Livelli di organizzazione degli
organismi vegetali
Nel corso dell’evoluzione forme di organizzazione
sempre più complesse:
11. Istologia vegetale
• Organizzazione dei vegetali
• Tessuti e pseudotessuti
1
• Protofite
• Aggregati cellulari
• Tallofite
- tallo sifonale
polienergide
- tallo pluricellulare
- tallo cormoide
• Cormofite
2
Euglena gracilis
Protofite
Da Raven et al.
Organismi unicellulari microscopici
cellula = organismo
9 procarioti
9 alghe mobili, es. Chlamydomonas, Euglena
9 alghe immobili (org. coccale), es. Chlorella, diatomee
9 lieviti (funghi)
Da Ferroni et al. 2004
Da Lüttge et al.
Cyclotella (diatomea)
Chlamydomonas
3
Da Lüttge et al.
Chlorella vulgaris
4
1
Aggregati cellulari
Alcuni organismi unicellulari tendono ad associarsi tra
loro in colonie
associazioni POST-GENITE
Gloeocapsa
9 procarioti (cianobatteri, es. Nostoc, Gloecapsa)
9 alghe mobili (es. Gonium, Pandorina)
9 alghe immobili (es. Scenedesmus)
Da Lüttge et al.
5
• La formazione delle colonie apre la strada verso la
pluricellularità
• CENOBIO = colonia tipica delle alghe verdi volvocali
(Pandorina, Eudorina). Massima espressione in
Volvox.
Le cellule restano
collegate da ponti
citoplasmatici e compare
una primitiva
suddivisione del lavoro
tra cellule vegetative e
cellule riproduttive. 7
Scenedesmus
6
Tallofite
La mitosi cessa di essere necessariamente legata alla
riproduzione:
per mitosi l’organismo cresce = aumenta di dimensioni.
TALLO
corpo di vegetale in cui non siano differenziati veri
organi (radici, fusto e foglie)
8
Da Lüttge et al.
2
9 APOCIZIO: le divisioni nucleari non sono seguite da
citodieresi
Caso 1: alla mitosi NON segue la citodieresi
- tallo unicellulare = organizzazione sifonale
(es. Valonia, Caulerpa, Acetabularia, Vaucheria)
- tallo pluricellulare = più cellule polienergidiche,
organizzazione sifonocladale (es. Cladophora)
- ife non settate dei funghi inferiori
- nelle piante superiori, i tubi laticiferi delle euphorbiacee
Formazione di cellule PLURINUCLEATE
ENERGIDE
9 nucleo circondato dalla porzione di citoplasma sulla
quale esercita la sua funzione coordinatrice
9 a differenza della cellula, è un’entità solo funzionale
(non morfologica)
9 la cellula plurinucleata è un’entità polienergidica /
cenocitica
Nelle piante superiori c’è anche la possibilità del
9 SINCIZIO: fusione di cellule uninucleate
Es. tubi laticiferi delle papaveracee.
9
Da Lüttge et al.
10
Caso 2: alla divisione del nucleo segue la citodieresi
polienergide
Associazione CONGENITA di cellule:
formazione di un tallo pluricellulare
• talli filamentosi (es. Spirogyra, Zygnema)
• talli laminari (es. Ulva)
• talli complessi (es. Gracilaria, Laminaria, Chara)
• talli cormoidi (es. briofite = muschi ed epatiche)
Vaucheria
Cladophora
Da Nultsch.
11
12
3
Tallo filamentoso
Tallo laminare
Tallo complesso di alga bruna
Tallo cormoide di muschio
Ulva lactuca
Zygnema
Tallo complesso
di alga rossa
Da Nultsch
Furcellaria fastigiata
13
Da Nultsch.
Da Lüttge et al.
14
Cormofite
Le cellule che costituiscono i talli più complessi possono
formare degli agglomerati compatti:
La conquista dell’ambiente terrestre comporta il massimo livello
di specializzazione per risolvere problemi di:
- plectenchima (alghe rosse)
- ifenchima (funghi superiori)
ife
Da Nultsch.
- economia dell’acqua, l’atmosfera sottrae acqua alla pianta
- nutrizione, acqua e sali minerali sono disponibili nel suolo
- sostegno, manca la spinta dovuta all’acqua
- riproduzione, progressivo svincolarsi dalla dipendenza di un
ambiente liquido
CORMO = radici + fusto + foglie
PSEUDOTESSUTI
Non presentano le caratteristiche dei tessuti veri (per es. possono
derivare da cellule che si dividevano in una sola direzione,
15
generando filamenti che poi si accostano tra loro).
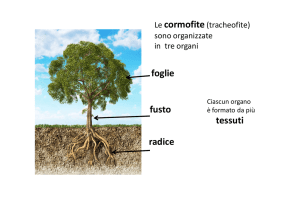
Cormofite = piante con veri organi e veri tessuti
• Pteridofite (felci, equiseti…)
• Spermatofite (gimnosperme + angiosperme)
16
4
CORMO
Tessuti veri
Aggregati congeniti di cellule che svolgono la stessa funzione.
Germoglio
Caratteristiche:
• derivati da cellule che si dividono secondo le 3 direzioni
dello spazio
• fragmoplasto e lamella mediana
• plasmodesmi e punteggiature
• specializzazione morfo-funzionale
Le cormofite sono
normalmente indicate
come tracheofite
o piante vascolari
• spazi intercellulari
Da Lüttge et al. 17
Tessuti veri e pseudotessuti
TESSUTI VERI
PSEUDOTESSUTI
1.
cellule madri che si dividono
secondo 3 direzioni
1.
2.
fragmoplasto e formazione
della lamella mediana
plasmodesmi e
punteggiature
2.
specializzazioni morfofunzionali caratteristiche del
tessuto
spazi intercellulari
(circolazione dei gas)
4.
18
Nei tessuti veri gli spazi intercellulari assicurano gli scambi
gassosi grazie alla circolazione dell’aria tra le cellule.
Origine durante la formazione del tessuto :
3.
4.
5.
3.
5.
cellule madri che si dividono
anche secondo una sola
direzione (es. ife)
anche altri tipi di citodieresi
(es. ficoplasto nelle alghe)
anche altri tipi di giunzione
tra cellule (es. giunzioni
alghe rosse)
livello di specializzazione
normalmente più basso
9 schizogena: le cellule si sono reciprocamente
allontanate, digerendo la lamella mediana che le
cementava
9 lisigena: alcune cellule, comprese le pareti, sono state
digerite
9 rexigena: alcune cellule sono state distrutte per
lacerazione o stiramento
assenza di veri spazi
intercellulari
19
20
5
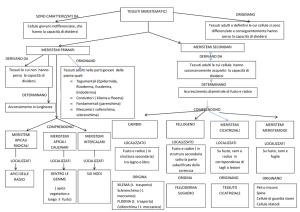
Differenziamento
CORMO
sdifferenziamento
Tessuti
MERISTEMATICI
(giovanili o formatori)
Tessuti
ADULTI
(definitivi)
Cellula
iniziale
IDIOBLASTI
Cellula
derivata
differenziamento
Cellula
adulta
Differenziamento = insieme di trasformazioni attraverso le
quali una cellula acquisisce le caratteristiche necessarie per
svolgere una specifica funzione.
Idioblasti = cellule adulte con caratteri differenti rispetto a
quelle del tessuto in cui si trovano (es. cellule ad essenza,
cellule a tannini)
21
22
Tessuti meristematici
Le piante conservano per tutta la loro vita cellule
capaci di dividersi (accrescimento indefinito).
12. Tessuti meristematici e
tessuti adulti
MERISTEMI
regioni della pianta costituite
da tessuti meristematici
• tessuti meristematici
• classificazione dei tessuti adulti
• tessuti parenchimatici
1. primari
2. secondari
23
24
6
Meristemi primari
•
9
9
9
9
• I meristemi primari comprendono le cellule iniziali e le loro
derivate che andranno incontro al differenziamento.
Tipiche cellule giovanili:
cellule piccole e isodiametriche
provacuoli
lamella mediana
proplastidi
• I tessuti adulti originati dai meristemi primari sono detti a
loro volta PRIMARI.
• Pteridofite e angiosperme monocotiledoni presentano solo
meristemi primari (no crescita secondaria).
• Comprendono i meristemi:
1. APICALI: apice di fusto e radice
2. INTERCALARI: interposti tra tessuti adulti
• Già presenti nell’embrione e attivi per tutta
la vita della pianta
• Responsabili dell’accrescimento
PRIMARIO longitudinale
25
26
Da Gerola.
Meristemi apicali
Meristemi intercalari
I meristemi intercalari (o
residui) si localizzano a livello
dei nodi e cooperano alla
crescita in lunghezza nei fusti
di monocotiledoni come
frumento, mais, canna…
Costituiscono punti di
debolezza per il fusto.
Da Nultsch.
27
Da Lüttge et al. Mod.
28
7
Meristemi secondari
Meristemi secondari cambiali
• Responsabili dell’accrescimento secondario in diametro
nelle gimnosperme e nelle angiosperme dicotiledoni:
originano i tessuti adulti SECONDARI.
• Derivano da cellule adulte che hanno riacquistato la
capacità di dividersi (sdifferenziamento): cellule più
grandi, provviste di parete primaria, sistema vacuolare
sviluppato.
• Comprendono:
1. meristemi CAMBIALI
2. meristemoidi: cellule che ritornano allo stato
embrionale per originare peli pluricellulari e apparati
stomatici.
3. meristemi avventizi: cellule che si sdifferenziano
per originare radici avventizie e tessuti cicatriziali.
• Comprendono:
1. cambio cribro-vascolare (cambio)
2. cambio subero-fellodermico (fellogeno)
• I cambi hanno la forma di un cilindro cavo che percorre
radici e fusti legnosi.
• Capacità di produrre cellule diverse sul lato esterno e su
quello interno del cilindro: attività DIPLEURICA.
29
Attività dipleurica dei cambi
Cambio cribro-vascolare
Cellule
del libro
1
Cribrovascolare
Cellule
del legno
INTERNO
ESTERNO
Cambio subero-fellodermico
(fellogeno)
30
Sughero
2
Felloderma
Fellogeno
31
32
8
Cellula
iniziale
Tessuti adulti
Le cellule derivate dai meristemi primari e secondari vanno
incontro al differenziamento = acquisiscono le
caratteristiche morfologiche/biochimiche fondamentali
per lo svolgimento di una determinata funzione.
differenziamento
sdifferenziamento
Nelle piante alcuni
processi di
differenziamento
conducono alla morte
della cellula
Cellula
adulta
•
•
•
•
Cellule vive o morte a maturità
Tipo di plastidio
Modificazioni parietali
Contenuti del vacuolo
33
34
Da Mauseth.
Classificazione dei tessuti adulti
•
•
•
•
•
Tessuti parenchimatici
• Struttura tissutale fondamentale
• Origine primaria o secondaria
• Tessuti plastici: possono assolvere più funzioni o
anche sdifferenziarsi
Parenchimatici = struttura fondamentale della pianta
Meccanici = sostegno
Tegumentali = protezione
Conduttori = trasporto
Segregatori = esclusione/secrezione di sostanze
Per ogni categoria esistono vari sottotipi con caratteristiche
morfo-funzionali specifiche.
Vari tipi di tessuti possono combinarsi per formare
complessi tissutali (legno, libro).
35
• Caratteristiche comuni:
9 sempre cellule vive
9 intensa attività metabolica
9 parete sottile (primaria)
9 ampi vacuoli
9 forma prismatica
9 spazi intercellulari
36
9
Parenchima clorofilliano (clorenchima)
Classificazione dei tessuti parenchimatici sulla base della
funzione vegetativa prevalente
• funzione fotosintetica
• tutte le parti verdi (foglie, fusti erbacei, frutti immaturi)
• origine prevalentemente primaria
• forma delle cellule: cilindriche, lobate, tondeggianti
Tessuti parenchimatici
Parenchima CLOROFILLIANO
Parenchima di RISERVA
Parenchima ACQUIFERO
Parenchima AERIFERO
Parenchima CONDUTTORE
Parenchima di TRASPORTO
Foglia di Arum
italicum, sezione
trasversale
37
Parenchima di riserva
Parenchima acquifero
• funzione di riserva
• midollo dei fusti, rizomi, bulbi, tuberi, radici, frutti, semi
• origine primaria o secondaria
• riserve accumulate in:
- amiloplasti (amido)
- vacuolo (proteine, fruttani, saccarosio)
- corpi lipidici (olii)
- parete (emicellulose nei semi di palme)
• funzione di riserva di acqua - piante succulente
• origine primaria o secondaria
• mucillagini idrofile contenute nei vacuoli (lento rilascio
d’acqua), parete sottile e flessibile
• gomme in piante non succulente
Parenchima amilifero in
tubero di patata
Cellule turgide del parenchima
acquifero
39
Da http://www.sbs.utexas.edu/mauseth/researchoncacti
38
Condizioni di stress idrico
40
10
Parenchima aerifero (aerenchima)
Parenchima conduttore
• circolazione dell’aria (scambi gassosi)
• piante acquatiche (idrofite)
• origine generalmente primaria
• le cellule si dispongono in catenelle/trabecole individuando dei
canali aeriferi
• altre funzioni: riserva, fotosintesi, galleggiamento
• assicura il trasporto radiale in fusti e radici in struttura
secondaria = raggi midollari
• cellule allungate in senso radiale
• origine secondaria
Trasporto
radiale
Trasporto
assiale
41
Picciolo di Arum italicum
42
Fusto di Potamogeton
Parenchima di trasporto
• associato agli elementi di
conduzione
• cellule transfer con
introflessioni della parete
cellulare: aumento della
superficie di scambio
• molti mitocondri: energia per i
processi di trasporto
• origine primaria o secondaria
13. Tessuti adulti
• tessuti meccanici
• fibre tessili
Da Lüttge et al.
43
44
11
La funzionalità del tessuto meccanico non deriva da
speciali attitudini del protoplasto, ma dalle caratteristiche
della parete cellulare
Tessuti meccanici
Le piante terrestri sono
sottoposte a:
• forze di pressione (peso della
chioma)
• forze di trazione (oscillazioni
dovute al vento)
Caratteristiche comuni ai tessuti meccanici:
• spazi intercellulari generalmente limitati
• ispessimenti parietali
Stereoma = insieme dei tessuti meccanici di un organo
- Piante giovani: turgore
cellulare
- Piante adulte: tessuti
specializzati per il sostegno
Tessuti meccanici
Sclerenchima
Collenchima
45
46
Da Nultsch.
Collenchima
• Cellule vive allungate in senso assiale
• Parete (primaria) con ispessimenti celluloso-pectici non
uniformi:
9 lungo le pareti tangenziali = collenchima lamellare
9 agli spigoli = collenchima angolare
9 in relazione a spazi intercellulari = collenchima lacunato
•
Le cellule collenchimatiche possono anche essere
fotosintetizzanti.
47
Collenchima angolare di
picciolo di Begonia
Collenchima lamellare di fusto
di Sambucus nigra
48
12
Collenchima in picciolo di Ninfea
Il collenchima è un tessuto che assicura elasticità
grazie agli ispessimenti cellulosici
Organi giovani,
ancora in via di
accrescimento
(es. fusti erbacei)
Collenchima in
fusto di girasole
Organi adulti per i
quali è necessaria
una buona
elasticità (es.
picciolo)
49
• I collenchimi sono presenti soprattutto nelle dicotiledoni.
• Nei fusti in genere occupano la porzione periferica,
come strato continuo o in forma di cordoni (es. fusto di
labiate: basilico, menta…).
50
Sclerenchima
• Parete cospicuamente e uniformemente ispessita
(parete secondaria)
• La deposizione degli strati parietali riduce
progressivamente il lume cellulare fino alla morte
del protoplasto: cellule morte
• Tipologie di cellule sclerenchimatiche:
Stachys alpina.
Rectangular stem. ©
2007 S. A. Mori.
51
9 Sclereidi: isodiametriche
9 Fibre: allungate
52
13
Sclereidi
Lo sclerenchima è un tessuto che assicura
resistenza grazie agli ispessimenti cospicui,
uniformi e spesso lignificati della parete cellulare
Cellule isodiametriche o di forma irregolare con parete lignificata.
• sclereidi organizzate in tessuto
- isole pietrose (polpa di pera, mela cotogna): deterrente per
erbivori
- endocarpo nei frutti (noce, pesca, ciliegia): protezione per il
seme
- tegumenti del seme (uva, cocomero…)
Organi che hanno
completato
l’accrescimento
A differenza del collenchima, le cellule sclerenchimatiche a
maturità svolgono esclusivamente una funzione meccanica:
il protoplasto viene riassorbito.
53
Esempi
Sclereidi filiformi
in foglia di olivo
• Sclereidi come idioblasti
- picciolo e lamina di Nymphaea (mantengono aperti i canali
aeriferi)
- foglia “cuoiose” (danno consistenza): tè, ulivo
54
Sclereidi in polpa di pera
Cellule pietrose
in pera
Sclereidi in
foglie di
Camellia
Sclereide a
estremità ramificate
in foglia di Hakea
Sclereide della corteccia
di Trochodendron
Da Gerola et al.
55
Colorazione con floroglucina HCl
56
14
Fibre
Sclereide come idioblasto
nell’aerenchima del
picciolo di ninfea
(Nymphaea alba)
Cellule sottili e allungate macroscopicamente e appuntite
alle estremità, spesso organizzate in fascetti/anelli.
Le fibre si distinguono sulla base della localizzazione:
• xilari: appartengono al legno, sempre lignificate
• extraxilari: corticali, liberiane… non sempre lignificate
Frequente crescita apicale: mentre aumenta in senso
centripeto lo spessore della parete, la cellula continua ad
accrescersi agli apici.
57
58
Fibre tessili
Lume del vaso
Le lunghe e resistenti cellule delle fibre possono essere
impiegate come fibre tessili.
Le fibre sono inserite nel contesto di un organo (fusto, foglia):
per il loro utilizzo devono essere separate.
• Fibre xilari
Difficilmente separabili dagli elementi di conduzione (fasci
conduttori); es. cordami ottenuti da foglie di monocotiledoni.
• Fibre extraxilari
Lume della fibra
Facilmente separabili per macerazione: processo di digestione
delle pectine (lamella mediana). Costituiscono le fibre tessili.
Da Lüttge et al.
59
60
15
Le fibre di canapa e lino differiscono per una varietà di aspetti
citologici usati a scopo diagnostico.
Aspetti citologici delle fibre tessili
La morbidezza della fibra decresce all’aumentare dal contenuto in
lignina nelle pareti cellulari:
Lino
Canapa
Terminazioni
affusolate
variabili, anche
biforcate
Superficie
striature trasversali
striature
longitudinali
Per uso tessile, la fibra deve avere un basso contenuto di lignina
(adatte alla tessitura): canapa e lino (fibre liberiane). Fibre con alti
contenuti di lignina possono essere solo intrecciate.
Dimensione del
lume
1/6-1/10 dello
spessore cellulare
1/2-1/3 dello
spessore cellulare
Punteggiature
minuscole,
difficilmente visibili
visibili e parallele
all’asse
longitudinale
Cotone (Gossypium sp.): non fibre sclerenchimatiche, ma sottili peli
che rivestono i semi (protezione, volo).
Sezione trasversale pentagonale o
esagonale
Linum usitatissimum (lino): sola cellulosa
Cannabis sativa (canapa): 50% lignina
Corchorus sp. (juta): 70% lignina
61
ovale o rotonda
62
Principali tessili di origine vegetale
in Europa
Canapa
Lino
• In Europa dal 3000 a.C., affianca il
lino.
• Meno pregiata per uso tessile.
• Campo pittorico: supporto dipinti,
rinforzo tele.
• Materiale “accessorio” (es.
riempimento ricami in rilievo)
• In Europa dal 4000 a.C., fibra più
importante fino al basso Medioevo.
• Vesti greche, tunica romana (bianca
per patrizi, scura per i plebei).
• Utilizzo per le vesti liturgiche in ambito
ecclesiastico.
63
64
16
Cotone
• In India nel 4000 a.C., nell’area
mediterranea da ca. 500 a.C.
• Uso frammisto al lino: i peli di cotone sono
relativamente corti e non offrono sufficiente
resistenza per la filatura. I cotoni più pregiati
erano importati dall’India.
• Industria cotoniera in Italia e Germania dal
XVII sec. Dall’800 il cotone diventa il filato
prevalente.
14. Tessuti adulti
• tessuti tegumentali
• tessuti segregatori
65
Tessuti tegumentali
H2O
Adattamento alla vita subaerea
Protezione contro gli agenti esterni
Regolazione dei flussi di sostanze
(liquidi, gas)
H2O
Elaborazione di tessuti specifici
per la protezione
67
66
Caratteristiche comuni:
9 assenza di spazi intercellulari tra le cellule del tessuto
9 modificazioni parietali caratteristiche (benché diverse per i vari
tessuti tegumentali)
9 creano una separazione tra due ambienti (anche non direttamente
con l’ambiente esterno)
• Origine primaria (dai meristemi apicali) – corpo primario
- esterni:
epidermide (germoglio)
rizoderma (radice)
esoderma (radice)
- interni:
endoderma (radice)
• Origine secondaria (dal fellogeno) – corpo secondario
- esterno: sughero
68
17
Funzioni generali:
Epidermide
Caratteristiche generali:
9 dal meristema apicale (protoderma), riveste il corpo
primario del germoglio
9 monostratificata (pluristratificata nelle piante di ambienti
aridi)
9 cellule vive, metabolicamente attive
9 parete cellulare: più spessa sul lato rivolto verso
l’atmosfera, cutinizzata
9 cuticola, rivestimenti cerosi
9 vacuolo: anche pigmentato (flavonoidi)
9 plastidi: indifferenziati, cloroplasti solo nelle piante
d’ombra
69
Cellule epidermiche “normali”
9 Protezione (agenti atmosferici, insetti, parassiti, eccesso di
irradiazione)
9 Regolazione della traspirazione e degli scambi gassosi
9 Altre funzioni metaboliche: secrezione, movimento, riserva,
percezione
Epidermide
Cellule epidermiche
“normali”
Formazioni
epidermiche
Apparati stomatici
Rivestimenti tricomatosi
70
La forma delle cellule epidermiche asseconda quella dell’organo che
rivestono. Esempio, foglie:
Cuticola con cere
Parete cellulare
Citoplasma
Nucleo
Casi particolari:
• pareti calcificate (foglie ruvide di cucurbitacee)
• pareti silicizzate (foglie taglienti di graminacee)
• pareti lignificate (foglie consistenti di conifere)
• pareti sottili nelle cellule bulliformi: grazie a variazioni di turgore fanno
arrotolare e srotolare la foglia delle graminacee
Monocotiledoni:
cellule allungate
71
Dicotiledoni: cellule con
contorno sinuoso
72
18
Rivestimenti tricomatosi - tricomi
I peli o tricomi derivano da cellule epidermiche che sono
andate incontro a divisione. A maturità possono essere vivi o
morti.
9 peli di protezione: morti a maturità, pieni d’aria, riflettono la
luce (contro l’eccesso di irradiazione solare e di
traspirazione)
9peli secretori: vivi, rilasciano metaboliti secondari (essenze)
o sali (alofite)
9 peli urticanti: vivi, rilasciano sostanze urticanti (istamina,
acetilcolina)
9 papille: peli conici vivi che conferiscono aspetto vellutato a
molti fiori
9 peli aggrappanti: organi di attacco, morti
9 squame assorbenti: assorbimento di acqua
73
Pelo lineare unicellulare
Gossypium sp.
Pelo lineare pluricellulare
Gynura aurantiaca
Pelo ramificato
Verbascum thapsus
Pelo peltato
Olea europaea
74
Pelo urticante
Urtica dioica
Pelo ghiandolare
capitato
Pelargonium sp.
La parete della cellula
urticante è mineralizzata:
- calcificazione (base)
- silicizzazione (apice)
Pelo contenente sale
Atriplex hymenelytra
(da Lüttge et al.)
Il pelo è portato da una base
pluricellulare (emergenza).
Da Lüttge et al.
75
Papille
Viola sp.
76
19
Rivestimenti tricomatosi emergenze
Peli aggrappanti - Phaseolus
Sono considerate rivestimenti
tricomatosi anche le emergenze.
Alla formazione delle emergenze
cooperano, oltre all’epidermide,
anche i tessuti sottoepidermici.
Squame assorbenti - Tillandsia
- aculei di rosa
- tentacoli di Drosera
Peter v. Sengbusch
www.biologie.uni-hamburg.de
77
78
Apparati stomatici
Posizione degli stomi:
Gli stomi sono presenti nelle parti aeree verdi (soprattutto
foglie). Rappresentano il sistema di regolazione della
traspirazione e degli scambi gassosi: apertura regolata.
1. sulla lamina fogliare
9
9
9
9
sulla pagina inferiore (caso comune): foglia ipostomatica
sulla pagina superiore (foglie natanti): foglia epistomatica
su entrambe le pagine (monocotiledoni): foglia anfistomatica
assenti (foglie sommerse): foglie astomatica
2. rispetto al piano dell’epidermide
Rima stomatica
9 allo stesso livello (caso comune)
9 sopraelevati (piante equatoriali)
9 infossati, anche in cripte stomatiche (piante di ambienti secchi)
Cellula di guardia
Cellula annessa
Camera sottostomatica
79
80
20
Distribuzione degli stomi:
dicotiledoni
Meccaniche stomatiche
monocotiledoni
1. Fenomeni osmotici a carico del vacuolo
2. Speciale deposizione delle fibrille di cellulosa
(micellazione radiale)
81
82
Da Raven et al.
Biogenesi dell’esoderma
Rizoderma ed esoderma
• Nelle cellule una lamella di suberina viene deposta contro la
parete. Successivamente si aggiungono strati di cellulosa e
lignina.
• Le cellule tendono a morire con la maturazione della radice.
• Alcune cellule non si modificano: punti di permeazione permettono
alla radice di non essere isolata dall’ambiente esterno.
• Epidermide della radice nella zona di attivo
assorbimento dell’acqua (zona pilifera)
• Cellule vive con parete sottile non cutinizzata
• Cellule con peli radicali, estroflessioni con funzione di
assorbimento
• Il rizoderma ha vita breve e la funzione tegumentale è
assunta dalle cellule dell’ultimo strato del cilindro
corticale = esoderma.
• Esoderma = tessuto primario che deriva dalla
modificazione delle cellule più esterne del cilindro
corticale.
Punti di permeazione
Rizoderma
Esoderma
Esoderma in
formazione
83
84
21
Endodermide:
Endodermide
• Tessuto interno primario
• Radice (ma anche fusti sotterranei e di piante acquatiche)
• Barriera al flusso apoplastico per la selezione delle
soluzioni che passano dal cilindro corticale al cilindro
centrale.
85
• monostratificata senza spazi intercellulari
• ultimo strato del cilindro corticale
• cellule vive
• presenza della banda del Caspary: banda della parete
primaria impregnata di suberina (zona idrofoba) – pareti
radiali e tangenziali
• il plasmalemma è strettamente associato alla banda del
Caspary
• il flusso di acqua che arriva all’endodermide attraverso
l’apoplasto si interrompe a livello della banda del Caspary,
dove avviene una selezione dei soluti
• i soluti selezionati entrano nel protoplasto e possono
riprendere la via apoplastica solo DOPO aver attraversato il
protoplasto dell’endodermide
86
Stadi di sviluppo dell’endodermide
Cilindro
corticale
1. Stadio primario: banda del Caspary
H2O
H2O Cilindro
centrale
Da Lüttge et al.
87
Da Mauseth
88
22
Via simplastica
2. Stadio secondario: deposizione di una lamella continua sottile di
suberina su ogni parete
Via apoplastica
Parenchima del
cilindro corticale
ENDODERMIDE
Parenchima del
cilindro centrale
Alcune cellule dell’endodermide non vanno incontro a
suberificazione: punti di permeazione o cellule di passaggio.
89
3. Stadio terziario: deposizione di strati successivi di cellulosa e
lignina sulle pareti radiali e tangenziali (funzione meccanica)
90
Da Lüttge et al.
1
2
3
Monocotiledoni (U)
Dicotiledoni (O)
Da Lüttge et al.
91
Da Raven et al.
92
23
Sughero
• Tessuto tegumentale del corpo secondario, ma anche
nei tuberi (patata).
• Origine dal fellogeno.
• Cellule morte, piene d’aria, appiattite trasversalmente
• Pluristratificato
• Parete cellulare: suberificazione, pigmentazione
(flobafeni)
• Barriera impermeabile a gas e acqua
• Proprietà coibenti e di difesa contro parassiti e agenti
chimici
Da Gerola
93
Lenticelle
Biogenesi delle lenticelle
• La barriera rappresentata dal sughero viene interrotta da
strutture dette lenticelle (nel fusto e nella radice).
• Scambi gassosi nella struttura secondaria (anche nelle
mele e nelle pere!)
Sughero
Fellogeno
della lenticella
• all’inizio della sua attività, in corrispondenza della futura
lenticella il fellogeno produce un tessuto parenchimatico
(anziché sughero) che giunge a lacerare l’epidermide
• le cellule esposte all’aria muoiono e si “sfarinano”, mentre
il fellogeno della lenticella continua a produrre nuove
cellule: lenticella aperta
Epidermide
lacerata
Botanical Society of America
Tessuto di
riempimento
94
95
• in autunno, il fellogeno della lenticella produce uno strato
di sughero: lenticella chiusa
• in primavera, il fellogeno forma di nuovo cellule
parenchimatiche che premendo contro il sughero lo
lacerano, riaprendo la lenticella
96
24
Tessuti segregatori
Segregazione nel vacuolo
Formati da cellule specializzate per l’elaborazione di sostanze
temporaneamente escluse dal metabolismo (spesso
metaboliti secondari).
Origine epidermica o parenchimatica.
Sulla base della localizzazione delle sostanze prodotte:
1.
2.
Cellule secretrici
Tubi laticiferi
Idioblasti con enorme vacuolo,
scarso citoplasma e pareti
impermeabilizzate con
suberina: a maturità spesso
muoiono mantenendo il loro
contenuto.
Latice = liquido sotto pressione
contenente una varietà di
sostanze del metabolismo
primario (glucidi semplici e
complessi) e secondario
(alcaloidi, tannini, resine,
politerpeni)
I tubi laticiferi sono polienergidi:
- di origine apociziale o
sinciziale
- ramificati o meno
- anostomosati o meno.
nel vacuolo = tessuti secretori
all’esterno delle cellule = tessuti ghiandolari
Spesso la funzione segregatrice è svolta da idioblasti.
Tessuti secretori
• Cellule a essenza
• Cellule a mirosina
(brassicacee)
• Cellule a mucillagini (liliacee)
97
98
Segregazione all’esterno della cellula
Tessuti ghiandolari
Cellule ghiandolari
Secrezione profonda, in spazi
intercellulari si origine
Cellule epidermiche specializzate
per la secrezione esogena:
1. schizogena: canali resiniferi
(conifere) e tasche
schizogene. Presenza di un
epitelio ghiandolare.
• peli ghiandolari (es. peli
geranio)
• peli urticanti (es. ortica)
• ghiandole del sale (alofite,
es.Atriplex)
• ghiandole nettarifere (fiori)
• peli digerenti (piante carnivore)
2. lisigena: tasche lisigene
(rutacee) generate dalla lisi
delle cellule che hanno
prodotto il secreto.
99
15. Tessuti adulti
• tessuti conduttori: vascolare e
cribroso
100
25
• Alghe: no necessità di trasporto dell’acqua; alcune alghe
brune (es. Laminaria, Macrocystis) elaborano vie di
conduzione degli assimilati con caratteri simili al tessuto
cribroso
Tessuti conduttori
Necessità di trasporto
delle soluzioni
• Briofite: semplici sistemi di conduzione solo nelle più
evolute (muschi)
9idroidi, con parete non lignificata, morti a maturità – flusso
ascendente
9leptoidi, cellule vive con protoplasto degenerato – flusso
discendente
Evoluzione di sistemi di trasporto
a lunga distanza specializzati:
tessuti conduttori
veri tessuti di conduzione solo nelle
Cormofite = TRACHEOFITE
• vascolare (linfa grezza)
• cribroso (linfa elaborata)
101
102
Caratteristiche fondamentali:
Tessuto vascolare
• cellule dalla forma allungata (elementi)
• disposizione longitudinale degli elementi (funzionale alla
conduzione)
• fortissima specializzazione
• origine primaria o secondaria
Movimento ascensionale (radici → foglie)
della linfa grezza = acqua & sali minerali
Tessuti
conduttori
Tessuto
VASCOLARE
Tracheidi
Tessuto
vascolare
Tessuto
CRIBROSO
Trachee
Cellule cribrose
e
cellule albuminoidi
Tubi cribrosi
e
cellule compagne
103
elementi
o
articoli
Cellule morte allungate che
conservano solo la parete
cellulare e che sovrapposte
formano i VASI
104
26
Trasporto ascensionale dell’acqua
Caratteristiche citologiche
• L’acqua si sposta spontaneamente dal suolo all’atmosfera
• Le piante si frappongono tra suolo e atmosfera, creando
un sistema entro cui l’acqua può fluire spontaneamente
• parete primaria sottile, non lignificata (permeabile)
• parete secondaria con ispessimenti lignificati disposti in
modo discontinuo (resistenza alla tensione)
• a maturità il protoplasto viene riassorbito (cellule morte)
VASI
• Per osmosi le cellule richiamano acqua
dalle cellule circostanti e infine dal sistema
di conduzione (= vasi)
Rigidità
• Nei vasi le molecole d’acqua formano una
colonna continua dalle radici fino alle foglie
Il trasporto dell’acqua in senso
ascendente avviene per la forza
105
aspirante della chioma
anulato
anulati
anulospiralati
spiralati
scalariformi
reticolati
punteggiati
spiralato scalariforme reticolato punteggiato
•Vasi rigidi, adatti agli organi
non in fase di crescita
•Alta resistenza
Tracheidi
pareti trasversali integre e punteggiate
piccolo diametro (0.03 mm)
elementi lunghi di forma affusolata
originano vasi chiusi: il trasporto nei vasi
è ostacolato dalla presenza delle pareti
trasversali
Piccolo diametro
Pareti trasversali
107
•Vasi estensibili, adatti ad
accompagnare la crescita
degli organi
•Limitata resistenza
106
•
•
•
•
Da Mauseth
Estensibilità
• L’acqua evapora dalle cellule delle foglie
FLUSSO LENTO
Da Raven
et al. Mod.
108
27
Le tracheidi costituiscono la forma più antica di tessuto
vascolare:
Trachee
• presenti in tutte le cormofite
• gruppi meno evoluti: spesso unico elemento di conduzione
• gimnosperme: fibrotracheidi, anche con funzione
meccanica
• angiosperme: primi elementi vascolari formati durante lo
sviluppo (anulati, spiralati, anulospiralati…) accompagnano
l’accrescimento
109
Le trachee si sono evolute dalle tracheidi
più volte in gruppi diversi di cormofite: casi
di convergenza morfologica
Da Mauseth
• pareti trasversali riassorbite parzialmente
o completamente (perforazioni semplici:
resta un cercine anulare tra gli articoli)
• ampio diametro (0.3 mm)
• articoli corti a forma di tamburo
• in genere, vasi reticolati e punteggiati
• originano vasi aperti: il trasporto non è
ostacolato
Ampio diametro
Perforazioni
Morfologia a
confronto
Perforazione
FLUSSO VELOCE
110
Da Raven et al. Mod.
Tracheide
Fibra
Fibrotracheide
• pteridofite evolute (es. Selaginella)
• gimnosperme Gnetafite (Gnetum,
Ephedra)
• tutte le angiosperme (eccetto le
Magnoliali)
111
Trachee
Da Raven et al. Mod.
112
Da Gerola
28
Flusso
dell’acqua a
confronto
Differenziamento delle trachee
Tracheidi
Trachee
113
Da Mauseth
Tessuto cribroso
• A livello delle foglie (sorgenti) i fotosintati prodotti vengono caricati
nei tubi cribrosi e diretti verso i siti di accumulo/utilizzo, es. radice
(pozzo).
• Altre vie: in primavera le sorgenti sono le radici che inviano
zuccheri alle gemme; durante la fioritura, i pozzi sono i fiori.
Tessuto
cribroso
Cellule vive
allungate
e sovrapposte
114
Trasporto dei fotosintati
Movimento della
linfa elaborata = acqua & fotosintati
Elementi
cribrosi
Da Raven et al.
• Ipotesi del flusso di pressione generato da
fenomeni osmotici
• La linfa elaborata è sottoposta a una
differenza di pressione generata tra sorgenti
e pozzi e che origina il flusso
Cellule
associate
115
1. La parete deve restare elastica, non
necessita di particolari “rinforzi”.
2. La cellula deve essere viva per
costituire un sistema osmotico. 116
29
Tubi cribrosi
Sorgente (foglia)
• Cellule (elementi) vive molto peculiari si
sovrappongono originando dei tubi
• Le pareti sono cellulosiche e sottili
• Pareti trasversali perforate = placche
cribrose semplici o composte
Placca
semplice
Placca
composta
Pozzo (radice)
Da Raven et al.
117
Le placche cribrose che
separano gli articoli
sono generalmente
poco oblique.
Aree cribrose
Da Mauseth
118
Caratteristiche degli elementi dei tubi
cribrosi
• assenza del nucleo
• pochi organuli, tra cui plastidi
• degenerazione del tonoplasto:
succo vacuolare e citoplasma non
sono distinti
• formazione dei pori cribrosi da
plasmodesmi “dilatati”
• proteine mucose formano sottili
filamenti che attraversano le
perforazioni delle placche cribrose
• plasmodesmi ramificati collegano i
tubi cribrosi con le cellule
compagne adiacenti
Piccole aree cribrose si
trovano sulle altre pareti
per il trasporto laterale.
Da Lüttge et al.
119
120
30
Caratteristiche delle cellule
compagne
Biogenesi degli elementi del tubo cribroso
• Origine comune col tubo che
affiancano (stessa cellula madre)
• La cellula compagna provvede a
“mantenere in vita” il tubo
cribroso, fornendogli l’energia
metabolica necessaria
(abbondanti mitocondri)
• Plasmodesmi multipli sono
l’espressione della dipendenza
del tubo cribroso dalla cellula
compagna
Da Raven et al.
121
Tubo
cribroso
Cellula
compagna
122
Cellule cribrose e cellule albuminoidi
Vita dei tubi cribrosi
• In genere, un anno: formazione del callo, un deposito di
callosio (β-glucano) che ottura le perforazioni delle aree
cribrose
• In alcuni casi, il callo può essere rimosso a primavera e
la funzionalità del tubo essere recuperata (es. vite, tiglio)
• Monocotiledoni arboree: anche 100 anni nelle Palme!
123
• Elementi del tessuto cribroso nelle tracheofite non
angiosperme
• Le cellule cribrose sono affusolate e comunicano tra loro
per mezzo di numerose aree cribrose con pori di ridotte
dimensioni
• Pareti trasversali molto oblique
• Aree cribrose si trovano su tutte le pareti
• Le cellule albuminoidi, che danno sostegno metabolico alle
cellule cribrose, non hanno origine comune con queste
ultime
124
31
I tessuti conduttori (vascolare e
cribroso) sono riuniti in
“cordoni” che percorrono il
cormo nel senso della
lunghezza:
continuità radice-fusto-foglie.
15. Xilema e floema
Gli elementi vascolari e quelli
cribrosi decorrono fianco a
fianco nello stesso cordone
• xilema e floema
• fasci cribro-fascolari
FASCI CRIBRO-VASCOLARI
125
126
Alla formazione del fascio contribuiscono:
porzione impegnata
nel trasporto della
linfa grezza
• elementi di conduzione: responsabili del trasporto delle
soluzioni in senso assiale
Fascio
cribrovascolare
porzione impegnata
nel trasporto della
linfa elaborata
• elementi meccanici (fibre): danno consistenza e
sostengono il fascio
• elementi parenchimatici:
9 funzione di riserva
9 trasporto a breve distanza (parenchima di trasporto)
9 trasporto a media distanza (in senso radiale)
127
XILEMA o LEGNO
FLOEMA o LIBRO
• trachee e tracheidi
• fibre xilematiche
• parenchima del legno
• tubi cribrosi
• cellule compagne
• fibre liberiane
• parenchima del libro
NON tessuti, bensì SISTEMI DI TESSUTI
128
32
Il differenziamento degli elementi che compongono i fasci
non è simultaneo nell’ambito del fascio stesso (distanza
dall’apice meristematico).
Dai
meristemi
primari
primo
differenziato
successivo
Dal
cambio
XILEMA
FLOEMA
protoxilema
protofloema
metaxilema
metafloema
deuteroxilema
deuterofloema
Le porzioni xilematica e floematica possono essere:
1. separate da uno strato di cellule ancora capaci di dividersi
e originare nuove cellule dello xilema e del floema:
cambio cribro-vascolare.
Fascio APERTO
Crescita secondaria possibile
2. essere a diretto contatto tra loro, senza strati cellulari
interposti.
Con l’attività del cambio, si modifica anche l’organizzazione degli elementi
(no fasci!).
129
Tipi di fasci
Fascio CHIUSO
Crescita secondaria impedita
130
Fasci radiali
I fasci vengono classificati sulla base della disposizione
reciproca di xilema e floema:
9 radiali
9 collaterali
9 bicollaterali
9 concentrici
La presenza di un tipo di fascio dipende dall’organo e dalla
posizione sistematica della specie.
131
• Partendo dal centro della struttura, xilema e floema
formano cordoni (arche) disposti su raggi alternati
• Le arche xilematiche e floematiche costituiscono una
struttura unitaria = fascio radiale (raggiato) o alterno
• Le arche xilematiche possono confluire verso il centro del
fascio
• Fascio presente nei fusti di alcune pteridofite (Psilotum) e
nelle radici.
132
33
Fasci radiali
Fasci collaterali
X
• Partendo dal centro della struttura, xilema e floema
formano cordoni disposti sullo stesso raggio
• Ogni unità è formata da un cordone xilematico e uno
floematico = fascio collaterale
• Presenza del cambio = fascio collaterale aperto
(gimnosperme e dicotiledoni)
• Assenza del cambio = fascio collaterale chiuso
(monocotiledoni)
F
F
X
133
134
Fasci bicollaterali
F
In talune famiglie di dicotiledoni (cucurbitacee, solanacee) i
fasci sono bicollaterali: lo xilema è compreso tra due
cordoni di floema.
Fasci anche aperti.
X
Cambio
F
F
X
X
F
135
136
34
Fasci concentrici
Fasci concentrici
perifloematico
• Perifloematici: il floema circonda lo xilema (fusto di
pteridofite)
• Perixilematici: lo xilema circonda il floema (rizoma di
monocotiledone)
• I fasci concentrici non presentano cambio (fasci chiusi)
X
F
perixilematico
F
X
138
Perixilematico
Perifloematico
Collaterale chiuso
Collaterale aperto
Bicollaterale
Radiale
137
139
35