O), formano catene lunghe, il cellobiosio è l’unità di base che forma la cellulosa. Cellulosa: .è un polisaccaride strutturale costituito da dimeri di cellobiosio→è costituito da due molecole di β-Dglucosio unite da legami 1-4. L’unità di base, il cellobiosio, si ripete. .le catene si riuniscono in fibrille tenute insieme da legami H .le fibrille si attorcigliano in fasci, per formare le fibre presenti nella parete cellulare, queste avranno orientamenti diversi rispetto alla parete cellulare e al tipo di ispessimento per conferire resistenza. Dalle fibrille che compongono la parete cellulare dipende la forma della cellula: disposizione ordinata orizzontale→parallelepipedo; disposizione disordinata→sfere .la cellulosa è sintetizzata (polimerizzata) dalla pianta. _Componente matriciale (ovvero ciò che compone la matrice dove sono immerse le microfibrille di cellulosa): le microfibrille di cellulosa sono immerse in una matrice ricca d’acqua e costituita da polisaccaridi (emicellulose, pectine) e proteine strutturali in minor quantità. *(pectine→componente strutturale della parete cell, sono in grado di formare gel sia all’interno della p.c. che dopo essere state estratte, per questa caratteristica sono ampiamente utilizzate nell’industria alimentare e cosmetica. Emicellulosa→polisaccaridi eterogenei legati alla componente fibrillare da legami H, ricoprono microfibrille e connettono le fibrille formando una rete). 2 >Legno: tessuto morto, conduttore che porta la linfa grezza (radici→foglie). Ha cellule con parete cellulare molto spessa che formano canali per il passaggio dell’acqua. Specializzazioni e modificazioni della parete secondaria: La parete cellulare, durante la vita della cellula, assume spesso nuove caratteristiche chimico-fisiche, in stretto rapporto con le funzioni che deve svolgere.Le principali modificazioni della parete avvengono per: INCROSTAZIONE→cioè infiltrazione di materiali tra gli spazi interfibrillari delle molecole di cellulosa, 1)Lignificazione:alla fine della crescita e differenziamento cell nella parete può essere deposta lignina; sono lignificate le pareti secondarie di cell specializzate nella conduzione di acqua e nel sostegno della pianta (trachee). La lignina sostituisce la maggior parte dell’acqau in parete, questo fa diventare l’ambiente idrofobico e porta la cell alla morte programmata. La lignina è deposta anche come difesa in quanto è difficilmente degradabile dai parassiti e dagli erbivori. 2)Pigmentazione: pigmenti che conferiscono resistenza 3)Mineralizzazione: in parete vengono deposte sostanze minerali, es silice-silicizzazione→tipica dell’equiseto, lo rende ruvido, o carbonato di calcio-calcificazione→nei peli urticanti dell’ortica, li rende rigidi in modo che possano penetrare la cute quando la pianta viene toccata. APPOSIZIONE→sulla parete di materiali che ne aumentano l’impermeabilizzazione 1) Cutinizzazione: la parte aerea delle piante (Es: erbacee) viene rivestita da cutina, macromolecola che forma una rete lassa nella quale vanno ad inserirsi le cere. Cutina e cere insieme formano la cuticola che è impermeabile. 2) Cerificazione: le cere inserite nelle maglie della cutina creano cuticola che provoca impermeabilizzazione per limitare perdita di acqua e impedire il ristagno di acqua sulla superficie della pianta. 3) Suberificazone: nella parte sotterranea (radice) e a livello del fusto della pianta l’impermeabilizzazione viene attuata dalla deposizione di suberina, polimero che aderisce alla parete cellulare. La suberina conferisce anche resistenza contro gli agenti patogeni. GELIFICAZIONE→le pectine sono in grado di legare molta acqua e formare gel, facendo diventare alcune cell mucillagginose, questo si verifica fisiologicamente ad es nella radice (cellule della cuffia) per far si che l’attrito tra radice e terreno venga ridotto e la radice non venga daneggiata. La gelificazione può essere anche un meccanismo di difesa per suturare ferite al fusto in alcune piante. Crescita della parete: La cell cresce per distensione→ la parete deve “rilassarsi”, quindi i polimeri di parete devono scivolare l’uno sull’altro per permettere l’espansione, questo avviene in modo controllato grazie a modifiche chimiche della parete e alla presenza di enzimi specializzati. La distensione avviene anche per entrata di acqua nel vacuolo. L’orientamento delle microfibrille influenza la direzione di crescita che può avvenire ugualmente in tutte le direzioni oppure di più in lunghezza. Funzioni della parete cellulare: _Agisce da esocheletro e mantiene o determina la forma della cellula e dell’intera pianta attraverso l’adesione tra le cellule (lamella mediana) _Funzione meccanica (sopporta pressioni e trazioni, consente la crescita) _Controbilancia la pressione di turgore _Trasporto intercellulare dell’acqua (parete cell: porosa, carica, idrofilica) 3 _Protezione (barriera) _Produzione di molecole segnale (frammenti di parete rilasciati a causa della presenza di un patogeno, lo segnalano alla cell e alla pianta) _Accumulo di sostanze di riserva (usate poi durante la germinazione del seme) Parete cellulare e impieghi commerciali: • Carta, tessuti, fibre (cotone, lino, canapa ecc.) • Carbone, legna e derivati del legno • Altri derivati (estratti polisaccaridici modificati, per la produzione di plastiche, pellicole, rivestimenti, gel, addensanti) • Riserva naturale di carbonio organico (ciclo del carbonio negli ecosistemi) • Fonte di fibra nell’alimentazione umana e animale PLASTIDI Organuli con funzione metabolica, fotosintetica, biosintetica per acidi grassi e amminoacidi. I plastidi sono costituiti da un involucro fatto da due membrane: la MEMBRANA ESTERNA e la MEMBRANA INTERNA, il compartimento tra le due è detto spazio intermembrana. All’interno delle membrane si trova lo STROMA o MATRICE, fluido contenente vari metaboliti ed intermedi di reazione, ioni, proteine, ribosomi, DNA, lipidi. Nelle cell non ancora differenziate si trovano i PROPLASTIDI, precursori dei plastidi, piccoli e con sistema membranoso poco sviluppato. Il differenziamento dei proplastidi nelle varie forme di plastidi (cloroplasti, leucoplasti, cromoplasti) dipende da fattori ambientali (luce, temperatura) e da meccanismi di regloazione programmati. Dai proplastidi si differenziano quindi i plastidi, che si dividono a seconda di funzione, colore, sviluppo delle membrane interne e contenuto dello stroma, in tre diversi tipi: .CLOROPLASTI .LEUCOPLASTI .CROMOPLASTI Nel corso dello sviluppo della pianta sono frequenti interconversioni di un tipo plastidiale in un altro. Per esempio, la buccia verde dei limoni diventa gialla nel corso della maturazione a causa della conversione dei cloroplasti in cromoplasti. L’inverdimento delle patate, causato dall’esposizione alla luce, riflette la conversione degli amiloplasti in cloroplasti. In generale, i cromoplasti rappresentano una forma senescente di plastidio, non più convertibile. .CLOROPLASTI→organuli in cui avviene la fotosintesi, i proplastidi esposti alla luce del sole si differenziano in cloroplasti verdi grazie alla presenza della clorofilla, un pigmento fotosintetico. I Cloroplasti sono presenti in numero compreso tra 40-50 per cellula. Hanno un diametro di circa 4-6 micrometri. I cloroplasti sono piccoli e numerosi: vantaggio evolutivo→si spostano indipendentemente l’uno dall’altro per orientarsi a favore della radiazione luminosa. Il cloroplasto è delimitato da un involucro a doppia membrana lipo-proteica (teoria endosimbionte), nella membrana esterna vi sono porine che formano canali per molecole piccole; la membrana interna è altamente selettiva. Formati da due componenti principali: 1.Stroma→matrice amorfa 2.Tilacoidi granali→sistema di lamelle lineari o impilate tra loro a formare i grana; Sulle membrane dei tilacoidi sono ancorati clorofille e altri pigmenti accessori, raggruppati in unità fotosintetiche (fotosistemi P700 e P680). Nei tilacoidi si svolge la fase luminosa della fotosintesi, che corrisponde alla cattura e conversione dell'energia luminosa in energia chimica. 4 Nello stroma, avviene la fase oscura→organicazione dell’anidride carbonica (CO2). Nello stroma sono presenti: numerosi ribosomi, fibre di DNA plastidiale, granuli di amido primario. Quando i cloroplasti fotosintetizzano rapidamente producono una grande quantità di zuccheri, superiore alle necessità della cell, l’eccesso viene temporaneamente polimerizzato a formare granuli di amido (AMIDO PRIMARIO) all’interno dei cloroplasti stessi. *Amido secondario→amido contenuto negli amiloplasti. .LEUCOPLASTI→ Sono i plastidi meno differenziati, sono privi di tilacoidi e di qualunque tipo di pigmento. Derivano prevalentemente dal proplastidio. Tipi di leucoplasti classificati in base alle sostanze che producono/accumulano (lipidoplasti→lipidi, proteinoplasti→proteine, amiloplasti→carboidrati sotto forma di amido). I più comuni sono gli amiloplasti. In essi vengono accumulate le sostanze di riserva, in genere amido (anche proteine e lipidi). Il glucosio prodotto dalla fotosintesi viene condensato in amido (polisaccaride del glucosio) all'interno del cloroplasto; questo viene detto amido primario. L'amido primario viene poi idrolizzato e utilizzato per le necessità energetiche e plastiche della cellula. Ciò che rimane in eccesso viene trasportato fino ai tessuti di riserva, dove viene ricondensato come amido secondario all'interno dei leucoplasti (amiloplasti). A seconda dei materiali di riserva che contengono si dividono in: 1.AMILOPLASTI→deposito di amido della pianta • si trovano nei parenchimi amiliferi dei semi, fusti, radice e frutti. • hanno struttura molto più semplice di quello di un cloroplasto. • pochissimi tilacoidi. • enzimi per la sintesi e l’idrolisi dell’AMIDO SECONDARIO. • Talora gli amiloplasti si trasformano in cloroplasti. 2.LIPIDOPLASTI→presenza di lipidi. 3.PROTEOPLASTI→accumulano proteine. AMIDO L’amido è un polimero ad alto peso molecolare del GLUCOSIO. La sua sintesi avviene nei cloroplasti (speciali organuli presenti solo nelle cellule vegetali in cui avviene la fotosintesi). L’AMIDO è l’unione di due diversi polimeri: l’AMILOSIO e l’AMILOPECTINA, entrambi formati da GLUCOSIO. L’amido si accumula in granuli le cui dimensioni sono molto varie (1-150 μm di diametro). I granuli hanno forme molto caratteristiche, tanto che, dall’esame microscopico si può risalire alla specie. I caratteri morfologici dei granuli d’amido sono un ottimo elemento diagnostico e rappresentano una valida salvaguardia contro le frodi alimentari. FORMA: SPECIE-SPECIFICA. _ILO: centro di cristallizzazione del granulo d’amido. L’ilo può essere centrale, eccentrico, puntiforme, lineare, stellato e ramificato. Anche l’ilo è una caratteristica della specie. _STRATI CONCENTRICI: Stratificazione intorno all’ilo con differente grado di idratazione. Nei granuli di fagiolo e patata sono molto evidenti mentre nei granuli di frumento sono invisibili. I granuli di amido possono essere semplici o composti: _Semplici→la deposizione dell’amido inizia in un solo punto (ilo), indipendentemente dalla forma dell’ilo. _Composti→quando l’inizio dell’accumulo dell’amido avviene contemporaneamente in più punti diversi dello stesso amiloplasto. 5 .CROMOPLASTI→ Plastidi in cui manca un sistema tilacoidale vero e proprio. Si differenziano a partire dai cloroplasti, ma anche da leucoplasti e proplastidi. In essi avviene la sintesi e l’accumulo di pigmenti carotenoidi responsabili del colore giallo, arancio e rosso di molti fiori (Calendula e Ranuncolo), di alcuni frutti (pomodoro, peperone), e radici (carota, barbabietola). Il colore di fiori e frutti riveste un importante ruolo nell’attrazione di animali impollinatori e disseminatori. VACUOLO Il VACUOLO è presente SOLO nelle cellule vegetali. Mentre le cellule vanno assumendo la forma e le dimensioni definitive, compaiono nel citoplasma numerosi piccoli VACUOLI, che aumentano sempre di volume, durante la crescita delle cellule vegetali. Il vacuolo è rivestito da una membrana detta TONOPLASTO e contiene una soluzione acquosa detta SUCCO VACUOLARE. Tonoplasto→ricco di lipidi e proteine per l’attività enzimatica. Succo vacuolare→ sostanze disciolte o variamente condensate, PH intorno a 5. Contiene: 1.Sali di acidi organici e non (potassio, calcio, magnesio; acido citrico e ossalico) 2.carboidrati: glucosio e fruttosio (monosaccaridi), disaccaridi e polisaccaridi (inulina=funzioni di riserva). 3.amminoacidi e proteine (leucina, asparagina, tirosina, istidina), per lo più presenti nel succo vacuolare dei semi. 4.lipidi (semi) 5.metaboliti secondari 6.depositi minerali di natura cristallina, le cell che li producono si chiamano cell ossalifere (ossalato di calcio) e stanno nella radice e nelle foglie. Genesi e differenziamento vacuolo. Le cellule non differenziate normalmente posseggono iniziali vacuolari→PROVACUOLI; il processo di formazione del vacuolo normalmente inizia durante le prime fasi del differenziamento. Si ritiene che i vacuoli abbiano origine da una porzione del RE localizzata in prossimità della faccia trans del Golgi, costituita da un sistema tubulare membranoso interconnesso a cisterne appiattite. Funzioni dei vacuoli: I vacuoli svolgono importantissime funzioni: 1)RUOLO OSMOTICO (insieme al citoplasma ed alla parete cellulare) La crescita dell cell veg avviene per distensione e questa è dovuta specialmente all’aumento del contenuto d’acqua nel vacuolo. Il vacuolo svolge un ruolo importante nel mantenimento del turgore cellulare. L’ambiente extra cell è ipotonico rispetto al citoplasma, il citoplasma è a sua volta ipotonico rispetto al succo vacuolare; questo determina un flusso d’acqua in entrata nel vacuolo per osmosi e ne causa un aumento di volume. La pressione di turgore (esercitata dal vacuolo sulle pareti cell) contribuisce alla rigidità della struttura cell. La pressione di parete è la pressione meccanica esercitata dalla parete. Queste due pressioni si equilibrano. a) SUPPORTO MECCANICO: il vacuolo insieme alla parete cellulare realizza una struttura rigida (la pressione dell’acqua nel vacuolo viene controbilanciata dalla rigidità della parete cellulare) che determina la PRESSIONE DI TURGORE responsabile sia della distensione cellulare sia della rigidità di tessuti non lignificati (es. foglie, giovani fusti) i quali quando la pressione di turgore diminuisce avvizziscono. b) FORZA MOTRICE PER LA DISTENSIONE CELLULARE: La pressione di turgore esercitata dal vacuolo in maniera uniforme sulla parete cellulare, rappresenta la forza guida dell’accrescimento delle cellule vegetali. Se la parete non fosse in grado di opporsi al continuo 6 richiamo d’acqua si avrebbe lo scoppio dei protoplasti e se il vacuolo perdesse acqua, determinando l’abbassamento della pressione di turgore, si avrebbe la PLASMOLISI cellulare. L’abbandono dello stato meristematico e l’inizio della differenziazione comporta sempre l’accrescimento per distensione che modifica profondamente dimensioni, forma e struttura delle cellule. Le dimensioni, prima di tutto: se una cellula meristematica ha un diametro di 15-20µm, una cellula adulta è grande almeno 150-200µm (ma alcune cellule possono essere lunghe mm). La forma: dall’aspetto poliedrico si passa a forme in genere allungate, con una dimensione decisamente superiore alle altre. La struttura interna: se durante la crescita embrionale, nella cellula che raddoppia di volume, si assiste a una sintesi di tutti i componenti cellulari, nella crescita per distensione il grosso dell’aumento di volume (che può essere di centinaia di volte) è dovuto alla parete cellulare e al vacuolo, con un contributo meno rilevante dei plastidi. Poiché il vacuolo è costituito principalmente di acqua, possiamo dire che la crescita per distensione è caratterizzata da un massiccio ingresso di acqua nella cellula. L’assunzione e la perdita di acqua da parte di una cellula avvengono per osmosi. Lo stato di turgore di una cellula è la risultante di una condizione di equilibrio fra il potenziale osmotico della cellula (dovuto ai soluti disciolti) e il potenziale di pressione, dovuto alla parete. 2) RUOLO TAMPONANTE del pH citoplasmatico, EQUILIBRIO E RISERVA DI IONI. Il succo vacuolare ha un pH acido il cui valore in condizioni fisiologiche normali è pH 4-5. Questo valore è sempre notevolmente più basso rispetto a quello del citoplasma (pH 6.8-6.9). La capacità di mantenere il succo vacuolare ad un pH acido è dovuto alla presenza sul tonoplasto(membrana del vacuolo) di pompe protoniche (H+ ATPasi e H+ PPasi) che attraverso l’idrolisi dell’ATP e del pirofosfato inorganico, forniscono protoni al succo vacuolare acidificandolo. La presenza di pompe protoniche sul tonoplasto è di notevole importanza fisiologica poiché alla loro funzionalità è strettamente legata la capacità di trasportare selettivamente ioni e metaboliti dal citoplasma al vacuolo e, in misura minima dal vacuolo al citoplasma, permettendo così il mantenimento del pH costante del citoplasma. Nel vacuolo si accumulano molti ioni inorganici la cui natura e quantità è dovuta al tipo di terreno su cui cresce la pianta. Tutti gli ioni non immediatamente utilizzati vengono immagazzinati nel vacuolo mediante proteine trasportatrici a livello del TONOPLASTO (TRASPORTO ATTIVO). Piante che vivono su terreni ricchi di elementi tossici tendono ad accumularli nel vacuolo. Gli ioni nel vacuolo possono rimanere sotto forma libera oppure formare dei sali e dei cristalli con gli acidi organici accumulati nel vacuolo (es. ossalato di Calcio). In particolare cristalli di ossalato di calcio formano degli inclusi solidi insolubili di varie forme (rafidi, prismi o stiloidi, druse, sabbia cristallina). 3) RISERVA di METABOLITI Vacuolo→ruolo di detossificazione, accumula metaboliti potenzialmente tossici per la pianta. Metaboliti organici di riserva accumulati nel vacuolo→aminoacidi, proteine, zuccheri monosaccaridi (glucosio nell’uva), disaccaridi (saccarosio barbabietola e canna da zucchero). Gli acidi organici, quando in eccesso, vengono accumulati nel vacuolo per rimuoverli dal citoplasma, in quanto possono interferire in varie vie metaboliche, es. acido citrico (agrumi), acido malico, acido ossalico. Nelle cell dei tessuti di riserva dei semi le proteine sono accumulate all’interno di piccoli vacuoli→i GRANULI DI ALEURONE, sono piccoli vacuoli modificati per immagazzinare proteine, nei quali le proteine, 7 convogliate probabilmente attraverso il RE, precipitano in seguito alla disidratazione del seme durante la maturazione. 4) SEGREGAZIONE DI METABOLITI Alcuni METABOLITI SECONDARI, non coinvolti direttamente nei processi vitali della cellula, vengono riversati nel vacuolo come sostanze di rifiuto anche se vengono poi riutilizzati in altre vie metaboliche. Queste sostanze sono: a) ALCALOIDI: gruppo eterogeneo di composti caratterizzati da anello eterociclico contenente azoto, spesso basici e inodore. In genere sono veleni o principi attivi di piante medicinali, possono avere importanti effetti biologici su piante (difesa), parassiti ed animali. Esempi: colchicina e vinblastina bloccano la proliferazione cellulare; atropina, scopolamina, caffeina, papaverina, cocaina, efedrina, morfina, nicotina agiscono a livello del sistema nervoso centrale e periferico; stricnina, aconitina hanno effetti tossici. b) GLICOSIDI: composti formati da uno zucchero (il più comune è il glucosio) complessato ad una molecola non zuccherina. In generale sostanze di sapore amaro, abbastanza diffuse. Possono essere anche molto tossici es. glicosidi cianogenici che liberano per idrolisi acido cianidrico. I glicosidi→mandorle amare, mele, pesche, susine (amigdalina),sambuco, granoturco, sorgo (murrina), varie Leguminose. c) TERPENI (olii essenziali, resine, balsami): sono lipidi (quelli a basso peso melocolare sono volatili) di natura aromatica, insolubili in acqua. .Sono i principali componenti degli OLI ESSENZIALI e conferiscono alle piante un’ aroma profumato o gradevole. Presenti nei fiori, nelle foglie, nelle foglie e nei frutti di piante aromatiche (es. Labiate: basilico, salvia etc., e Apiaceae: finocchio, sedano, prezzemolo). .RESINE: Oleoresine (trementine) Gommoresine (es. mirra e incenso) Gomme naturali (es. caucciù dall’Hevea brasiliensis che è un politerpene). .BALSAMI: costituiti da olii essenziali e resine, liquidi e oncolore, o giallo-bruno, odore balsamico e sapore acre e amaro. e) FENOLI: composti organici che possiedono uno o più gruppi sostituenti ossidrilici (OH) legati a un anello aromatico. Comprendono i pigmenti che conferiscono i colori ai fiori alla frutta etc. (flavonoidiantiossidanti, antociani-colore blu/viola, flavoni e a volte tannini). f) TANNINI: polifenoli presenti nei vacuoli di corteccia e foglie di alcune piante, hanno attività antisettica per la pianta, quindi le conferiscono maggior resistenza contro i patogeni. 5) FUNZIONI LITICHE La presenza di idrolasi acide (peptidasi, glicosidasi, esterasi) nel vacuolo fa sì che esso venga considerato il più importante compartimento litico della cellula vegetale. Quando i vacuoli hanno funzione idrolitica sono analoghi ai LISOSOMI delle cellule animali. Le idrolasi vacuolari contribuiscono alla rottura e al riciclo dei componenti cellulari e garantiscono il ricambio delle strutture. I TESSUTI VEGETALI Tessuto→insieme di cellule che svolgono la stessa funzione e sono associate fra loro in modo congenito. I tessuti vegetali sono classificati in meristematici/embrionali e adulti/differenziati. Nelle piante a Cormo (=il cormo definisce una struttura della pianta costituita da radice, fusto e foglie che poi si evolveranno in tracheofite quando inizieranno a presentare i vasi per il trasporto della linfa.) diversi 8 tipi di cellule sono raggruppati in tessuti, ciascuno dei quali è specializzato per una determinata funzione. Questa specializzazione implica profonde modifiche rispetto allo schema base della cellula vegetale tipica. Tessuti Meristematici o embrionali→cell indifferenziate che si dividono, dalle quali traggono origine tutti i tessuti. Tessuti Adulti o definitivi→cell pienamente differenziate che non si dividono più (rare eccezioni). I tessuti presentano caratteri morfologici e funzionali caratterizzanti: - Presenza di una lamella mediana - Presenza di una parete che può presentare modificazioni - Presenza di plasmodesmi (canali): scambio tra cellule vive - Presenza di spazi intercellulari: scambio gassoso anche in tessuti profondi TESSUTI MERISTEMATICI Primari e Secondari: Le CELLULE MERISTEMATICHE Caratteristiche: .più piccole di quelle adulte (20 micrometri) .parete sottile pecto-cellulosica (manca parete secondaria) .provacuoli (vacuoli più piccoli) .il nucleo occupa la maggior parte del volume cell (non il vacuolo) .proplastidi, molti ribosomi. Meristemi primari→derivano da cell embrionali indifferenziate, il successivo differenziamento delle cellule meristematiche primarie da origine ai tessuti primari, che nel loro insieme costituiscono il corpo primario della pianta. I meristemi primari sono localizzati ai 2 poli della pianta: apice del fusto e apice della radice (meristemi apicali) e anche a livello dei nodi(=meristemi intercalari); sono responsabili della crescita in lunghezza della pianta (crescita indefinita). .Meristema apicale del fusto (apice caulino)=apice vegetativo (produce foglie e rami), nello stadio riproduttivo si trasforma in apice fiorale (origina sepali, petali, stami, carpelli). .Meristema apicale della radice (apice radicale)=forma radice principale e cuffia radicale. Meristemi secondari o cambiali→non presenti nell’embrione, ma nell’individuo adulto tra i tessuti definitivi. Solo in alcune piante (gimnosperme e alcune angiosperme dicotiledoni). Dalla divisione delle cell meristematiche secondarie si formano i tessuti secondari che costituiscono il corpo secondario. Le cell meristematiche secondarie si formano da cell adulte (mature) tramite “sdifferenziamento”: riacquiscono capacità di dividersi. Responsabili della crescita in spessore. Si distinguono in: 1)cambio subero fellodermico→forma sistema tegumentale del corpo secondario-produrrà sughero e felloderma2)cambio cribro vascolare/legnoso→genera il sistema conduttore-tessuto cribroso e vascolare- . Hanno in comune con quelli primari la capacità di moltiplicazione cellulare per mitosi e di produzione di cellule destinate a diventare adulte, ma hanno forma, dimensioni e proprietà citologiche differenti, a causa della loro origine: da tessuti adulti, appunto, e non dallo zigote. 9 La specializzazione delle cell meristematiche dipende dalla loro posizione nell’organo in via di sviluppo, la crescita della cell avviene per distensione (il vacuolo va occupare lo spazio diventando il compartimento cell più grande). Il differenziamento è il risultato di un programma genetico (attivazione/repressione di determinati geni). TESSUTI ADULTI o DEFINITIVI Costituiti da cell che perdono la capacità di dividersi, si differenziano e specializzano per svolgere una particolare funzione. 1)TESSUTI TEGUMENTALI (esterni e interni) Tessuti tegumentali esterni: . Epidermide→origine primaria. Ricopre il corpo della pianta (foglie radici e giovani fusti). Deriva dallo strato più superficiale dei meristemi apicali. La funzione fondamentale dell’epidermide è la protezione. Le cell epidermiche sono appiattite, disposte una accanto all’altra senza spazi intercellulari per evitare ingresso di patogeni; la forma varia in base all’organo che esse proteggono. Funzioni accessorie→fotosintesi (pigmenti disciolti nel vacuolo), secrezione, riserva di acqua,percezione degli stimoli, formazione di tessuto cicatriziale. Modificazioni della parete cell quali ad es mineralizzazione e lignificazione aumentano la resistenza della pianta. a)Tricomi ed emergenze: appendici epidermiche costituite da cell singole o complessi di cell,localizzate sulla superficie. Peli o tricomi=prodotti da epidermide, funzione protettiva, cell morte ripiene di aria (lucidi e biancastri), alcuni peli sono sensibili a stimoli tattili (piante insettivore: dionea). Emergenze=prodotte da epidermide e tessuti sottoepidermici, funzione simile a peli, spesso portano cell secretrici che invischiano e digeriscono piccole prede animali. b)stomi: aperture tramite cui si realizzano scambi gassosi, stoma=bocca. Sono formati da cell di guardia che delimitano una cavità detta apertura. Lo stoma si apre quando aumenta il turgore (le cell si distendono),che causa una variazione di forma delle cell di guardia e l’apertura per gli scambi gassosi. Se il turgore diminuisce lo stoma si chiude (le cell si restringono). c)velamen radicale: epidermide multipla, caratteristica di piante che devono economizzare acqua. Tessuto privo di spazi intercellulari, multistratificato, le cui cell muoiono e la cui parete presenta ispessimenti e perforazioni. Serve al rapido assorbimento di acqua e a ridurrne la perdita, funge da protettivo osmotico e meccanico. . Rizoderma→riveste le radici nelle parti non lignificate. Funzioni: protezione e assorbimento, infatti pareti cell sottili e niente cutina. Presenta peli assorbenti=peli radicali, sono strutture tubulose date da espansioni delle cell del rizoderma, man mano che la radice cresce se ne formano di nuovi. . Esoderma→origine primaria. Localizzato in: radice, fusti sotterranei. Mono o pluristratificato. Si forma in quanto la vita del rizoderma è di breve durata, quando il rizoderma cade e si disgrega i tessuti sottostanti assumono la funzione tegumentale (suberificano la parete) per garantire protezione. . Sughero→origine secondaria, funzione di barriera. Sostituisce l’epidermide quando, in seguito ad accrescimento secondario, questa non è più sufficiente a coprire il corpo della pianta e viene quindi lacerata ed eliminata. E’ formato da cellule morte a maturità, con pareti cellulari impregnate di suberina ed altre sostanze impermeabili che aiutano a ridurre l'evaporazione. 10 Periderma: tessuto che riveste il fusto e le radici delle piante legnose, una volta terminata la fase di crescita primaria, formando la corteccia esterna. Viene prodotto dal fellogeno o cambio subero-fellodermico, un'area meristematica che si origina nella regione periferica degli organi predisposti a crescita secondaria. L'attività del fellogeno porta alla produzione del sughero nella regione esterna e del felloderma internamente. Il felloderma è un tessuto parenchimatico con funzione di riserva. Tessuti tegumentali interni: Endoderma→tessuto tegumentale interno caratteristico della radice, ma presente anche nel rizoma e in alcuni fusti di piante acquatiche. Strato di cellule appressate che formano l'ultimo strato del cilindro corticale. Le pareti trasversali e radiali di queste cellule presentano una banda di materiale impermeabile, detta "del Caspary", che costituisce un ostacolo al movimento dei liquidi che fluiscono attraverso le pareti cellulari verso il cilindro centrale. La banda del Caspary viene tradizionalmente considerata come formata da un'impregnazione di sostanze tipo suberina e lignina. La funzione dell’endoderma è: impedire libero accesso di sostanze dannose ai tessuti conduttori presenti al suo interno, la banda del caspary è impermeabile e garantisce questa funzione. 2)TESSUTI PARENCHIMATICI I tessuti parenchimatici o fondamentali sono costituiti da cell vive che anche una volta mature mantengono la capacità di dividersi. Le cell presentano in genere solo la parete primaria, hanno grandi vacuoli. .Parenchima fondamentale: è costituito da cellule con forma poliedrica e parete sottile. Si trova in diverse parti della pianta, ad es. nella corteccia primaria del fusto e delle radici. Il tessuto parenchimatico può svolgere diverse funzioni in base a queste viene classificato in: .parenchima di riserva→specializzato nell’accumulo di sostanze di riserva di varia natura. Cell ricche di leucoplasti (amiloplasti, lipidoplasti, proteoplasti a seconda di ciò che accumulano). E’ localizzato in radice (corteccia,midollo). Molto sviluppato in organi specializzati x accumulo (bulbi, tuberi, frutti, semi). Situato nelle parti più profonde della pianta, non esposte direttamente alla luce (nella radice, ma anche nelle zone più interne dei fusti). Il più diffuso è il parenchima amilifero. .parenchima clorofilliano→detto clorenchima, cell con pareti sottili, facilmente attraversabili dalla luce, ricche di cloroplasti, è abbontante nel mesofillo fogliare. Il parenchima clorofilliano è formato da cellule cilindriche, tondeggianti o lobate, con un grande vacuolo centrale che preme i cloroplasti contro la parete cellulare in un unico strato favorendone la ricezione della luce e l'assorbimento della CO2. Nel corso del differenziamento i proplastidi sono diventati cloroplasti, presenti in numero molto variabile (20-30/cellula), dispersi nello strato di citosol periferico, adiacente alla parete cellulare. .p. aerifero→favorisce diffusione e circolazione dell’aria in piante sommerse o che si trovano in ambienti asfittici. Costituito da sistema di canali aeriferi e grandi spazi intercell. (Caratteristico di piante viventi in ambienti paludosi o acquitrinosi). .p. acquifero→specializzato nella conservazione dell’acqua, abbondante in piante grasse che stanno in ambienti aridi. Cell grandi, pareti sottili, vacuoli con mucillaggini che trattengono acqua. 3)TESSUTI CONDUTTORI Funzione di trasporto. Comparsi con aumento dimensioni. Cellule allungate sovrapposte in file, in direzione del trasporto. Il trasporto avviene in due direzioni: 1.acqua e nutrienti dal terreno→verso le foglie 2.sostanze prodotte dal metabolismo dalle foglie→verso tutte le zone che necessitano. 11 .Tessuto vascolare (xilema)→per il trasporto della linfa grezza (acqua e minerali) dalle radici al resto della pianta. Tracheidi e trachee: I vasi sono costituiti da cellule conduttrici allungate, che sono di due tipi: trachee e tracheidi. Le trachee si sono evolute indipendentemente in varie specie, a partire dalle tracheidi. Le trachee hanno un lume ampio e sono più corte. Le tracheidi sono più primitive, hanno un lume stretto e sono molto allungate. Le pareti longitudinale dei vasi vengono irrobustite x facilitare il trasporto. La loro parete primaria è sottile, la secondaria è lignificata. La parete secondaria lignificata può essere apposta contro la parete primaria: .in forma di anelli più o meno ravvicinati (tracheidi anulate), .di spirali (tracheidi, spiralate), .di combinazione dei due tipi (tracheidi anulo-spiralate) .o in modo più esteso (tracheidi reticolate-reticolo a maglie-, scalariformi- le punteggiature, cioè punti ovali non ispessiti, sono disposti come i pioli di una scala). La deposizione di parete secondaria è massima nelle tracheidi punteggiate, nelle quali le zone di parete primaria non lignificata, sono ridotte a numerose ma piccole aree circolari o ellittiche. Un tipo particolarmente elaborato di tracheide punteggiata è quello caratteristico delle Gimnosperme, detta tracheide con punteggiature areolate. .Tessuto cribroso (floema)→per il trasporto della linfa elaborata (acqua, zuccheri, amminoacidi), dai tessuti fotosintetici ai siti di utilizzo. I principali elementi conduttori del floema sono le cellule cribrose e gli elementi dei tubi cribrosi. I tubi cribrosi sono formati da cellule vive, ma a maturità la loro struttura interna è profondamente alterata: nucleo e ribosomi sono assenti, l’ultrastruttura di mitocondri e plastidi è rudimentale e alterata, il tonoplasto, in tutto o in parte, è stato distrutto, per cui citoplasma e succo vacuolare non sono più separati e al loro interno compaiono numerosi gruppi di fibrille proteiche. Le cellule dei tubi cribrosi sono allungate, sovrapposte le une alle altre, hanno una parete primaria, spesso sottile, non lignificata e sono provviste di “placche cribrose” (aree che mettono in comunicazione gli elementi adiacenti del tubo) costituite da grossi porocanali attraversati da plasmodesmi al cui livello spesso si riscontrano fascetti di fibrille proteiche. Fasci conduttori Xilema e floema: Elementi cribrosi e vascolari di origine primaria sono riuniti in fasci longitudinali in fusti e radici, mentre nella foglia la loro distribuzione varia in base ai taxa. Denominazioni: Fascio vascolare= fascio legnoso, xilema, legno-(origine primaria). Fascio cribroso=fascio liberiano, floema, libro → comprende cell parenchimatiche, cell compagne (annesse da plasmodesmi, partecipano al trasporto), parenchima del libro. I fasci vascolare e cribroso sono ravvicinati e formano il fascio cribro-vascolare. 4)TESSUTI MECCANICI Sostegno e resistenza al piegamento, due tipi in base alle caratteristiche della parete: .Collenchima→tessuto adulto primario. Cell vive. Situato in periferia sotto l’epidermide, si trova nelle parti della pianta in accrescimento (es germogli) ed è costituito da cellule vive con parete celluloso-pectica e non lignificata, ispessita in alcuni punti. (In base ai punti ispessiti vari tipi di collenchima: angolare, lamellare…). .Sclerenchima→costituito da cellule che dopo aver completato il differenziamento muoiono. Tipico delle parti della pianta che hanno completato la crescita per distensione. Le cellule presentano parete secondaria lignificata, è più resistente del collenchima, costituito da due tipi di cell: fibre e sclereidi. 12 Fibre: allungate e affusolate, aggregate in fasci o isolate, parete ispessita e lume ridotto. Due tipi di fibre, xilematiche (parete secondaria sempre lignificata) ed extraxilari (comprendono tutte le fibre che non fanno parte del legno). Sclereidi: cell allungate e brevemente ramificate, parete lignificata e ispessita solcata da molti poro-canali, presenti in tutto il corpo della pianta, morfologia varia (tondeggianti,a forma di stella). 5)TESSUTI SECRETORI Sono molto diversi fra loro in quanto a meccanismi, grado di specializzazione, localizzazione nella pianta. Le sostanze secrete (terpeni, tannini ecc..) rientrano nella categoria dei “prodotti secondari del metabolismo”. .peli secretori: superficie di foglie fusti e fiori, interagiscono con l’ambiente. Es: Nell’ ortica il pelo ghiandolare è costituito da una cell che secerne sostanze irritanti, avvolta alla base da un piedistallo. .ghiandole saline: ambienti litorali, secrezione di soluzioni con sali minerali. .Nettari: strutture ghiandolari che secernono un liquido contenente zuccheri, lipidi, con funzione di richiamo x impollinatori, comuni nel fiore. .canali secretori: strutture allungate con epitelio secernente che circonda un lume; talvolta l’epitelio è rivestito da una guaina; possono produrre resine, gomme. .tasche secretorie: strutture tipiche di foglie e frutti, esse riversano il loro secreto nel lume. (es limone, pompelmo, arancio). .lacticiferi: cell o file di cell che producono latice=emulsione acquosa che contiene gomme, resine o amido, enzimi; color latte, fuoriesce facilmente in seguito a ferita. IL FUSTO Il fusto o caule è l’organo di sostegno delle piante vascolari e stabilisce il collegamento tra le radici e le foglie. In fase giovanile può svolgere attività fotosintetica. E’ deputato alla conduzione. Accumula sostanze di riserva. MORFOLOGIA DEL FUSTO: Il germoglio comprende: .gemma apicale, costituita da bozze fogliari (foglie ai primi stadi di sviluppo)e cono vegetativo (gruppo di cell meristematiche da cui si origina il corpo primario del fusto). .foglie, si inseriscono nel fusto a livello dei nodi .nodi .internodi (regioni comprese tra un nodo e il nodo successivo) .gemme ascellari/laterali (inserite sopra le inserzioni fogliari), da cui traggono origine i rami laterali. DIFFERENZIAMENTO CORPO PRIMARIO DEL FUSTO Nel corpo primario del fusto, in sezione longitudinale, abbiamo le seguenti zone: cono vegetativo, zona di determinazione, zona di differenziamento, zona di struttura primaria. 1)Cono vegetativo: Da origine al corpo primario del fusto grazie all’attività meristematica di cellule iniziali (derivate da cell meristematiche dell’embrione). E’ costituito da tessuti primari. In alcune piante può esserci un corpo secondario che si origina dai meristemi laterali (cambio cribro legnoso e cambio subero fellodermico). Cono vegetativo 2 parti: .tunica, dal cui strato più esterno deriva l’epidermide del fusto (ne deriva il protoderma che poi origina l’epidermide). 13 .corpus, da cui derivano i tessuti che compongono il cilindro centrale. 2)Zona di determinazione In cui si realizza l’organizzazione cellulare in complessi meristematici specifici: .protoderma (dà luogo a epidermide), .meristema fondamentale (costituito da protocorteccia, che dà origine a parenchima corticale e protomidollo, che dà origine a parenchima midollare). .procambio (da cui originano i tessuti conduttori). In questa zona c’è la formazione di bozze fogliari e primordi dei rami. 3)Origine di bozze fogliari e primordi dei rami Compaiono sulla superficie del germoglio in corrispondenza della zona di determinazione. La formazione di foglie è dovuta a cell iniziali fogliari. Dopo la formazione di bozze fogliari si ha la formazione di primordi di rami. 4)Zona di differenziamento Le cell perdono le caratteristiche giovanili e iniziano a differenziardi. Si ha un incremento delle dimensioni del fusto per distensione (aumento volumetrico dovuto all’accumulo di acqua nelle cell). Differenziamento: Protoderma→da origine a corteccia e midollo nella ziona più interna del fusto. Procambio→cellule procambiali che daranno origine a protoxilema(verso interno fusto) e protofloema (verso esterno del fusto). STRUTTURA PRIMARIA DEL FUSTO Il corpo primario del fusto è la zona costituita da tessuti primari (meristemi primari). Nel fusto sono distinguibili 3 zone concentriche: epidermide, corteccia, cilindro centrale o stele. 1)epidermide: sistema tegumentale derivato dal protoderma, rivestita da cuticola (cutina e cere); dotata di stomi (assicurano scambi gassosi). 2)Corteccia: insieme di tessuti posti tra l’epidermide e il cilindro centrale, formata da strati di cellule parenchimatiche, può avere anche funzioni fotosintetiche (clorenchima/parenchima clorofilliano). 3)Cilindro centrale o stele costituito da: a)Sistema conduttore→fasci conduttori (cribro legnoso e cribro vascolare). Nei fasci conduttori di piante ad accrescimento secondario è presente il procambio (coinvolto nella formazione di tessuti conduttori primari), mentre nelle piante che non presentano accrescimento secondario è assente. .EUSTELE: I fasci conduttori sono disposti a formare un anello tra corteccia e midollo, in ogni fascio lo xilema è rivolto all’interno del fusto e il floema all’esteno. .ATACTOSTELE: i fasci conduttori sono immersi in tessuto parenchimatico, disposti apparentemente in disordine. b)Midollo→cell parenchimatiche ricche di amiloplasti, funzione di riserva. STRUTTURA SECONDARIA DEL FUSTO-DIFFERENZIAMENTO DEL CAMBIO CRIBRO LEGNOSO E SUBERO FELLODERMICO Dal cambio cribro-legnoso e dal cambio subero-fellodermico traggono origine i tessuti che costituiscono il corpo secondario della pianta. Meristemi laterali (a-b): a)cambio cribro legnoso→da cui traggono origine cell di xilema e floema secondari. Si forma dal procambio. b)cambio subero fellodermico/fellogeno→da origine al sughero verso l’esterno, mentre al felloderma verso l’interno. Fellogeno+sughero+felloderma=periderma. 14 L’attività del fellogeno è breve, quindi il sughero si stratifica per sovrapposizione in più anni. L’insieme di tessuti localizzati all’esterno del fellogeno= scorza (impropriamente corteccia). Gli scambi gassosi (sughero è impermeabile) sono assicurati dalle lenticelle (=strutture di cell arrotondate con parete non suberificata che lasciano spazi intercell attraverso cui l’aria arriva ai tessuti sottostanti). c) Xilema secondario→costituito da elementi vasali (trachhe/tracheidi); si origina per attività del cribro legnoso. Si compone di tessuto radiale(tessuti parenchimatici, funzione di collegamento e riserva) e assiale(tessuti conduttori, funzione meccanica e di riserva) compenetrati e con funzioni integrate. OMOXILO→xilema delle gimnosperme, è semplice e omogeneo. Presenta tracheidi con funzione di conduzione e sostegno. Spesso presenti dotti resiniferi ETEROXILO→xilema delle angiosperme. Presenta tracheidi e trachee, fibre di sostegno e cell parenchimatiche .Cerchie annuali: nella sezione trasversale dei fusti si osservano strati concentrici di xilema secondario definiti anelli di crescita; le cerchie sono riconoscibili grazie a differenze strutturali tra le pareti interne ed esterne di ogni strato, che si formano in tempi diversi. d)Floema secondario: costituito da cell e tubi cribrosi e parenchima. SPECIALIZZAZIONE E ADATTAMENTI DEL FUSTO Specializzazioni e adattamenti del fusto Fusti con funzione di riserva di nutrienti: tuberi, bulbi, bulbo-tuberi, rizomi (fusti ingrossati, sotterranei o lungo la superficie del terreno). Fusti con funzione di riserva idrica: succulente (piante grasse). Fusti con funzione di sostegno: viticci Fusti con funzione fotosintetica: cladodi (es asparagi) Fusti con funzione di difesa: spine(foglie o rami modificati), emergenze(protuberane della corteccia). LA FOGLIA Organo per lo più laminare che nasce sempre sul fusto o sulle sue ramificazione. Funzioni: fotosintesi e traspirazione. (foglie con funzione clorofilliana=nomofilli). Altre funzioni di foglie adattate per altri scopi: .accumulo di nutrienti→cotiledoni, sono le prime foglie della plantula sviluppate allo stadio embrionale. .difesa→spine fogliari (es piante climi aridi, x difesa meccanica, non fotosintetizzanti) .riserva d’acqua→foglie succulente .sostegno→cirri (piante rampicanti) .Sclerofille→foglie coriacee di piante adatttate x ambienti aridi. .Foglie delle conifere→aghiformi, quasi tutte persistenti. .Cattura di insetti e digestione→foglie delle piante insettivore. Due tipi di foglie con diversa origine: Microfilli→sistema vascolare formato da un solo fascio conduttore, piante microfogliate (piccole foglie). Macrofilli→sistema conduttore complesso, formato da più fasci conduttori, piante macrofogliate (grandi foglie-felci). Filotassi= disposizione delle foglie sul fusto. 1)filotassi alternata: ad ogni nodo 1 foglia. 2)filotassi verticale: 1 nodo, più foglie. 3)filotassi oppsta: 1 nodo, 2 foglie. 4)filotassi distica: le foglie sono disposte su due file. 5)filotassi elicoidale: 1 foglia per nodo, foglie inserite a spirale intorno al fusto. 15 ANATOMIA .Sistema tegumentale: epidermide fogliare, regola gli scambi gassosi, lascia filtrare la luce x far si che arrivi al parenchima fotosintetico quindi ha cell epidermiche incolori. Presenta spesso peli per la difesa meccanica e la prevenzione da eccessiva traspirazione. Le variazioni della struttura dipendono dagli habitat: .Mesofite (ambienti né troppo umidi, né troppo caldi) .Idrofite (piante acquatiche) .Xerofite (piante in ambienti secchi) .Sistema fondamentale: parenchima clorofilliano, 2 tipi: 1.Parenchima a palizzata→sul lato assiale della foglia; fotosintetico, tra una cell e l’altra spazi intercell con ampiezza ridotta. 2.Parecnhima spugnoso→sul lato abissale della foglia; cell di forma irregolare con numerosi e ampi spazi intercell, cloroplasti in num minore. a)foglie bifacciali: palizzata faccia ssiale, spugnoso faccia abissale. b)isofacciali: palizzata faccia assiale e abissale, spugnoso nel mezzo. c)unifacciali: una delle due facce è ridotta o assente. .Sistema conduttore: formato dall’insieme delle nervature, formate a loro volta dall’insieme dei fasci. Fasci: 1.xilema (faccia assiale) 2.floema (faccia abissale) Nervature maggiori→calibro superiore, trasporto aq e minerali da fusto a foglie tramite xilema, trasporto prodotti di fotosintesi da parenchima a siti di uso attraverso floema. Nervature minori→calibro inferiore, scambio sostanze con cell parenchimatiche. Genesi e sviluppo .Procambio→tessuti conduttori .Allungamento foglie→1.cell meristematiche, 2.meristema intercalare. .Crescita in larghezza→meristemi marginali. GUTTAZIONE: l'eliminazione di acqua allo stato liquido dalle foglie, si verifica quando per l'eccessiva umidità atmosferica, la traspirazione (perdita di acqua sotto forma di vapore) non può avvenire regolarmente, o quando la quantità d'acqua assorbita dalle radici è superiore a quella traspirata dalle foglie. Il fenomeno avviene attraverso gli stomi (bocche, aperture). Funzioni della foglia 1)Fotosintesi 2)Regolazione della perdita di acqua (stomi) 3)Riserva (aloe) 4)Protezione (spine, scaglie) 5)Attrazione e cattura (piante carnivore) Stomi-bocche→formati da cell di guardia (contengono clorolpasti) che somigliano a 2 labbra, servono x scambi gassosi tra interno ed esterno pianta; si trovano sull’epidermide. Quando le cellule di guardia perdono Acqua si restringono e lo stoma si chiude; 16 Quando le cellule di guardia assorbono Acqua si rigonfiano e le pareti interne Si separano per distensione delle Pareti esterne, lo stoma si apre. LA RADICE Organo generalmente sotterraneo, specializzato nell’ assorbimento dell’acqua e dei sali minerali dal terreno, che verranno poi portati a tutta la pianta. Può servire come da sito di accumulo di amido che verrà usato durante la ripresa vegetativa e per lo sviluppo di fiori e frutti. La radice primaria è la prima a svilupparsi e origina dall’apice radicale embrionale; la radice secondaria si sviluppa dalla primaria, il sistema radicale si irradia in tutte le direzioni e ancora la pianta al terreno. Vi è un forte legame tra lo sviluppo della radice e quello del germoglio. FUNZIONI: ASSORBIMENTO ANCORAGGIO RISERVA PRODUZIONE DI ORMONI e di sostanze di difesa. Struttura: L’apice radicale è protetto dalla cuffia (tessuto protettivo); l’accrescimento della radice avviene dall’apice radicale grazie a un meristema apicale. La zona della radice sopra l’apice è costituita da cell in fase di differenziazione: la zona di differenziazione, questa comprende una zona liscia che cresce per distensione tramite l’ingresso d’acqua nel vacuolo. Poi al di sopra c’è la zona di struttura primaria che comprende una zona pilifera: i peli radicali sono strutture per l’assorbimento capillare di acqua e Sali minerali. Al di sopra della zona pilifera si originano e crescono le radici laterali. Nelle gimnosperme e in alcune angiosperme c’è anche una zona di struttura secondaria caratterizzata da tessuti che hanno origine da meristemi secondari. Cuffia: ricopre l’apice radicale, tessuto protettivo contro l’abrasione causata dalla penetrazione nel terreno. Le cell si rinnovano costantemente anche perché nella porzione terminale c’è un continuo sfaldamento dovuto alla crescita. Cell della Columella→contengono granuli di amido, coinvolte nella percezione della gravità. Struttura primaria sezione trasversale, 3 zone: 1)rizoderma→esterno, funzione protettiva, pareti non cutinizzate, assorbimento acqua. Dopo la caduta del rizoderma la radice diviene rivestita da esoderma. 2)cilindro corticale→intermedio (comprende parenchima corticale di riserva non suberificato, e più internamente endoderma suberificato con bande del Caspary) 3)cilindro centrale/stele→interno, contiene il sistema vascolare. Dalla sua parte esterna (il periciclo) prendono origine le radici secondarie. Nella radice xilema e floema hanno disposizione raggiata, detta actinostele; si alternano arche xilematiche e arche floematiche e sono sempre in numero uguale. Struttura secondaria: Solo gimnosperme e alcune angiosperme. Alla struttura primaria che è breve segue la struttura secondaria che si estende x il resto di tutta la radice. E’ cosstituita da tessuti secondari, il suo accrescimento è determinato dal cambio cribro legnoso e dal cambio subero fellodermico. Il cambio cribro legnoso si origina nel cilindro centrale e segna il passaggio alla struttura secondaria; genera xilema e floema secondario. Grazie al fellogeno il rizoderma si modifica e al suo posto abbiamo il sughero verso l’esterno 8t. tegumentario secondario) e il felloderma verso l’interno. 17 1)Apparato radicale a fittone: con radice primaria (di diretta derivazione embrionale) preminente per tutta la vita della pianta e radici laterali (o secondarie) a minore sviluppo. 2)Apparato radicale fascicolato: la radice primaria muore presto, l’apparato presenta radici laterali (ed avventizie) molto sviluppate e superficiali. Specializzazioni e adattamenti: 1.Radici tuberiformi→riserva e accumulo. Strutture rigonfie e ingrossate ricche di acqua e nutrienti (es ravanelli, barbabietole da zucchero, carote), spesso presentano molecole aromatiche con attività biologiche (ginseng). Le radici tuberiformi da sole non possono generare una nuova pianta perché non hanno gemme nella parte ingrossata, ma portano la nuova vegetazione dove radice e fusto si incontrano. 2.Pneumatofori→radici respiratorie che crescono verso l’alto, tipiche di zone paludose, assorbono ossigeno dall’aria, presentano nella struttura numerose lenticelle. 3.Austori→apparati succhiatori delle piante epifite, utilizzano le sostanze elaborate dalle piante ospiti. 4.Micorrize→associazione simbiontica tra funghi e radici: la pianta fornisce ai funghi i carboidrati prodotti con la fotosintesi, i funghi micorrizici metaboizzano i minerali presenti nel suolo e li rendono biodisponibili alle radici della pianta. ORGANI RIPRODUTTIVI IL FIORE Il fiore è l’apparato preposto alla riproduzione tipico delle Fanerogame, composto dall’insieme di: .parti fertili (sporofilli) = STAMI + PISTILLO .parti accessorie (antofilli) = SEPALI + PETALI È un germoglio a crescita determinata con internodi non sviluppati e foglie modificate. Un fiore completo ed ermafrodito è composto da: .calice, formato da antofilli verdi detti sepali. .corolla, formata da antofilli colorati detti petali. .androceo, composta da foglie trasformate staminali dette stami. .gineceo, composto da foglie carpellari dette carpelli o pistilli. .Perianzio: sepali e petali sono ben differenziati tra loro, si identificano il calice e la corolla. .Perigonio: formato da tepali sepali e petali non sono distinguibili tra loro Simmetria •Fiori actinomorfi: sono evidenti + piani di simmetria, cioè sono divisibili in più parti uguali (simmetria raggiata). •Fiori zigomorfi: è individuabile un solo piano di simmetria che divide il fiore in due parti uguali. •Fiori asimmetrici: non è possibile individuare piani di simmetria. CALICE Verticillo più esterno del fiore, formato dai sepali, generalmente colore verde, con funzione protettiva e clorofilliana. Saldatura: gli organi fogliari possono essere liberi o saldati l’uno con l’altro. dentato: se la saldatura dei sepali è totale, partito: se la saldatura è solo alla base. Nella saldatura si forma un tubo che si apre in una fauce, sopra la quale si trova la parte libera del calice cioè il lembo. 18 COROLLA Ha funzione vessillare (attrazione insetti) per le varie colorazioni. La parte ristretta del petalo, quella innestata nel ricettacolo della corolla, è detta unghia, mentre la parte più larga, che si espande all'esterno, ha il nome di lamina o lembo. Alla base del petalo si trova spesso il nettario, la ghiandola che secerne il nettare. La corolla dura poco: dopo la fecondazione appassisce e dissecca. Rispetto alla saldatura, anche la corolla può essere: 1.Dialipetala: con petali liberi, 2.Gamopetala: con petali fusi e uniti tra loro. Le più evidenti manifestazioni di evoluzione nella struttura del fiore si osservano nella struttura della corolla: la forma comparsa più anticamente è dialipetala, ovvero con uno svariato numero di petali separati (Rosa canina), mentre è più evoluta quella gamopetala, cioè composta da petali saldati tra loro (Digitalis purpurea) che forniscono una protezione maggiore a stami e pistilli. Dal punto di vista della simmetria, anche le corolle possono essere: 1.actinomorfe o regolari (più piani di simmetria) 2.zigomorfe o irregolari (asimmetrica sul piano orizzontale). Rispetto alla saldatura, anche la corolla può essere Dialipetala: con petali liberi Gamopetala: con petali fusi e uniti tra loro. Le più evidenti manifestazioni di evoluzione nella struttura del fiore si osservano nella struttura della corolla: la forma comparsa più anticamente è dialipetala, ovvero con uno svariato numero di petali separati (Rosa canina), mentre è più evoluta quella gamopetala, cioè composta da petali saldati tra loro (Digitalis purpurea) che forniscono una protezione maggiore a stami e pistilli. Dal punto di vista della simmetria, anche le corolle possono essere: actinomorfe o regolari (più piani di simmetria) zigomorfe o irregolari (asimmetrica sul piano orizzontale). 19 ANDROCEO apparato riproduttore maschile Formato da stami fatti di un filamento e di una parte fertile detta antera Le antere sono formate da due teche a loro volta divise in due sacche o logge polliniche Nelle sacche sono contenute le microspore che origineranno i granuli pollinici che verranno liberati con varie modalità. Numero di stami: L’androceo può essere formato da uno, due o più filamenti. Saldatura degli stami: possono essere liberi, oppure uniti assieme concresciuti (si valuta quanti fascetti formano). Gli stami sono organi riproduttivi maschili che contengono i granuli di polline. Nelle specie più primitive essi avevano l'importante funzione di attrarre con il loro numero e con il loro colore i primi insetti impollinatori, i coleotteri in particolare, che si nutrivano di essi. La riduzione del numero di elementi fiorali è considerata un carattere più evoluto e rispecchia la coevoluzione con altri gruppi di insetti specializzatisi, ad esempio, nel succhiare il nettare. GINECEO Gineceo: apparato riproduttore femminile Può essere costituito da uno o più pistilli Ogni pistillo è costituito da ovario, stilo e stigma Nell’ovario, che dopo la fecondazione si trasformerà in frutto, sono contenuti gli ovuli, che in seguito origineranno i semi. La parete dell’ovario può essere costituita da uno o più carpelli (foglie modificate) che si fondono tra loro. I margini carpellari che si sono fusi formano un tessuto: placenta, sulla quale si inseriscono gli ovuli mediante un filamento: funicolo. Numero di carpelli: 1)monocarpellare: il gineceo può essere formato da un solo carpello, 2)pluricarpellare: formato da più carpelli e in questo caso possono rimanere tra loro indipendenti e formare tanti pistilli oppure concrescere e formare un solo pistillo. 20 Aspetto della regione ovariale: si possono distinguere: 1)ovari uniloculari formati da una o più foglie carpellari saldate per i margini a costituire una sola cavità. 2)Ovari pluriloculari fatti da più foglie carpellari che ripiegandosi verso l’interno provocano la formazione di tanti setti che dividono la cavità in tante logge. Placentazione: disposizione degli ovuli sulla placenta. Posizione dell’ovario nel ricettacolo: Ovario supero: si inserisce sopra il ricettacolo fiorale da cui emerge in modo evidente. Ovario semiinfero: è parzialmente immerso nel ricettacolo. Ovario infero: è completamente immerso nel ricettacolo, emerge lo stilo. La protezione dell'ovario è un elemento ricorrente nell'interpretazione della direzione evolutiva. L'embrione in esso contenuto è portatore dei caratteri ereditari e delle ricombinazioni geniche conseguenti la fecondazione. Ciò spiegherebbe, ad esempio l'affermazione degli ovari inferi, ovvero inseriti inferiormente agli altri elementi fiorali, stami, coralla e calice. In tale posizione è garantita una maggiore protezione del prezioso contenuto genetico e una maggiore probabilità che questo si sviluppi senza danni. Calice: costituito da sepali (generalmente verdi con funzione fotosintetica e protettiva); 1)dialisepalo: sepali liberi, 2)gamosepalo o sinsepalo: sepali saldati tra loro. Può essere caduco o persistente (se rimane dopo la fecondazione). Corolla: parte del fiore deputata alla funzione vessillare. Costituita da più petali. Corolla dialipetala (petali separati), gamopetala o sinpetala (petali fusi tra loro). Androceo: apparato riproduttore maschile. Stame: filamento che sostiene la parte fertile (antera). Le antere sono costituite da due teche che a loro volta contengono due sacche polliniche. Gineceo: apparato riproduttore femminile. Pistillo: composto da ovario, stilo e stimma. L’ovario può essere costituito da uno o più carpelli. L’inserzione dell’ovario nel ricettacolo determina la sua condizione di ovario infero, ovario supero). Il fiore può essere suddiviso in base agli elementi sessuali: 1)Bisessuato (ermafrodito - perfetto): se possiede stami e carpelli, cioè sia l’apparato maschile sia quello femminile. 2)Unisessuato (imperfetto): se manca uno dei due apparati, quindi possiede o solo l’apparato femminile (pistillifero) o solo quello maschile (staminifero). I diversi tipi di fiore determinano anche il sesso della pianta: a) Piante monoiche (una casa): se fiori maschili e femminili sono sulla stessa pianta, o come fiori unisessuati o come fiori ermafroditi (mais, quercia, betulla …). b) Piante dioiche (due case): se un individuo porta solo fiori maschili e uno solo femminili (tasso, salice…), quindi avremo una pianta maschile ed una femminile. c) Piante poligame se sulla stessa pianta sono presenti fiori ermafroditi ed unisessuati. DOPPIA FECONDAZIONE Attraverso il tubetto pollinico il granulo di polline scarica due nuclei aploidi nel sacco embrionale. Uno si fonde col nucleo della cellula uovo, l’altro col nucleo dell’endosperma secondario. I due nuclei fecondati cominciano subito a dividersi. Dalla cellula uovo fecondata si sviluppa l’embrione (diploide); dal nucleo dell’endosperma secondario si sviluppa l’endosperma (triploide). 21 IMPOLLINAZIONE: Vento e Insetti Le Angiosperme più antiche si ritiene siano state anemogame (impollinate dal vento). Con la diffusione degli insetti, a partire da 100 milioni di anni fa, esse passarono gradualmente alla impollinazione entomogama: dal Cretaceo si ebbe una continua coevoluzione tra piante-insetti impollinatori che portò ad una progressiva trasformazione sia morfologica, soprattutto dei fiori e degli apparati boccali di insetti, che biochimica, con la sintesi di composti chimici di richiamo (pigmenti, odori, sostanze zuccherine). In alcune specie l'evoluzione della modalità di impollinazione entomogama ha portato ad una estrema specializzazione legata alla comparsa di strutture altamente complesse. La salvia comune (Salvia pratensis) ha un petalo trasformato in modo tale da favorire l'appoggio dell'insetto ed un sistema di bilanciamento dei 2 stami (ulteriore specializzazione) che fa in modo che essi si pieghino in avanti all'arrivo di quest'ultimo, così da applicare granuli di polline maturi sul suo dorso. Alcune orchidee (Ophrys sphegodes) simulano con forme e colori del labello la femmina dell'insetto pronubo, altre hanno sviluppato passaggi obbligati in ingresso e in uscita dal fiore. Nel fiore di rosmarino gli stami e lo stimma sono inarcati verso l’alto, sporgono dal fiore ed entrano in contatto col dorso peloso dove depositano altro polline. COEVOLUZIONE Due o più specie agiscono con forza selettiva reciproca inducendo modificazioni. Le pressioni evolutive esercitate da differenti condizioni ambientali possono aver influito sulle modalità di impollinazione di alcune specie sia vegetali che animali. - Coleotteri: molto sviluppato l’olfatto. Sono attirati da fiori grandi o da infiorescenze con corolle bianche o neutre ma che emanano un odore forte (fruttato, speziato o maleodorante da fermentazione). -Api: vivono di nettare, che serve anche per nutrire le larve. Hanno modificato e adattato parti del corpo per trasportare nettare e polline: setole grossolane, apparato buccale allungato per la suzione. I fiori presentano petali vistosi e brillanti (blu, gialli) con nettario situato alla base della corolla unita a tubo e spesso con “piattaforma di atteraggio” e “indicatori del miele”. - Farfalle: combinazione di vista ed odorato. Presentano un apparato buccale modificato per la suzione. Percepiscono il rosso come colore distintivo. In alcuni generi di Asteracee come Ambrosia e Artemisia la pressione ambientale ha indotto il passaggio dall'entomogamia all'anemogamia. E’ la risposta di adattamento ad ambienti asciutti, soleggiati e ventosi, per favorire l'impollinazione di un gran numero di fiori. I fiori impollinati dal vento non producono nettari, hanno colori smorti e sono generalmente inodori. Infiorescenze: insieme dell’asse fiorale e dei fiori riuniti insieme INFIORESCENZE INDEFINITE accrescimento indefinito (racemose): Presentano un asse principale che tende ad allungarsi indefinitamente con produzione laterale di nuovi fiori. I fiori nuovi sono posti nelle vicinanze dell’apice e quelli più vecchi vicino alla base dell’infiorescenza (accrescimento centrifugo). Si distinguono: 1)Infiorescenze con asse allungato longitudinalmente 2)Infiorescenze con asse principale raccorciato INFIORESCENZE INDEFINITE: indefinito, l’asse di accrescimento produce solo fiori laterali -Infiorescenze con asse allungato longitudinalmente: • racemo o grappolo→ ha fiori peduncolati (giacinto, ippocastano, digitale...); • spiga→ ha fiori sessili (graminacee: grano, orzo, riso …); • amento→ con l’asse flessibile e pendulo (nocciolo, pioppo, salice, quercia, betulla …); 22 • spadice→con l’asse ingrossato ed una brattea (spata) più o meno avvolgente (calla, anthuriun ..); • corimbo→ racemo con peduncoli fiorali che giungono tutti allo stesso livello (pero, biancospino) -Infiorescenze ad asse raccorciato: • ombrella, peduncoli fiorali di uguale lunghezza, inseriti nello stesso punto in modo che i fiori formino un’ombrella (finocchio, carota, geranio, primula …) -Infiorescenze con l’asse raccorciato ed ingrossato: •capolino, i fiori sessili si inseriscono su una dilatazione dell’asse infiorescente (girasoli, dalia, margherite, gerbere, calendula …) INFIORESCENZE DEFINITE accrescimento definito (cimose): Presentano un asse principale che termina con un fiore al di sotto del quale si formano nuovi assi con nuovi fiori terminali. I fiori vecchi sono più prossimi all’apice mentre quelli più giovani sono più lontani (accrescimento centripeto). Accrescimento (indefinito e definito): acropeto (verso alto), acrofugo (verso basso), centrifugo e centripeto. INFIORESCENZE DEFINITE: l’asse principale dell’infiorescenza termina con un fiore .Monocasio o cima unipara: sotto il fiore apicale si forma un solo ramo che porterà il fiore terminale .Dicasio o cima bipara: sotto al fiore terminale dell’asse principale, si formano due assi laterali opposti con fiore terminale e che continuano a ramificarsi nello stesso modo Infiorescenze composte: l’asse infiorescenziale si ramifica originando molti assi distinti in: - Pannocchia - Spiga composta - Ombrella composta - Corimbo composto I fiori delle asteracee (composite) sono raggruppati in capolino. Il capolino delle composite ha l’aspetto di un unico grande fiore, mentre in realtà è formato da tanti piccoli fiori strettamente raggruppati. Ognuno di questi minuscoli fiori ha un ovario infero. Gli stami sono di solito fusi tra loro e con la corolla. I petali sono anch’essi fusi tra loro e con l’ovario, mentre i sepali sono assenti o ridotti ad una serie di squame (pappo). Si distinguono: • Fiori del disco (tubulosi) che costituiscono la parte centrale; • Fiori del raggio (ligulati) disposti all’estremità periferica, sono spesso femminili. I fiori individuali che formano il capolino maturano in tempi diversi seguendo una spirale centripeta. Importanza economico farmaceutica del fiore: Farmaceuticamente sono fonte di: oli essenziali, flavonoidi, pigmenti ecc. Riportate in Farmacopea: •camomilla e.s., •calendula (Ph.Eur.), •lavanda e o.e. (Ph Eur.), •arancio amaro o.e. •essenza di neroli (Ph. Eur.), •chiodi di garofano (Ph. Eur.), •stimmi zafferano (Ph. Eur) 23 IL SEME Struttura tipica delle spermatofite (piante a seme), comprende gimnosperme e angiosperme. Il seme è un ovulo maturo che contiene un embrione e del materiale di riserva (endosperma), avvolto da 1 o 2 tegumenti. L’embrione è formato da un asse su cui si inseriscono i cotiledoni, sopra abbiamo l’epicotile con il meristema apicale cauliano (o del germoglio), al di sotto dei cotiledoni c’è l’ipocotile con il meristema apicale radicale. A seme maturo i tegumenti dell’ovulo si induriscono e diventano tegumenti del seme. Il seme svogle due funzioni fondamentali: 1.propagazione della specie nel territotio, occupando zone sempre più vaste. 2.protegge l’embrione anche in condizioni avverse. Per facilitare la dispersione e quindi la colonizzazione di nuovi ambienti, nel corso della selezione sono stati favoriti: semi di piccole dimensioni, semi che hanno sviluppato particolari dispositivi allo scopo di facilitare la dispersione, ad esempio: .semi con strutture piumose per volare (pioppo) .semi che vengono trattenuti all’interno del frutto che possiede uncini per potersi aggrappare al pelo degli animali .semi che possono restare vitali per molti anni (dormienza). Embriogenesi: eventi che vanno dalla fecondazione alla formazione dell’embrione. La prima divisione dello zigote da origine a cell basale (forma sospensore che ancora embrione e trasferisce sostanze nutrienti) e cell apicale (crea embrione vero e proprio). Gli organi della pianta adulta si formano tramite meristemi dopo l’embriogenesi, il piano strutturale è stabilito già nell’embrione (polarità): meristema apicale germoglio (forma parte aerea) ad un estremità e meristema apicale radicale (forma apparato radicale) all’estremità opposta. Quiescenza: prima di essere rilasciato dalla pianta madre il seme perde fino al 90% d’acqau ed entra in uno stato di quiescenza, questo gli permette di sopravvivere per lungo tempo in attesa delle condizioni più favorevoli. Nelle prime fasi di germinazione il seme non può fotosintetizzare perché è avvolto dai tegumenti, le sostanze di riserva accumulate nel seme gli permettono di sopravvivere fino al momento in cui la pianta diverrà fotosintetica grazie alle prime foglie. Le sostanze di riserva sono accumulate nell’endosperma. In molte specie l’endosperma si esaurisce durante le prime fasi, in questo caso le sostanze di riserva sono depositate nei cotiledoni. Semi albuminosi→riserve endosperma Semi esalbuminosi→riserve cotiledoni Le sostanze di riserva nei semi sono di 3 tipi: 1.carboidrati all’interno degli amiloplasti, 2.grassi sotto forma di goccioline citoplasmatiche, 3.proteine accumulate nei granuli di aleurone. La quantità e la proporzione variano a seconda della specie. In base alla distribuzione delle riserve si possono dividere i semi in alcuni gruppi: a) semi che contengono in prevalenza grassi e proteine (ricino, soia, girasole). b) semi che contengono in prevalenza proteine e amido (pisello, fagiolo, leguminose). c) semi con netta prevalenza di amido accompagnato da quantità minore di proteine e grassi (tutti i semi dei cereali) d) semi che contengono in abbondanza tutti le sostanze di riserva (arachide) 24 Modalità di dispersione dei semi: 1.attraverso il vento, semi leggeri (orchidee); semi con appendici pelose o alate. 2.attraverso gli animali: tramite strutture ad uncino che si aggrappano al pelo, l’animale poi pulendosi deposita sul terreno e il seme può germinare. Tramite frutti commestibili che vengono mangiati, poi i semi rilasciati con gli escrementi possono germinare. 3.dispersione attraverso acqua piovana, corsi d’acqua, correnti marine. 4.per autodisseminazione: particolari frutti a maturazione avvenuta contengono gas sotto pressione e spruzzano a distanza semi e sostanze mucillagginose e la capsula vuota che li racchiudeva in direzioni opposte. Germinazione Il seme controlla l’ambiente aspettando il momento favorevole. I 3 fattori fondamentali che interrompono lo stato di quiescenza del seme sono: .presenza di acqua→il seme deve imbibirsi d’acqua per riattivare il metabolismo .presenza di ossigeno a sufficienza →x sostenere il metabolismo .temperatura adeguata Fasi germinazione comuni a tutti i semi : 1.imbibizione del seme, seguita dalla riattivazione del metabolismo, 2.l’embrione inizia a crescere: il rpimo organo ad uscire è la radice primaria (ancoraggio e assorbimento di acqua e nutrienti). .Germinazione epigea (sopra la terra)→ nel fagiolo, i cotiledoni escono dal terreno insieme all’apice. .Germiazione ipogea (sotto la terra)→pisello, i cotiledoni restano nel terreno. IL FRUTTO Il frutto è l'organo vegetale in cui si trovano racchiusi i semi. E’ presente nelle spermatofite, ossia nelle piante produttrici di semi (gimnosperme e angiosperme). Il frutto è l'organo vegetale in cui si trovano racchiusi i semi. E’ presente nelle spermatofite, ossia nelle piante produttrici di semi (gimnosperme e angiosperme). Veri frutti= si sviluppano dal gineceo in seguito alla fecondazione: l’ovario si trasforma in frutto e gli ovuli in semi. Se alla formazione del frutto partecipano altri organi si parla di falsi frutti. .Classificazione secondo l’origine da un singolo carpello (apocarpici, es legume e dattero) o da più carpelli saldati (sincarpici, es oliva e uva). .Classificazione secondo le caratteristiche del pericarpo(=involucro che costituisce il frutto) : a)Frutti secchi→disidratati e coriacei b)Frutti carnosi→ricchi d’acqua e con la maturazione acquisiscono colore, sapore e profumo. Nei frutti il pericarpo, derivato dai carpelli dell’ovario, può essere diviso in 3 strati: 1) esocarpo: è l'epidermide esterna, spesso determina i caratteri organolettici dei frutti. 2) mesocarpo: è il tessuto della zona centrale del frutto. Costituisce la polpa nei frutti carnosi. 3) endocarpo: è la parte più interna che può anche lignificare dando origine ad una cavità dove sono alloggiati i semi. Può contenere sclereidi che aiutano l'identificazione della specie. Formazione e maturazione del frutto Normalmente seme e frutto si sviluppano insieme dopo la fecondazione grazie a segnali ormonali portati dal tubetto pollinico. Successivamente nei frutti secchi avvengono la disidratazione e a volte sclerificazione 25 che preparano il frutto per l’apertura (deiscenza), mentre nei frutti carnosi avviene il processo di maturazione (acquisizione colore sapore..) e il rammollimento della polpa. CLASSIFICAZIONE FRUTTI SEMPLICI: derivano da 1 solo fiore, si distinguono in base alla consistenza del pericarpo 1.FRUTTI SECCHI→scarsi parenchimi e un contenuto di acqua piuttosto basso; possono distinguersi in: a)indeiscenti, contengono di solito 1 solo seme (monospermi) e restano chiusi mantenendo il seme al loro interno. b)deiscenti, tipicamente contengono più semi (plurispermi), si aprono per permettere ai semi di fuoriuscire. 2.FRUTTI CARNOSI→parenchima ricco d’acqua FRUTTI COMPOSTI: si originano da tanti fiori, derivano dalla fusione di molti ovari. Sono infatti costituiti da molti piccoli frutti. 1.FRUTTI SECCHI: gli strati hanno scarsi parenchimi e un contenuto di acqua piuttosto basso; il pericarpo può quindi essere duro, papiraceo o legnoso. Per la modalità di liberazione dei semi si distinguono in: a)Frutti secchi deiscenti→a maturità liberano i semi aprendosi spontaneamente, plurispermi, presentano delle zone di deiscenza sprovviste di fibra, con cellule sottili cellulosiche: • follicolo: frutto monocarpellare, deisce (a maturità si apre) lungo la linea di sutura del carpello (anice stellato) o sulla linea dorsale (magnolia). • legume o baccello: frutto monocarpellare, deisce in 2 valve su 2 linee opposte (sutura e dorso). È tipico della famiglia delle Leguminose (fagiolo, pisello, fava, ginestra). • capsula: deriva da un ovario pluricarpellare e presenta vari tipi di deiscenza (tabacco, lilium, tulipano, papavero) • siliqua e siliquetta: derivano da un ovario bicarpellare, sincarpico, pluri/monospermio. A maturità le silique si fendono ma non si separano completamente. Le due valve sono separate da un setto persistente, membranoso detto replo, su cui sono inseriti i semi. Il frutto si chiama siliquetta quando è isodiametrica (crucifere). b)Frutti secchi indeiscenti: presentano una parete completamente sclerificata, a maturità non liberano i semi che vengono dispersi ancora protetti dal pericarpo, monospermi. • achenio: frutto mono/pluricarpellare, con pericarpo membranoso, pergamenaceo o anche cuoioso, il seme è aderente al frutto, ma non saldato (Fagacee, Betulaceae, Composite); •samara: achenio con pericarpo membranoso con espansione alare laterale (frassino) o periferica (olmo); se il frutto è formato da due samare aderenti si chiama disamara (acero); • cariosside: bi-tri-carpellare, con pericarpo saldato all’episperma (tegumento esterno del seme), tipico delle Graminacee. • noce: pericarpo legnoso, mono/pluricarpellare (nocciolo), se ha dimensioni ridotte si chiama nucula. 2.FRUTTI CARNOSI Mesocarpo succoso e polposo (carnoso). Il seme è contenuto in un nocciolo legnoso che non è in grado di aprirsi spontaneamente (indeiscente). • Drupa: epicarpo sottile, mesocarpo carnoso, endocarpo legnoso; alcune drupe sono monospermie monocarpellari (pesco, susino, albicocco), pluricarpellari (olivo, noce), plurispermie pluricarpellari (caffè, pepe) Possono anche avere mesocarpo coriaceo (noce, mandorlo) o fibroso (cocco) • Bacca: epicarpo membranoso, mesocarpo ed endocarpo carnoso. (Uva, pomodoro, mirtillo, banana, avocado, peperone, melanzana) 26 • Esperidio: tipico frutto degli agrumi: bacca particolare con epicarpo sottile, colorato, ghiandolare, mesocarpo bianco e spugnoso, endocarpo settato formare spicchi. • Peponide: frutto delle Cucurbitacee costituito da una bacca rivestita da uno strato esterno non separabile dal frutto. Falsi frutti→La porzione edule(=commestibile) del frutto è formata da tessuti che non appartengono all’ovario, principalmente è il rivettacolo a contribuire. Es: Pomo: tipico di molte rosacee, quali la mela, la pera. La parte edule è costituita dal ricettacolo fiorale ingrossato. Il “frutto vero” è il torsolo: ovario infero pentacarpellare saldato con ricettacolo ipertrofizzato, endocarpo coriaceo, calice persistente. FRUTTI COMPOSTI Derivano dalla fecondazione di fiori compositi o infiorescenze, ovvero dalla fusione o dall’ evoluzione contemporanea di diversi ovari posti sullo stesso ricettacolo. Siconio: nei fichi, l’asse infiorenziale cavo e carnoso contiene veri frutti che sono piccoli acheni. Sorosio: falso frutto aggregato, derivante dal concettacolo dell'infiorescenza (massa carnosa). Nelle more e lamponi si ha l’unione di tante piccole drupe, separate da calice e corolla persistenti. Nella fragola i frutti sono le piccole nucole sulla superficie del ricettacolo ingrossato. Nell’ananas è tutto l’asse infiorescenziale a farsi carnoso. Importanza economico-farmaceutica dei frutti e dei semi: La maggior parte dei frutti è consumata allo stato fresco. Contribuiscono alla composizione della dieta con zuccheri, vitamine, minerali ecc. Frutti di impiego farmaceutico ed erboristico sono: .papaia (contiene la papaina enzima digestivo proteolitico), .arancia e agrumi, .capsula del papavero (alcaloidi dell’oppio), .anice nostrano .l’anice stellato .capsico .senna frutto. Anche i semi hanno impiego alimentare: .cereali (frumento, riso, mais, orzo, segale, avena, miglio, sorgo) . .Alcune piante danno semi per l’estrazione di oli di uso alimentare e farmaceutico: colza, semi di lino, di cotone, di ricino, di noce di cocco, di sesamo, di palma ... .Altri semi utilizzati: caffè, cacao, mandorlo dolce ed amaro, noce, nocciola, castagna. Di uso solo farmaceutico ricordiamo: i semi di Fava del Calabar (fisostigmina: miotizzante, antimiastenico, enterocinetico), i semi di Strofanto (strofantina, tipo digitalico, azione sul cuore), semi di Colchico (antigottosi ed antinevralgici), semi di Noce vomica (sostanze amare eupeptiche, stimolanti nervosi). Nella Farmacopea ufficiale Italiana (F.U.I.) sono riportati: • come frutti: senna, anice, capsico, cardo mariano, arancio amaro; • come semi: lino, ippocastano, psillio, noce vomica. 27