09/10/16
LA CELLULA VEGETALE
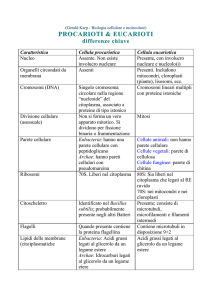
Come tutte le cellule eucariotiche, la cellula vegetale
racchiude il suo genoma all’interno del NUCLEO. Il
genoma è l’insieme dei geni. Ogni gene contiene
l’informazione per produrre una proteina (tutti gli enzimi
sono proteine).
La cellula possiede anche un ricco sistema di
endomembrane e diversi tipi di organelli semiautonomi
e non. Alcune di queste dotazioni sono comuni a tutti
gli eucarioti, altre sono caratteristiche.
•
Solo le cellule vegetali possiedono:
•
Organelli semiautonomi detti cloroplasti che permettono la
fotosintesi.
I plasmodesmi (che collegano le cellule attraverso la parete
cellulare e contengono RE).
Apparati del Golgi liberi in numero di diverse centinaia.
Vacuoli di diverso tipo.
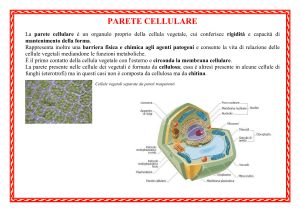
La parete cellulare.
•
•
•
•
1
09/10/16
•
La cellula è delimitata e divisa in compartimenti ed organelli da membrane
biologiche composte da un doppio strato lipidico in cui sono inseriti vari tipi di
proteine. Le membrane sono impermeabili alla soluzione acquosa che le
circonda ma rimangono fluide ed i loro componenti possono spostarsi
secondo
necessità
(MOSAICO
FLUIDO).
Specifiche
proteine
transmembrana, formano PORI e CANALI con la capacità di controllare il
passaggio di diverse molecole e quindi assicurare lo scambio controllato di
molecole fra i diversi compartimenti ed organelli della cellula. Un esempio
sono i pori nucleari, composti da molte proteine e molto grandi. Essi fanno
uscire gli RNA messaggeri ma lasciano entrare solo le proteine con specifici
segnali.
GLI ORGANELLI SEMIAUTONOMI
Si dicono tali poiché contengono geni propri, necessari alla loro replicazione
“semiautonoma”. Sono MITOCONDRI, in cui si ha produzione di energia (ATP)
in seguito alla respirazione, e PLASTIDI.
I plastidi hanno la possibilità di acquistare molte forme. Nelle cellule
meristematiche indifferenziate il plastidio altrettanto indifferenziato prende il
nome di proplastidio. Esso può, sotto il controllo di vari stimoli (luce, ormoni),
differenziarsi in CLOROPLASTO (fotosintetico), CROMOPLASTO (funzione
vessillare con accumulo di pigmenti), AMILOPLASTO (riserva di amido),
IALOPLASTO (riserva di lipidi) o EZIOPLASTO (dedifferenziato per assenza di
luce). La forma che determina l’AUTOTROFIA è il cloroplasto ed in esso si
svolge la fotosintesi. Questo processo porta alla cattura dell’energia luminosa
attraverso 2 complessi fotosistemi (PSI e PSII) che la trasformano in energia
chimica: ATP e NADPH. I fotosistemi e l’ATP sintetasi sono situati in modo
strategico sulle membrane interne dei cloroplasti, dette TILACOIDI. Queste
membrane sono organizzate in GRANA. Sulle membrane avviene la cattura
della luce con consumo di H2O e produzione di O2, nello stroma del cloroplasto
invece si fissa la CO2 in composti organici.
2
09/10/16
INTERCONVERSIONE PLASTIDIALE
Nei cloroplasti avviene la fotosintesi
3
09/10/16
A cosa servono
acqua e la luce.
La cellula vegetale ha
caratteristiche uniche
che la distinguono da
quella animale:
I plastidi (autotrofia);
Plasmodesmi
Complesso vacuolare
Golgi organizzato in
dittiosomi;
Parete cellulare.
Si riferiscono al
sistema di secrezione
4
09/10/16
Fatta eccezione per le proteine destinate al citosol, che vengono
semplicemente rilasciate nel citoplasma dopo la sintesi, tutte le proteine
contengono sequenze aminoacidiche (segnali) che le dirigono verso la loro
destinazione finale: RE, nucleo, mitocondri, plastidi, altri microcorpi
Il “sistema di secrezione”
o “sistema di endomembrane”
caratterizza tutte le cellule eucarioiche
5
09/10/16
Il 17% circa del genoma vegetale
è dedicato al mantenimento di questo sistema
Nella sua compartimentazione appare
più ricco e complesso che in altri eucarioti
Ma la sua organizzazione è simile in tutti gli eucarioti:
Reticolo Endoplasmico, Apparato di Golgi,
Trans Golgi Network, Vacuoli/lisosoma, Endosomi
Plasmalemma
apoplasto
Golgi
Reticolo
Endoplasmatico
Trans
Golgi
Network
Vacuoli
6
09/10/16
Il Traffico di membrana si
svolge in modo anterogrado e
retrogrado, la polarità deriva
dalla porta di ingresso delle
proteine e dal progressivo
arricchimento delle
modificazioni post-traduzionali:
REGolgiTGNVacuoli/PM
De Caroli et al. Plant Journal 2011
Le proteine entrano nel
sistema attraverso la
traslocazione
Nei compartimenti
endomembranosi
subiscono numerose
modificazioni post
traduzionali necessarie
al controllo di qualità ed
al funzionamento
7
09/10/16
Inizio sintesi su ribosomi liberi
Riconoscimento primi aa
(N-ter = segnale di traslocazione)
Particella Riconoscimento Segnale
(SRP)
Traslocazione
Tramite canale proteico:
Il traslocone
Maturazione:
Primo evento
taglio segnale di traslocazione)
N-glicosilazione
(Glicosiltransferasi lato citosolico e luminale)
Ripegamento
Ponti disolfuro
(servono “proteine che aiutano”
Le ciaperonine come
BiP)
Le proteine entrano nel sistema
attraverso la traslocazione.
Una volta nel “lume”
Possono restare nel RE o
essere esportate nel Golgi
da qui andare a:
Vacuoli/tonoplasto
Parete/plasmalemma
NO:
plastidi, mitocondri, nucleo
Nel caso di MPT errate, vengono
retro-traslocate al citosol e
distrutte dal “proteasoma” oppure
smistate ai vacuoli per digestione
e riciclo.
8
09/10/16
Nel Reticolo
Endoplasmico
(RE) sono
riconoscibili
oltre 16 diversi
domini
I più evidenti:
L’involucro nucleare
Il RE liscio (REL)
Il RE rugoso (RER)
I plasmodesmi
I corpi proteici
Gli ERES (ER export sites)
I siti di contatto con:
il citoscheletro,
i plastidi,
i mitocondri
ect.
FUNZIONI DEL RE
Il compartimento può accumulare, oltre che lipidi, anche proteine
di riserva. Infatti mentre nella maggior parte dei casi questa funzione è
svolta dai vacuoli (specie nelle dicotiledoni), in molte erbacee e cereali,
proteine di riserva classificabili come prolamine vengono accumulate
proprio nel RE in “corpi proteici”. Ricordiamoci che i cereali sfamano il
mondo intero.
9
09/10/16
Strutture
fusiformi
estremamente
mobili per
l’accumulo di
proteine ci
anticipano
quanto il RE
sia dinamico
sGFPKDEL, un semplice
marcatore solubile del RE
10
09/10/16
L’apparato di Golgi dei vegetali è organizzato in dittiosomi.
A questo punto i sistemi vegetali, animali e fungini
divergono.
Ogni dittiosoma è una unità funzionale
11
09/10/16
I dittiosomi sono peculiari dei vegetali ma la frammentazione del Golgi avviene
anche durante la divisione cellulare animale o durante il differenziamento dei
neuroni…
I dittiosomi scambiano materiale con gli ERES e
con il TGN sia tramite vescicole che tramite
maturazione delle membrane.
12
09/10/16
FUNZIONI DELL’APPARATO DI GOLGI
Sintesi di diversi tipi di polisaccaridi come pectine ed
emicellulose, con ruolo essenziale nella costituzione della
parete cellulare;
processamento delle catene oligosaccaridiche N-legate;
O-glicosilazione delle proteine, che consiste nell'aggiunta di
zuccheri al gruppo ossidrilico (-OH) di specifici aminoacidi;
Fosforilazione: un elemento di regolazione importante
dell’attività delle proteine (integrato nella trasmissione dei
segnali e nella risposta agli stimoli);
Smistamento delle proteine.
Ulteriore
complessità si
riscontra nei
compartimenti postGolgi:
Pre-vacuoli e
vacuoli.
13
09/10/16
La sintesi della parete cellulare dipende da
questo complesso sistema.
Struttura parete
14
09/10/16
La parete cellulare si modifica
La PARETE CELLULARE, deve molto al Golgi
nonostante si trovi all’esterno della cellula (apoplasto) può essere definita un organulo tipico della cellula
vegetale
I primi passi nella formazione della parete corrispondono alla divisione cellulare.
La divisione della cellula si realizza mediante la formazione di un setto di separazione la cui porzione centrale è
detta lamella mediana (priva di cellulosa). Le prime fasi della sua formazione corrispondono alla formazione del
così detto FRAGMOPLASTO, cui contribuiscono, microtubuli e dittiosomi.
15
09/10/16
RE e Golgi sintetizzano e smistano tutto il materiale della
componente di matrice della parete cellulare.
Golgi
RE
La PARETE CELLULARE,
Ultimata la divisione cellulare, si forma la PARETE PRIMARIA
con tutti i componenti di base:
COMPONENTE FIBRILLARE
Cellulosa (sintetizzata sul plasmalemma)
MATRICE
Sostanze pectiche ed emicellulose (sintetizzate nel Golgi)
Proteine (sintetizzate sul reticolo endoplasmico)
16
09/10/16
A ridosso della membrana ogni cellula costituisce la PARETE
PRIMARIA. Essa è destinata ad accrescersi anche notevolmente
per distensione (tessitura dispersa della cellulosa). Qui la matrice
è preponderante.
Alla fine della crescita cellulare, si forma uno strato all’interno
della parete primaria, detto PARETE SECONDARIA (tessitura
parallela della cellulosa: fibrosa, ad elica, anulare). Qui la parte
fibrillare è preponderante.
Il segreto della resistenza meccanica è
nella successione di strati con una diversa
TESSITURA della CELLULOSA
17
09/10/16
La parete non è compatta e quindi, dopo la sua completa
formazione, può subire l’incrostazione con :
LIGNINA (LIGNIFICAZIONE),
FENOLI e PIGMENTI (PIGMENTAZIONE),
CARBONATI (MINERALIZZAZIONE),
CUTINA (CUTINIZZAZIONE)
o SUGHERO (SUBERIFICAZIONE).
Le funzioni principali della parete sono:
regolare la velocità di crescita della cellula e determinarne la forma
e la grandezza;
controbilanciare fisicamente la pressione osmotica;
regolare il movimento di ioni, acqua e piccole molecole;
costituire un'imponente biomassa;
regolare l'espressione genica, controllando la crescita e il
differenziamento cellulare.
18
09/10/16
La composizione chimica della parete cellulare è differente nelle
Dicotiledoni e nelle Monocotiledoni (specialmente la parete delle
Graminacee presenta caratteristiche peculiari).
Essenzialmente si compone di:
Cellulosa
Sostanze pectiche
Emicellulose
Proteine strutturali ed enzimatiche
Caratteristica comune a tutte le piante superiori è la presenza
della componente fibrillare rappresentata da microfibrille di
cellulosa, costituite dall'associazione di lunghe catene glucaniche
nelle quali i residui glucosidici sono tenuti insieme da legami di
tipo b-(1-4).
Ciascuna catena glucanica può essere lunga diverse migliaia di
residui glucosidici.
19
09/10/16
Solo il 10% del peso secco della parete delle Graminacee è
rappresentato dalle sostanze pectiche che, invece, nelle
Dicotiledoni, rappresentano un terzo del peso secco della parete.
Svolgono un ruolo strutturale di fondamentale importanza nella
parete; infatti le caratteristiche plastiche della parete dipendono
appunto dalla presenza di pectine.
Le emicellulose nelle Graminacee rappresentano il 50% della
parete. Esse sono rappresentate da xilani la cui struttura base è
costituita da una catena lineare formata da residui di D-xilosio
tenuti insieme da legami di tipo b-(1-4).
20
09/10/16
I polimeri di matrice vengono biosintetizzati a livello
dell'apparato di Golgi
ad opera di polisaccaride sintasi,
utilizzando i nucleosidi difosfato zuccheri attivati.
I polimeri neosintetizzati si muovono attraverso le cisterne
dei dittiosomi e arrivano al plasmalemma tramite vescicole.
La sintesi della cellulosa si realizza a livello di
plasmalemma, ad opera del complesso cellulosa
sintasi che utilizza UDP-glucosio come substrato.
21
09/10/16
Più si manipola la cellulosa e più
si degrada la struttura cristallina.
Ciò è molto rilevante ai fini del
biodeterioramento.
22
09/10/16
La tessitura delle fibrille di cellulosa è DISPERSA
nella PARETE PRIMARIA
mentre è PARALLELA
nella PARETE SECONDARIA.
In quest’ultima vi sono ben 3 strati di fibrille
parallele ma con orientamento diverso da strato a
strato.
La disposizione parallela aumenta la compattezza e
riduce quindi lo spazio libero per modificazioni
secondarie come la lignificazione.
La forza meccanica aumenta comunque
notevolmente grazie a questa successione di strati.
Il segreto della resistenza meccanica è
nella successione di strati con una diversa
TESSITURA della CELLULOSA
23
09/10/16
Un'altra importante componente della parete cellulare
è rappresentata dalle proteine strutturali ed
enzimatiche che possono costituire fino al 10% del
peso secco della parete.
24
09/10/16
Gli ENZIMI di parete e le proteine strutturali
arrivano nella PARETE CELLULARE per
secrezione ma entrano nel sistema già nel RE.
I POLISACCARIDI di matrice sono sintetizzati nel Golgi,
attingendo al pool di NUCLEOTIDI DIFOSFATO-ZUCCHERI nel
citoplasma
dal Golgi vengono secreti nella PARETE CELLULARE attraverso
vescicole.
La CELLULOSA viene sintetizzata sulla MEMBRANA
PLASMATICA da complessi enzimatici a rosetta e polimerizza
direttamente nella parete.
Anche i monolignoli vengono secreti ma la
LIGNINA in quanto tale polimerizza direttamente
nella parete cellulare.
Troveremo tanta più lignina quanto minore è il
contenuto di cellulosa.
25
09/10/16
Lamella mediana
Sost. Pectiche
Emicellulose
proteine
Parete primaria
Cellulosa
Sost. Pectiche
Emicellulose
proteine
Parete secondaria
Cellulosa
(tessitura parallela =fitta)
Sost. Pectiche
Emicellulose
Proteine
26
09/10/16
INTRUSIONE DI LIGNINA
LIGNIFICAZIONE
27