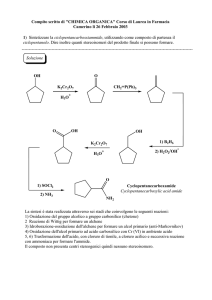
Il ciclo dell’acido citrico
Il ciclo dell’acido citrico
• Come visto in precedenza alcune cellule ricavano energia
(ATP) dalla fermentazione in particolare digerendo il
glucosio in assenza di ossigeno. Per molte cellule
eucariotiche e molti batteri la glicolisi è solo il primo step
della completa ossidazione del glucosio. Il piruvato
prodotto dalla glicolisi, invece di essere fermentato a alcol
etilico, acido lattico o altro è ulteriormente ossidato, in
presenza di ossigeno, fino a dare anidride carbonica e
acqua.
• In presenza di ossigeno possiamo distinguere 3 fasi del
catabolismo:
Formazione
dell’acetil
CoA da vari
substrati
Ossidazione
dell’acetil CoA
durante il ciclo
dell’acido citrico
Fosforilazione ossidativa
Il Coenzima A
Acido
pantotenico
Il CoA è un importante coenzima formato (cominciando da
destra) da una base purinica, l’adenina, il ribosio fosforilato in
posizione 2’ due fosfati, l’acido pantotenico (che è una
vitamina) e la cisteamina. Cisteamina e acido pantotenico
insieme formano la cosidetta panteteina.
Cisteamina
Coenzima A
• Il CoA ha la proprietà di formare tramite il
suo gruppo SH legami tioestere con gruppi
carbossilici. Si ricorda che il legame
tioestere ha caratteristiche peculiari. Si
tratta di un legame altamente energetico la
cui idrolisi comporta un ∆G°’ di – 31
Kcal/mole. Vediamone il perché strutturale
Legame tioestere
Rispetto all’estere il tioestere non ha una stabilizzazione per
risonanza. Questo comporta che la sua idrolisi libera più
energia: quella che nell’estere con l’ossigeno è la
stabilizzazione per risonanza (in celeste in figura)
Legame tioestere
• Ricordate, quindi, che come per l’ATP la
formazione di un legame altamente energetico
quale quello fosfoanidridico o tioesterico
comporta sempre che ci sia una grande quantità di
energia impiegata. Quando incontrate che si forma
ATP o un tioestere chiedetevi sempre come si sia
potuto formare, da quale processo chimico derivi
l’energia chimica necessaria alla formazione di
questi legami.
La formazione dell’acetilCoA dal
piruvato
Questa reazione, apparentemente semplice, è invece molto
complessa: come vedete viene decarbossilato l’acido piruvico
che è il prodotto della glicolisi. La sua decarbossilazione ad
acetile comporta l’uso di un complesso multienzimatico: la
piruvico deidrogenasi.
Piruvico deidrogenasi
• Questo complesso multienzimatico richiede
cinque coenzimi: il Coenzima A, la tiamina
pirofosfato, l’acido lipoico, il FAD e il NAD.
• Si ricorda che i coenzimi NAD e FAD sono i
coenzimi delle deidrogenasi e, quindi, si trovano
in reazioni di ossido-riduzione. In questo caso, il
gruppo chetonico dell’acido piruvico viene
ossidato a gruppo carbossilico acido con
contemporanea decarbossilazione del gruppo in
alfa del substrato piruvato
Il Coenzima A
L’Acido Lipoico
L’Acido lipoico è un acido ottanoico
con due gruppi SH separati da un
CH2. La sua caratteristica è di poter
formare legami tioestere e in più di
potersi ossidare nella sua forma
disolfuro (ponte –S—S-)
La tiamina pirofosfato (derivato
della vitamina B1)
Il gruppo reattivo della tiamina pirofosfato è il carbonio indicato in rosso:
l’idrogeno in rosso è un idrogeno acido. Il carbanione derivante dall’uscita del
protone è stabilizzato dall’azoto e lo zolfo con un concomitante effetto
elettron- attrattore dell’anello adiacente.
Decarbossilazione dell’acido piruvico
(Enzima 1, piruvico decarbossilasi)
• Durante questa prima fase della reazione
multienzimatica il piruvato si lega alla
tiamina pirofosfato. Il carbanione
dell’anello tiazolico della tiamina attacca il
carbonio chetonico del piruvato come si
vede nelle figure seguenti.
• Una volta legato, il
piruvato subisce una
decarbossilazione con
formazione di un
intermedio detto idrossietil
TPP che, se idrolizzato,
darebbe acetaldeide.
Quindi al coenzima TPP si
legano delle cosidette
aldeidi attivate. Questo
coenzima si ritrova anche
in altri processi che
vedremo in seguito.
Step 2: l’enzima diidrolipoiltransacetilasi.
• L’idrossietil-TPP trasferisce a questo punto l’aldeide attivata sull’acido
lipoico in forma ossidata che si riduce e forma un tioestere. Questo
secondo step è uno step di ossidazione: il gruppo aldeidico attivato,
infatti, viene ossidato a gruppo carbossilico acido che può reagire con
il gruppo SH dell’acido lipoico per dare un tioestere. I due elettroni
rimossi dall’aldeide durante l’ossidazione vengono trasferiti all’acido
lipoico ossidato che si riduce e, quindi, può legare il gruppo
carbossilico formando il tioestere.
• Dunque: avviene un’ossidazione del substrato con contemporanea
riduzione dell’acido lipoico; si forma un tioestere che per formarsi usa
l’energia derivante dall’ossidazione del substrato. Questa reazione è
molto efficiente perché oltre ad essere una ossidoriduzione anche
l’energia derivante dalla ossidazione viene recuperata sottoforma di
legame altamente energetico.
SCHEMA DI REAZIONE DELLA
PIRUVICO DIDROGENASI
Step 3 diidrolipoil deidrogenasi
• Lo step 3 che potete vedere nello schema precedente
comporta la riossidazione dell’acido lipoico che riforma il
ponte disolfuro, la contemporanea riduzione del FAD a
FADH2 e il trasferimento del tioestere all’acetil CoA.
• In sintesi il gruppo acetile che prima stava sull’acido
lipoico legato tramite un tioestere viene trasferito al CoA,
sempre legato al suo gruppo SH tramite un tioestere,
Contemporaneamente i gruppi SH dell’acido lipoico
vengono ossidati a ponte disolfuro e gli elettroni trasferiti
al FAD che si riduce. Il FAD fa parte di questo terzo
enzima.
Trasferimento degli elettroni al NAD
• Nell’ultima fase di questa complessa reazione gli elettroni
dal FADH2 passano al NAD che così si riduce a NADH.
• Ricordate che la riduzione del NAD da parte del FADH2 è
un evento che può avvenire se e solo se le concentrazioni
dei due reagenti lo permettono. Infatti guardando ai loro
potenziali standard sarebbe possibile solo la riduzione del
FAD da parte del NADH, ma cambiando le concentrazioni
il potenziale reale può essere diverso da quello standard. In
particolare il FAD in questa reazione risulata legato
all’enzima mentre il NAD è libero.
Schema generale del ciclo dell’acido
citrico
• Una volta che il piruvato derivante dalla glicolisi ma, come vedremo
anche da altri processi metabolici, è stato trasformato in acetil-CoA,
subisce una serie di reazioni che nell’insieme vengono dette “ciclo
dell’acido citrico” o “ciclo di Krebs” nel quale, in ultimo, il gruppo
acetile (che anch’esso può avere derivazioni diverse che quella dal
piruvato) viene ossidato ad anidride carbonica ed acqua.
• Un buon esercizio per voi studenti è di scrivere tutte le reazioni che
adesso andremo ad esaminare colorando in rosso i carboni del gruppo
acetile legato al CoA per vedere che fine fanno durante il ciclo.
• Il ciclo viene detto dell’acido citrico perché la prima reazione che
subisce il gruppo acetile è di condensazione con l’ossalacetato per
dare, appunto, il citrato come si vede nella figura successiva.
La reazione ha un ∆G negativo derivante dalla contemporanea
condensazione e idrolisi del legame tioestere altamente energetico.
L’idrolisi del legame tioestere rende la reazione fortemente spostata verso
la formazione del citrato e forza il gruppo acetile a entrare nel suo ciclo di
ossidazione
L’enzima citrato sintasi
Le figure riportano la struttura cristallografica dell’enzima citrato sintasi. La “a” è l’enzima
libero, la “b” è l’enzima legato al substrato (in giallo l’ossalacetato in rosso un analogo del CoA).
Si noti il cambiamento conformazionale dell’enzima dopo che lega il substrato. Quando l’enzima
lega il primo substrato che è l’ossalacetato si ha un grande cambiamento conformazionale che
porta alla creazione del sito di legame dell’acetil-CoA. Una volta che si è formato l’intermedio
citroilCoa si ha un nuovo cambiamento conformazionale che porta un residuo di aspartato in una
posizione cruciale che permette l’idrolisi del tioestere.
Dal citrato all’isocitrato tramite
l’enzima aconitasi
Ricordate che l’aconitasi è una proteina Ferro-Zolfo. Questo
tipo di proteine le rincontreremo anche in seguito. Nella figura
che segue trovate la struttura del cosidetto CLUSTER ferro
zolfo dell’enzima aconitasi. Domandatevi a quale classe di
enzimi appartiene l’aconitasi
L’ossidazione dell’isocitrato ad α-chetoglutarato
tramite la isocitrato deidrogenasi
Notate che gli elettroni derivanti dall’ossidazione vengono
trasferiti al NAD o al NADP (ripassate le strutture dei due
coenzimi) e che la reazione ha una variazione di energia libera
standard molto negativa. Notate che qui la decarbossilazione è
più facile rispetto a quella del piruvato, infatti dopo l’ossidazione
dell’isocitrato il carbonio interessato diviene chetonico da
alcolico e il gruppo carbossilico che poi esce come anidride
carbonica diviene un gruppo carbossilico in beta. Vi ricordo che i
beta cheto-acidi decarbossilano spontaneamente
La reazione dell’α-cheto glutarato
deidrogenasi
Questa reazione è del tuto simile a quella
della piruvato deidrogenasi. Viene catalizzata
da un sistema multienzimatico che ha
esattamente gli stessi coenzimi di quelli visti
per la decarbossilazione/ossidazione del
piruvato.
La formazione del primo GTP
Il GTP è analogo all’ATP e contiene i legami altamente
energetici utili poi per moltissimi scopi. In questa
reazione il gruppo fosfato libero si lega al GDP usando
l’energia derivante dall’idrolisi del legame altamente
energetico presente come tioestere sul succinil CoA. Vi
ricordo che il GTP è un nucleotide formato da Guanina,
ribosio e tre gruppi fosfato
La succinato deidrogenasi produce
FADH2 e fumarato
La fumarasi produce malato
La malato deidrogenasi produce NADH e
ossalacetato, chiudendo il ciclo
Schema riassuntivo
Resa
• Come avete potuto studiare durante un
intero ciclo nel quale l’acetilCoa si
condensa con l’ossalacetato e si ossida a
CO2, si hanno in uscita: due molecole di
anidride carbonica, un GTP (che poi è un
ATP), 3 NADH e un FADH2.
• Cosa vuol dire questo?
•
•
Vi ricordo che negli organismi anaerobi o in condizione anaerobie, dopo la
glicolisi il piruvato viene fermentato. Questo accade anche nei nostri muscoli
scheletrici in carenza di ossigeno dove il iruvato viene ridotto a lattato con
contemporanea ossidazione del NADH. Perché avviene ciò? Il NADH e il
FADH2 devono essere in continuazione riossidati per dar modo ai vari cicli
ossidativi di andare avanti. Se non fossero efficientemente riossidati ben presto
non si avrebbe più nella cellula neanche una molecola di coenzima ossidato
disponibile per le attività di ossidazione proprie del metabolismo.
Gli organismi aerobi in condizioni normali riossidano questi cofattori
trasportando gli elettroni sottratti all’ossigeno molecolare che con quattro
elettroni può ridursi ad acqua. Questo processo, che nelle cellule eucariotiche
avviene nei mitocondri, è molto efficiente. Il trasporto degli elettroni avviene a
step durante i quali lo stesso trasporto è accoppiato, tramite un meccanismo
che vedremo in seguito, alla sintesi di ATP. Per ogni NADH che viene
ossidato si generano nel trasporto degli elettroni 3 ATP e 2 ne provengono
invece dal FADH2. Quindi si può facilmente calcolare che un ciclo di Krebs
produce quando è efficiente 12 ATP, per ciascun acetil-CoA che entra, non
usando nessun legame altamente energetico per attivare il substrato.
• La tabella che segue vi illustra proprio la resa in ATP dell’intero
processo di ossidazione di una molecola di glucosio, a partire dalla sua
attivazione a glucosio-6-fosfato a finire alla produzione di CO2 nel
ciclo di Krebs. Vi faccio notare che in tabella a partire dalla
gliceraldeide-3-fosfato tutte le rese sono raddoppiate. Da una molecola
di glucosio, infatti, a 6 atomi di carbonio, si producono due molecole
di gliceraldeide e conseguentemente due molecole di Acetil-CoA che
si ossida nel ciclo dell’acido citrico.
• Noterete anche che per ogni NADH vengono calcolate 2,5 molecole di
ATP che si formano durante la riossidazione e dal FADH2 1,5. Questo
perché l’autore tiene conto dell’efficienza media del processo. Questo
aspetto lo vedremo bene studiando la catena di trasporto degli elettroni.
Altre caratteristiche del ciclo di
Krebs.
•
•
•
•
Il ciclo appena visto è un ciclo che serve sì a ossidare l’acetile legato al CoA,
ma anche a produrre importanti intermedi catabolici. Un esempio per tutti.
Prendiamo ad esempio l’ossalacetato che viene prodotto dall’ossidazione del
malato:
Si tratta di una molecola organica a quattro atomi di carbonio che contiene due
gruppi carbossilici acidi. Se ci pensate bene l’acido ossalacetico somiglia
molto all’acido aspartico, uno dei venti aminoacidi che abbiamo studiato. Se
solo al gruppo chetonico sostituite un gruppo aminico
Vedremo in seguito che l’ossalacetato può essere trasformato in aspartato da
un processo di transaminazione. Lo stesso dicasi per l’α−chetoglutarato che
può essere transaminato a glutammato. Isnomma gli intermedi del ciclo di
Krebs sono importanti nel nostro metabolismo.
Quando gli intermedi del ciclo vengono rimossi per altri scopi essi stessi
possono essere reintegrati attarverso reazioni anaplerotiche e per mezzo di
questi meccanismi le concentrazioni degli intermedi del ciclo romangono
pressochè costanti. Alcune reazioni anaplerotiche sono mostrate nella tabella
che segue la figura
• REAZIONI DI
CARBOSSILAZIONE: Esse
dipendono da una vitamina, la
Biotina che solitamente è legata
all’enzima che catalizza la
reazione attraverso il gruppo εaminico di una lisina. La
Biotina è capace di legare lo
ione bicarbonato attivandolo
poi per il trasferimento sul
substrato da carbossilare.
• Il legame della molecola
inorganica alla biotina ha
bisogno di energia che viene
fornita attraverso l’idrolisi di
ATP ad ADP + Pi