Ecofisiologia Vegetale
Aa 2014-2015
Prof. Piero Picciarelli
Budda
From Mc Curry: Mali
From McCurry: Bamiyan, Afghanistan
Fotosintesi 2 parte
2. La luce come sorgente di energia (fotosintesi)
• Regolazione del ciclo C3, funzione della fotorespirazione;
metabolismi C4 e CAM e loro significato adattativo. I prodotti
della fotosintesi (amido, saccarosio e fruttani); le diverse
vie fotosintetiche: il rapporto isotopico del carbonio;
efficienza della conversione fotosintetica dell’energia solare
in energia chimica;
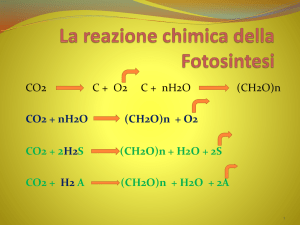
La reazione generale della fotosintesi
L’ATP e l’NADPH prodotti dalla fase luminosa sono
utilizzati nelle reazioni della fase oscura
Ribulosio 1,5bisfosfato
Ciclo C3 o ciclo di Calvin
Processo metabolico universale per la
fissazione e riduzione della CO2
Perchè ciclo C3?
Carbossilazione
Rigenerazione
Riduzione
il primo prodotto stabile
della reazione di
carbossilazione è uno
zucchero a 3 atomi
di C:
3-fosfoglicerato
o 3-PGA.
Ciclo C3 o ciclo di Calvin
Ribulosio 1,5bisfosfato
Il 3-PGA è ridotto utilizzando
l’ATP e il NADPH prodotti
nella fase luminosa.
Carbossilazione
Rigenerazione
Riduzione
Il processo di organicazione
della CO2 è un ciclo:
in cui viene
rigenerato l’accettore
iniziale della CO2 a 5C:
il ribulosio 1,5 difosfato
(Ru1,5 BP).
x
Ciclo C3 o ciclo di Calvin
x
3-fosfoglicerato
Ribulosio 1,5
bisfosfato
Processo ciclico costituito da
13 reazioni enzimatiche
(11 enzimi) localizzate nello
stroma del cloroplasto.
x
x
Carbossilazione
L’intero ciclo può essere
suddiviso in tre fasi distinte:
x
Rigenerazione
x
Riduzione
x
x
1) carbossilazione della
CO2 (1 enzima)
2) Riduzione del 3-fosfo
glicerato (3-PGA)
(2 enzimi)
x
x
x
x
x
3) rigenerazione del
substrato
(8 enzimi – 10 reazioni)
1) Carbossilazione della CO2
(3-PGA)
La reazione è catalizzata dall’enzima RUBISCO;
(25 % delle proteine totali della foglia, 50 % delle proteine solubili della foglia;
proteina più abbondante della biosfera).
Due caratteristiche dell’enzima favoriscono la reazione:
- G0 negativo grande (- 51 kJ mole-1)
- Alta affinità per la CO2 (Km = 8-20 M)
RUBISCO: carbossilasi e ossigenasi
(CA1,5P)
(3-PGA)
Per favorire la carbossilazione il sito catalitico della RUBISCO è
molto affine all’intermedio CA1,5P questo crea un:
complesso RUBISCO + CA1,5P molto stabile
riducendo il turnover catalitico (kccat ) dell’enzima
Costante catalitica di un enzima
RUBISCO costante catalitica carbossilante (c)
kccat = 1-10 s-1
RUBISCO
CO2
1-10
Con un turnover così basso servono molte molecole di RUBISCO
per sostenere un adeguato tasso di fotosintesi
Inoltre l’attività RUBISCO è inibita dalla produzione di prodotti secondari che si
formano durante l’attività catalitica, come: xylulose-1,5-bisphosphate (XuBP) e
il pentodiulose-1,5-bisphosphate (PDBP)
Enediolo composto intermedio
instabile; XuBP epimero del RuBP
(XuBP)
+ CO2
(RuBP)
+ O2
La formazione di questi due
composti aumenta con la
temperatura (35-45 °C)
(PDBP)
Struttura dell’enzima Rubisco
- 8 subunità proteiche grandi (Large)
Codificate nel cloroplasto
55 KDa x 8 = 448 KDa
(Colore blu chiaro e intenso)
- 8 subunità proteiche piccole (Small)
Codificate nel nucleo
14 KDa x 8 = 112 KDa
(Colore rosso e arancio)
Il sito catalitico si trova nelle subunità
grandi (1 sito per ciascuna) (Colore giallo)
Ciclo C3 o ciclo di Calvin
Ribulosio 1,5bisfosfato
Carbossilazione
Rigenerazione
Riduzione
2) Riduzione del 3-PGA
2 reazioni
fosforilazione
riduzione
(3-PGA)
Gliceraldeide
3-fosfato (G3P o GAP)
Nella fase di riduzione vengono consumati 2 ATP e 2 NADPH
prodotti nella fase luminosa ogni 2 di 3-PGA
Ciclo C3 o ciclo di Calvin
Ribulosio 1,5bisfosfato
Carbossilazione
Rigenerazione
Riduzione
3) Rigenerazione dell’accettore Ribulosio 1,5bifosfato
La rigenerazione dell’Ru-1,5BP è operata da una complicata
serie di reazioni enzimatiche (10) molto simili al ciclo dei
pentosi fosfati nel citosol
Soltanto l’ultimo passaggio
da Ru 5P + ATP
Ru 1,5BP + ADP
fosforibulosio chinasi
richiede energia sottoforma di ATP
3) Rigenerazione dell’accettore Ru 1,5bifosfato: reazioni enzimatiche
Ribulosio 1,5-bifosfato
Stechiometria del ciclo C3
Soltanto 1 su 6 molecole di triosi fosfati (TP) può essere
utilizzata per la sintesi di carboidrati
5 TP (15C)
Da
3 carbossilazioni
sono prodotti
1 TF (3C)
6 TF (18C)
Ricostituzione dell’accettore
Ribulosio 1,5 bifosfato
Sintesi
carboidrati
Consumo energetico del ciclo di Calvin
Ribulosio 1,5
bisfosfato
per fissare e ridurre
1 molecola di CO2
3 ATP
2 NADPH
Il ciclo di CALVIN è regolata da diversi meccanismi
A) Velocità di utilizzazione del carbonio fissato per la
sintesi dei carboidrati (amido e saccarosio);
B) Regolazione enzimatica operata dalla luce: il
sistema delle tioredoxine;
C) Attivazione catalitica della RUBISCO
B) Regolazione enzimatica: il sistema delle tioredoxine
Struttura secondaria
3--eliche e 4-foglietti-β
Tioredoxine
piccole proteine di 12 kDA ;
due residui di cisteina (cys 32 e 35)
legano lo solfo nel sito attivo;
La forma ossidata ha un ponte disolfuro
cys32-gly-pro-cys35
S
S
nella forma ridotta si forma il sulfidrile
cys32-gly-pro-cys35
SH
SH
B) Regolazione enzimatica: il sistema delle tioredoxine
Enzima INATTIVO
Enzima ATTIVO
Le tioredoxine sono ridotte dalla ferridossina, e a loro volta riducono i ponti
disolfuro di alcuni enzimi del Calvin, provocando una modifica
conformazionale che ne favorisce l’attività enzimatica.
C) Attivazione catalitica della RUBISCO
La RUBISCO è attivata dal legame con una molecola di
CO2 (non è la CO2 organicata) con la formazione della
forma carbamilata dell’enzima.
La carbamilazione è quindi stabilizzata da Mg2+
Rubisco attivasi: enzima che favorisce la carbamilazione
Attivazione RUBISCO da parte della Rubisco attivasi
In precedenza abbiamo visto che l’attività della RUBISCO è inibita da molecole che
si formano durante l’attività catalitica, come: XuBP e PDBP;
queste molecole si legano al sito attivo della RUBISCO, causano un
cambiamento conformazionale del sito inibendo l’enzima;
+
(XuBP e PDBP)
Sito attivo inibito
Sito attivo
la riattivazione della RUBISCO richiede l’intervento della Rubisco attivasi
La Rubisco attivasi è un enzima ATPasi che usa l’energia di idrolisi dell’ATP
per attuare un cambiamento conformazionale
e il rilascio degli zuccheri-P (Inibitori) attivando la RUBISCO
Il legame tra RUBISCO e gli inibitori (XuBP e PDBP)
avrebbe la funzione di proteggere l’enzima dalla proteolisi
in condizioni di stress abiotici transienti;
La RUBISCO può essere riattivata rapidamente al ristabilirsi di
condizioni ambientali ottimali senza la necessità di sintetizzare
de novo l’enzima;
(XuBP e PDBP)
RUBISCO, Rubisco attivasi e temperatura
Camelina sativa
1800 µmoli m-2 s-1
CO2 380 ppm
O2 21 %
La RUBISCO è completamente attivata ad alta irradianza
e con temperature sotto 30 °C;
Sopra 30 °C l’attivazione della RUBISCO cala molto con
diminuzione dell’assimilazione della CO2;
Tutto questo dipende dalla Rubisco attivasi che è inattivata
alle temperature elevate
Fam. Brassicaceae
Falso lino
Ottimo di temperatura della Rubisco attivasi
Camelina: cool-crop specie
Cotton: warm-crop specie
L’ottimo di temperatura della Rubisco attivasi è diverso tra specie
adattate ad ambienti freddi (~ 25 °C) e caldi (~ 30 °C)
Ottimo di temperatura della Rubisco attivasi
Camelina: cool-crop specie
Cotton: warm-crop specie
L’introduzione della Rubisco attivasi di cotone in Camelina potrebbe
migliorare la termotolleranza di questa pianta di circa 5 °C
Questa stesso approccio potrebbe essere perseguito anche con altre
specie coltivate (es. Grano) per migliorare la termotolleranza
RUBISCO: ribulosio 1,5 bifosfato carbossilasi-ossigenasi
Le reazioni di carbossilazione e ossigenazione
avvengono nello stesso sito catalitico dell'enzima:
CO2 e O2 sono substrati competitivi
Quale delle due reazioni prevale ?
Fornendo uguali concentrazioni di CO2 e O2
prevale la reazione di carbossilazione con rapporto:
3
Carbossilazioni
a
1
Ossigenazione
Se prevale la carbossilazione significa che la Km della
RUBISCO per la CO2 è più bassa della Km per l' O2:
Km CO2 = 8 - 20 M
Atmosfera 0.04 % = concentraz. in soluz. 8-22 M
Km O2 = 180 - 400 M
Atmosfera 21 % = concentraz. in soluz. 230-400 M
L’affinità della RUBISCO per la CO2 e la O2 è diversa tra le specie;
studiata solo in 100 specie !!
Fotorespirazione
Fotorespirazione: consumo di O2 ed evoluzione di CO2 alla luce
Il 2-fosfoglicolato da inizio alla fotorespirazione (Ciclo C2)
I cicli C2 e C3 operano in modo integrato
- Il ciclo C2 dipende dal ciclo C3 per
l’accettore di O2, il Ru 1,5 BP
- Il ciclo C2 è attivo soltanto alla
luce perché, richiedendo energia,
dipende dal trasporto fot. degli e-
- La CO2 prodotta nel ciclo C2
è un substrato nel ciclo C3
- L’O2 prodotto nella fase luminosa del
ciclo C3 è un substrato nel ciclo C2
Nel ciclo C2
2 x di fosfoglicolato (4C),
(RUBISCO + O2 x 2)
sono convertite in:
1 di CO2 (1C)
1 di 3-PGA (3C)
(rientra nel ciclo C3)
Il ciclo si svolge in
tre organelli:
cloroplasto
perossisoma
mitocondrio
Nei cloroplasti
il fosfoglicolato è
defosforilato
a glicolato
ed esportato
ai perossisomi
Perossisoma
Il glicolato è convertito
in gliossilato
(glicolato ossidasi,GO)
GO produce H2O2
eliminata dalla catalasi
Per transaminazione
(aminotransferasi) il
gliossilato (x 2) è
convertito in glicina (x 2)
La glicina è esportata
nei mitocondri
Mitocondrio
Due enzimi la
glicina decarbossilasi e la
serina metiltransferasi
catalizzano le reazioni:
due glicine (4C)
sono convertite in:
- 1 serina (3C)
- 1 CO2
- 1 NH3 (al cloroplasto
per la riassimilazione)
- 1 NADH (mitocondrio o
perossisomi ?)
La serina è esportata
nel perossisoma
Perossisoma
Dalla serina per
transaminazione si forma
l’idrossipiruvato
+ NADH
NAD+idrossipiruvato
reduttasi
forma il glicerato
Il glicerato è esportato
nel cloroplasto
+ ATP
Glicerato chinasi
forma il 3-PGA
(rientra nel ciclo C3)
Costo energetico del ciclo C2
2 ATP (glicerato chinasi e riassimilazione NH3)
e 2 Fdx ridotte (o 1 NADPH) (riassimilazione NH3)
per ogni 2 molecole di fosfoglicolato prodotte
Considerando anche il costo energetico per fissare e ridurre la
CO2 liberata e la rigenerazione del Ribulosio 1,5 BP il costo
complessivo arriva a (3+2) 5 ATP e (2+1) 3 NADPH
Funzione della fotorespirazione
Il ciclo C2 permette il rientro parziale del C del
fosfoglicolato nel ciclo di Calvin
Il ciclo C2 perciò recupera 3/4 ( 75 %) del C che sarebbe
perduto a causa della reazione
di ossigenazione della RUBISCO
Quali sono i fattori che influiscono sulle due reazioni
catalizzate della RUBISCO ?
1. Proprietà cinetiche della RUBISCO
2. Concentrazione di CO2 e O2 nel sito catalitico
1. Proprietà cinetiche della RUBISCO
Il rapporto tra le due attività (carbossilasi / ossigenasi) è in
funzione dei parametri cinetici delle due reazioni competitive:
τ = Vc Ko / Vo Kc
Vc e Vo = Vmax carbossilazione
e ossigenazione
Kc e Ko = Km per CO2 e O2
Questi parametri cinetici rappresentano il fattore di specificità
(τ): il rapporto tra la velocità di carbossilazione/
ossigenazione quando i gas CO2 e O2 sono presenti
alla stessa concentrazione
τ degli organismi
fotosintetici
Cianobatteri
40-50
Alghe Rosse
100-160
Piante vascolari
70-120
Nelle piante, a parità di concentrazione dei gas, la RUBISCO
reagisce 70-120 volte più velocemente con la CO2 che con l’O2
Specificity factor (SF; τ) fattore di specificità
Alto: favorita la carbossilazione
Basso: favorita l’ossigenazione
Con le attuali condizioni dei gas nell’atmosfera
CO2 = 0.04 % = in soluzione 8 µM;
O2 = 21% = in soluzione 250 µM
e con τ = 100
vc
CO2
---- = ------ × τ
vo
O2
Quindi il rapporto tra vc\vo = 8 /250 x 100 = 3.2
Relazione tra specificity factor (SF) e kccat
Relazione inversa tra SF e kccat : in piante C4 la RUBISCO lavora
in un ambiente ricco di CO2 ed ha valori di kccat più alti mentre i valori
di SF sono più bassi viceversa nelle piante C3.
Tasso di fotosintesi in grano e Limonium (Ac) in funzione
della CO2 interna (mesofillo) e ad irradianza saturante
Rubisco di Limonium:
- più affine alla CO2
- meno affine all’O2
La RUBISCO di Limonium ha proprietà catalitiche migliori di quelle
del grano e questo si riflette con un aumento del tasso di fotosintesi;
Inserire la RUBISCO di Limonium in grano potrebbe incrementare
in modo significativo il tasso di fotosintesi e quindi la produzione in grano
2. Concentrazione di CO2 e O2 nel sito catalitico
CO2
O2
CO2/O2
Atmosfera
0.04 %
21 %
0.0019
Soluzione
acquosa
8 M
250 M
0,032
In acqua il rapporto CO2 / O2 è più alto che nell’atmosfera
questo favorisce la carbossilazione
Quanta CO2 viene effettivamente liberata nell’ambiente
naturale per effetto della fotorespirazione?
La temperatura è il principale fattore
ambientale che influenza il rapporto
tra le due reazioni della RUBISCO
La temperatura influisce:
Solubilità in
acqua di CO2 e O2
Fattore di
specificità τ
Diminuisce
all’aumentare della temperatura
All’aumentare della temperatura la solubilità di CO2 e O2 in
acqua diminuisce, ma per la CO2 diminuisce di più rispetto all’O2
CO2 (M)
O2 (M)
CO2/O2
5 °C
22
400
0,055
15 °C
16
320
0,05
25 °C
12
265
0,045
35 °C
9
230
0,039
L’aumento della temperatura favorisce la reazione di
ossigenazione rispetto alla carbossilazione.
PIANTE C 4
1% delle piante superiori
Mais, sorgo, canna da zucchero, miglio, numerose piante
tropicali e 8 su 10 piante infestanti (gramigna, amaranto)
Perchè C4?
Primo prodotto stabile è un
composto a 4 atomo di carbonio
Caratteristiche distintive del metabolismo C4
Anatomia Kranz
Due carbossilazioni
successive
Piante C4: anatomia Kranz
Kranz (corona)
- Cellule della guaina del fascio (BS)
- Cellule del mesofillo (M)
Schema generale del
metabolismo C4
Le due carbossilazioni
1a carbossilazione
PEP carbossilasi (PEPasi)
cellule del mesofillo (M)
2a carbossilazione
RUBISCO
cellule della guaina
del fascio vascolare (BS)
Tra i due tipi di cellule vi è uno
scambio di metaboliti favorito dalla
presenza di numerosi plasmodesmi
Dettagli reazioni del metabolismo C4
Nelle foglie C4 la PEPcarbossilasi costituisce ~ il 15 % delle
proteine solubili (circa 20 volte più alta che
nelle foglie C3). Localizzata nel citosol
PEP + HCO3-
OAA + Pi
PEPcarbossilasi
G° = - 33 KJ mole-1
PEPasi Km (HCO3-) = 30 M (= 6,4 M CO2 a pH 7)
In acqua
pKa 3.45 (non 6.35)
HCO3- + H3O+
CO2 + 2H2O
In H2O si trovano essenzialmente CO2 e HCO3H2CO3 è 1/400 della CO2 disciolta
Nelle cellule del mesofillo è presente l’anidrasi carbonica
(AC) che converte rapidamente una forma nell’altra e viceversa
AC
CO2
HCO3L’anidrasi carbonica è assente nelle BS
La decarbossilazione dell’acido organico nelle cellule
BS porta ad un accumulo di CO2 che raggiunge una
concentrazione fino a 6 volte quella dell’atmosfera
2100 ppm ( 0,21% )
Le cellule BS hanno una elevata resistenza alla
diffusione della CO2
parete cellulare suberificata
la reazione ossigenasica
della RUBISCO è
sostanzialmente impedita
Le piante C4 non hanno fotorespirazione
Metabolismo C4
Ricostituzione del PEP nei cloroplasti cellule mesofillo
dal piruvato ha un costo energetico elevato (2 ATP);
reazione catalizzata dall’enzima piruvato fosfato dichinasi ( PPDK)
caratteristico delle piante C4 e assente nelle piante C3.
Alcuni enzimi del ciclo C4 sono regolati dalla luce
Questa regolazione è critica per coordinare le attività
metaboliche tra le cellule del M e della BS
- NADP malico deidrogenasi
attivato dalle tioredoxine
La Luce attiva la
PEPcarbossilasi
tramite fosforilazione
La luce attiva
la PPDK tramite
defosforilazione
Nelle piante C4 si verifica una effettiva divisione del ciclo di
Calvin nei due tipi di cellule fotosintetiche
Cloroplasti cellule M
producono ATP e NADPH
non hanno RUBISCO né
diversi enzimi del ciclo C3
Cloroplasti cellule BS
carenti di grana e del PSII
non formano NADPH
Cellule BS
Carenti di grana e del PSII non
formano NADPH. Producono ATP
con il trasporto ciclico.
3-PGA
Cellule M
producono ATP e NADPH non hanno
RUBISCO né diversi enzimi del ciclo C3
3-PGA
Rubisco
Ru 1,5 BP + CO2
Fosfoglicerato
chinasi;
Gliceraldeide 3P
deidrogenasi
ATP
NADPH
Triosi-P
Triosi-P
Amido primario
Sintesi di saccarosio
(citosol)
Funzione: nelle cellule BS l’assenza del PSII (assenza di fotolisi dell’H2O),
favorisce ulteriormente la reazione di carbossilazione
della RUBISCO
3 varianti del metabolismo C4
Sulla base
enzima che opera la
decarbossilazione nelle cellule BS
Enzima malico NADP+dipendente
tipo di acido C4 trasportato
malato
Enzima malico NAD+dipendente
aspartato
PEPcarbossichinasi
aspartato
Variante enzima malico NADP+ dipendente
Piante: sorgo, giavone, gramigna, mais, canna da zucchero
Variante enzima malico NAD+ dipendente
Cellule del mesofillo
Cellule della guaina del fascio
Piante: Amaranto, Portulaca olearacea, miglio
Variante PEPcarbossichinasi
Cellule del mesofillo
Cellule della guaina del fascio
Piante: panico verga (Panicum virgatum)
(Produzione di biocarburante)
Bilancio energetico del metabolismo C4
Richiesta energetica per organicare 1 molecola di CO2:
2 NADPH
+
5 ATP
(3 + 2 da PPDK)
Le piante C4 sono energeticamente meno efficienti
delle piante C3 ?
No, perchè le C3 perdono una quota rilevante
di carbonio con la fotorespirazione
La fotorespirazione aumenta con la temperatura
• A temperature < 25-30 °C le C3 sono avvantaggiate
perchè
la perdita di CO2 per fotorespirazione è
contenuta.
• A temperature > 25-30 °C le C4 sono avvantaggiate perché
la perdita di CO2 e l’aumento del costo energetico
causati dalla fotorespirazione sono superiori al maggior
costo energetico del ciclo C4.
Le piante C4 si sono originate nelle
regioni calde della terra
Il tasso di fotosintesi netto delle piante C4 è più elevato
di quello delle piante C3
C4 = 25-30 moli CO2 m-2 s-1
C3 = 15-25 moli CO2 m-2 s-1
Vantaggi accessori del metabolismo C4 rispetto
al metabolismo C3
1. Migliore uso dell’N
2. Migliore uso dell’H2O
1. Migliore uso dell’N
Nelle piante C4 in assenza della reazione ossigenasica la
RUBISCO è più efficiente nella reazione di carbossilazione.
La stessa quantità di CO2 può essere fissata da
una quantità minore di enzima con risparmio di N
La RUBISCO costituisce
nelle piante C3 ~ il 25 % (25 % N totale della foglia)
nelle piante C4 ~ il 9 %
delle proteine totali della foglia
2. Migliore uso dell’H2O
Le piante C4 hanno una migliore efficienza nell’uso
fotosintetico dell’H2O (WUE) in confronto alle C3.
C3
C4
1 - 3 g CO2 fissata Kg-1 H2O traspirata
2-5g «
«
«
«
Questo per le piante C4 è
correlato ad una minore
conduttanza stomatica
Come conseguenza anche la concentrazione della CO2 nel
mesofillo fogliare è minore rispetto alle piante C3:
C3
C4
250 ppm
150 ppm
Tuttavia questo non limita l’attività della PEPasi
poichè non ha attività ossigenasica
Metabolismo C4 in cellule singole
Metabolismo C4 in piante senza anatomia Kranz
(Nature, 2002)
Borszczowia aralocaspica
Bienertia cycloptera
Chenopodiacee
semi-succulente
dei deserti salini
dell’Asia centrale
In queste due piante il metabolismo C4 si realizza
all’interno delle singole cellule
Metabolismo C4 in cellule singole di Borszczowia aralocaspica
Distribuzione non uniforme degli enzimi e dei
cloroplasti fra zone diverse della stessa cellula:
- Pochi cloroplasti (grana ridotti) superficie foglia,
privi di amido e con PPDK;
- PEPCasi (citosol)
- Zona centrale citoplasmatica priva di organelli
- Numerosi cloroplasti (grana sviluppati) con RUBISCO,
accumulano amido;
- Decarbossilazione malato (mitocondri):
ME NAD-dipendente
Metabolismo C4 in cellule singole di Bienertia cycloptera
Cellule del mesofillo
Mostrano un comparto citoplasmatico
centrale circondato dal vacuolo
e un comparto citoplasmatico periferico
Comparto citoplasmatico centrale
- Decarbossilazione malato
- Rubisco nei cloroplasti centrali
Canali citoplasmatici (CC) connettono
i due comparti per lo scambio di metaboliti
Comparto citoplasmatico periferico
- PEPCasi (citosol)
- PPDK (cloroplasti)
Strategie biotecnologiche per aumentare l’assimilazione
del carbonio nelle piante coltivate
a)
b)
c)
d)
e)
Migliorare le proprietà catalitiche della RUBISCO
Introduzione del metabolismo C4 in piante C3
Aumentare il livello di CO2 in prossimità della RUBISCO
Riciclaggio del 2-fosfoglicolato nei cloroplasti
Favorire la rigenerazione del RuBP aumentando l’attività
dell’enzima sedoeptulosio 1,7BP
b) Introduzione del metabolismo C4 in piante C3: riso (Science 2012)
Studi recenti hanno dimostrato che non è sufficiente inserire in riso i geni chiave
del metabolismo C4 occorre modificare anche l’anatomia fogliare.
Anatomia foglie di Riso (C3)
Cellule guaina del fascio poco sviluppate
e con un basso numero di cloroplasti.
Elevato numero di cellule nel mesofillo tra
i fasci vascolari (tipico delle specie C3)
C4
Anatomia foglie di Sorgo (C4)
Cellule guaina del fascio sviluppate e con
diversi cloroplasti.
Solo 2-3 strati di cellule nel mesofillo tra i
fasci vascolari (tipico delle piante C4)
c) Aumentare il livello di CO2 in prossimità della RUBISCO
I cianobatteri hanno delle pompe
(CO2 e HCO3-) per concentrare la
CO2 (di 1000 volte)
in prossimità della RUBISCO
L’inserimento di queste pompe
nella membrana del cloroplasto
di piante C3 potrebbe aumentare
il tasso di assimilazione della
CO2 di circa 5-15 %
d) Riciclaggio del 2-fosfoglicolato nei cloroplasti
La via catabolica del glicolato
di E. coli (in rosso) inserita
nei cloroplasti di Arabidopsis;
(in nero) la fotorespirazione;
Il glicolato è trasformato in
glicerato nei cloroplasti e
questo riduce il flusso
fotorespiratorio ma non lo
elimina
Le piante transgeniche crescono più
velocemente, producono più biomassa
(germogli + radici) e con più zuccheri solubili.
e) Favorire la rigenerazione del RuBP aumentando l’attività
dell’enzima sedoeptulosio 1,7BP (SBPasi)
Piante di tabacco che overesprimono SBPasi mostrano un aumento
significativo di biomassa ad un livello di CO2 di 550 ppm
PIANTE CAM (Crassulacean Acid Metabolism)
10 % delle piante superiori hanno il metabolismo CAM
Adattamento del metabolismo fotosintetico in
piante che vivono in ambienti aridi
- piante grasse come cactaceae, crassulaceae ed alcune euphorbiaceae
- piante tropicali epifite come orchidaceae e bromeliaceae.
Alcune CAM sono importanti dal punto di vista economico:
ananas ( bromeliaceae ), vaniglia ( orchidaceae )
e agave (produzione di tequila).
Caratteristiche anatomiche delle piante CAM
- Tessuti succulenti (foglie e steli)
Succulenza = basso rapporto superficie/volume
Pagina superiore,
(adassiale);
Pagina inferiore,
(abassiale);
Crassula ovata
Foglie spesse con superficie liscia
per la presenza di cere
Caratteristiche anatomiche delle piante CAM
- A livello cellulare la succulenza è caratterizzata da cellule
grandi con un grosso vacuolo (permette grande capacità di
stoccaggio di acidi organici) e una parete cellulare spessa.
1- Cellula con tannini;
2- Parete cellulare;
3- Spazio intercellulare;
4- Vacuolo cellule
clorenchimatche;
5- Cloroplasti
Caratteristiche anatomiche delle piante CAM
- Basso numero di stomi e cuticola più spessa:
forte limitazione degli scambi gassosi con l’esterno.
Strip di foglia
1- Stomi
2- Epidermide
3- Pezzi di cuticola
La cuticola è formata
da cere e cutina
Caratteristiche del metabolismo CAM
stomi chiusi di giorno e aperti di notte
1 notturna
2 carbossilazioni
1 diurna
Il ciclo di carbossilazione e decarbossilazione
ricorda quello delle piante C4 ma è separato nel
tempo anziché nello spazio
Metabolismo CAM: notte
Carbossilazione notturna
operata dalla
PEPcarbossilasi
Il PEP è rigenerato dall’amido
Metabolismo CAM: giorno
Decarbossilazione del malato
ad opera della
NADP+ enzima malico
e rifissazione della CO2
Sintesi di amido e saccarosio
La concentrazione di acido malico nel vacuolo alla fine
della notte raggiunge valori molto elevati
( 200 mM con pH 3)
Decarbossilazione diurna
dell’acido malico
[CO2] = 2-4 % (20-40mila ppm)
L’elevata concentrazione di CO2 inibisce
la fotorespirazione
Le CAM minimizzano la perdita di H2O,
WUE è molto alta: 10-40 g CO2 fissata Kg-1 H2O traspirata
Canna da zucchero
Ananas
mesofillo
CO2 incorporata in
acidi organici a 4 C
guaina del
fascio
Separazione spaziale
notte
giorno
Decarbossilazione
acidi organici, CO2
fissata e ridotta nel
ciclo di Calvin
Separazione temporale
Regolazione del metabolismo CAM
flusso di malato in e
out dal vacuolo
Sensibilità della PEP
carbossilasi al malato
Il flusso di malato:
• in è passivo (canali);
• out è attivo (proteine carrier)
3.0 mM
(notte)
Meno sensibile
al malato
0.3 mM
(giorno)
Più sensibile
al malato
Il cambiamento di sensibilità della PEPasi
è regolato tramite fosforilazione/defosforilazione
Piante CAM inducibili (Cycling)
Molte piante CAM in condizioni
ambientali favorevoli (disponibilità di
H2O) operano la fotosintesi tramite il
metabolismo C3.
Es: specie floricola il
Mesembryanthemum spp
Ciclo CAM futile (minimo = idling)
Con siccità molto forte le piante CAM
tengono gli stomi chiusi anche
di notte riciclando la CO2 respiratoria.
Le piante non presentano accrescimento
ma sopravvivono molto a lungo
Guzmania monostachia (L.)
I principali prodotti di accumulo della fotosintesi
Amido
Sintesi nei
cloroplasti
Triosi fosfati
Saccarosio
Sintesi nel
citoplasma
Fruttani
Sintesi nel vacuolo
In parallelo con la sintesi di saccarosio c’è sintesi di piccole quantità di trealosio
TPS: trealosio sintasi
TPP: trealosio fosfatasi
Tl: trealasi
Il trealosio è presente nelle piante a concentrazione µM, questo suggerisce
un ruolo di regolazione piuttosto che un ruolo metabolico
Amido
Polimero formato da molecole di glucosio con legame 1
Due componenti: l’amilosio e l’amilopectina
Amilosio: polimero lineare (circa 1000 unità di glucosio)
L’amilopectina ha struttura ramificata (legami 1 6)
(circa 105-106 unità di glucosio)
Composizione approssimativa amido
amilosio ~ 10-30 %
amilopectina ~ 70-90 %
assemblati in granuli con piccole quantità di lipidi, proteine e ioni
Endosperma
grano
Endosperma
mais
Endosperma
riso
Scala bar
10 μm
Tubero patata
Ø granuli
Patata 100 m
Radici cassava
Foglie
Arabidopsis
Ø granuli
Arabidopsis
1-2 m
La struttura del granulo di amido
Struttura ad albero:
catene vicine (es. A e B)
formano una doppia elica
Una singola elica
Le doppie eliche di amilopectina sono assemblate in ordinate lamelle cristalline
di due tipi A-type (tipico dei cereali) e B-type (tipico dei tuberi e amido fogliare).
Le lamelle cristalline (6 nm) sono alternate con lamelle amorfe
(3 nm: contengono le ramificazioni delle catene di amilopectina).
Le singole eliche di amilosio sono orientate in modo casuale (random)
all’interno delle lamelle amorfe
Lamelle cristalline + amorfe (= 9 nm) ripetute formano sovrastrutture
dette blocchetti (Ø tra 20-500 nm)
I blocchetti possono essere di due tipi: normali e difettosi
Blocchetti normali
Blocchetti difettosi:
assemblamento
amilopectina difettoso
per la presenza di
amilosio e altre molecole
L’assemblaggio dei blocchetti porta alla formazione degli anelli tipici dei granuli:
anelli cristallini (blocchetti normali)
e anelli semi-cristallini (o amorfi) (blocchetti difettosi)
L’amido è insolubile in H2O fredda: gli anelli cristallini
impediscono l’ingresso di H2O nel granulo.
Amilosio e preparazioni alimentari
Il contenuto di amilosio ha un effetto molto forte sulle
proprietà funzionali dell’amido utilizzato nelle
preparazioni alimentari (es. preparazione della pasta)
Amido primario e amido secondario
Leaf cell
Sintesi amido primario:
stroma del cloroplasto
(di giorno)
Sintesi amido secondario:
in organi di riserva dall’amido
primario (di notte )
Amido secondario
Saccarosio
E’ un disaccaride: glucosio + fruttosio (legame 1 2)
Adatto al trasporto floematico perché non-riducente
Maltosio: glucosio + glucosio (legame 1 4)
zucchero riducente, gruppo aldeidico libero,
in grado di ridurre ioni inorganici (es. Ag+1 e Cu2+)
La sintesi di amido nel cloroplasto e di saccarosio
nel citosol sono processi in competizione
Il citosol e il cloroplasto sono collegati attraverso il
traslocatore Pi / triosi-P (piccola proteina 30 kDa
della membrana interna del cloroplasto)
trasporto di triosi-P e Pi (1:1) in direzione
opposta tra il cloroplasto e il citosol
La sintesi di Amido e
Saccarosio è controllata
principalmente
concentrazioni relative di
triosi-P e Pi nei due
comparti cellulari
Sintesi amido e saccarosio
Bassa concentrazione
di [Pi] nel citosol
limita l’esportazione dal
cloroplasto di triosi-P
Favorita la
sintesi di amido
Sintesi amido e saccarosio
Alta concentrazione
di [Pi] nel citosol
stimola l’esportazione dal
cloroplasto di triosi-P
Favorita la
sintesi di saccarosio
Sintesi di amido nel cloroplasto
Amido
sintasi
ADP-glucosio pirofosforilasi: controlla la sintesi di amido
- stimolata da alta concentrazione di 3-PGA (basso esporto di triosi-P)
- inibita da alta concentrazione di Pi
(alto esporto di triosi-P)
Sintesi di amilosio ed amilopectina
Sintesi amilosio: amido sintasi.
Sintesi amilopectina: enzimi ramificanti I e II idrolizzano i legami
1 dell’amilosio e con un legame 1 6 attaccano una
catena laterale; di norma una catena ogni 20 molecole di glucosio.
Sintesi di saccarosio nel citosol
Regolata principalmente dall’attività della
Saccarosio fosfato sintasi (SPS)
Saccarosio fosfato sintasi (SPS)
- fosforilazione / defosforilazione
(- attivo)
(+ attivo)
- effettori allosterici positivi (glucosio 6-P) e negativi (Pi)
Sintesi di saccarosio nel citosol
Altro punto di regolazione
della sintesi di saccarosio
Nel citosol questo passaggio
è catalizzato da tre enzimi.
PFK-ATP
PFP-PPi
FBPasi
L’attività di questi enzimi è regolata da metaboliti tra
cui il fruttosio 2,6 BiP
Fruttani
Sono polimeri del fruttosio:
Fruttani
ad una unità di glucosio si legano
le molecole di fruttosio con
legami , 2-1 (inulina) o , 2-6
(levani)
La sintesi è localizzata nel vacuolo
a partire dal saccarosio
Si accumulano nei tuberi, radici e
bulbi di:cipolla, asparago, tulipano,
dalia, ecc.
Di particolare interesse è la
presenza di fruttani negli steli e
foglie delle graminaceae.
Nei cereali i fruttani svolgono una funzione importante
nell’acclimatazione alle basse temperature
Durante una prolungata esposizione alle basse temperature
(3-10°C) non c’è traslocazione di saccarosio, perché la
crescita è rallentata.
Si accumulano i fruttani (aumenti del 20-30 %)
Quando la temperatura risale riprende la crescita delle piante
accompagnata da una forte diminuzione del pool di fruttani.
I fruttani hanno effetti benefici
sulla salute umana
La principale fonte di fruttani per l’industria sono i tuberi del Topinambur
(Helianthus tuberosus) e le radici di cicoria (Chicorium intybus)
Trasferimento di zuccheri dalle cellule vegetali
alle cellule di funghi simbionti (es. micorrize)
La pianta cede al fungo fino al 30 % degli zuccheri prodotti
mentre il fungo fornisce alla pianta P (fino all’80 %) e azoto
Il fungo assorbe esosi tramite un trasportatore (Hex carrier)
presente negli arbuscoli e ife del fungo
Nelle cellule vegetali il saccarosio viene scisso in glucosio e fruttosio tramite
l’invertasi del vacuolo (VIN) o del citosol (CIN) o dell’apoplasto (CWIN);
Le micorrize inducono nelle cellule vegetali l’espressione di queste
invertasi in particolare, CWIN e CIN, tramite effettori proteici
prodotti dalle cellule del fungo ancora sconosciuti
Trasferimento di zuccheri dalle cellule vegetali
alle cellule di patogeni biotrofici (funghi, batteri e virus)
Bacterial blight is caused
by Xanthomonas oryzae
Pseudomonas syringae
I patogeni aggrediscono le cellule vegetali per ottenere zuccheri
senza alcun beneficio per l’ospite
Batteri fitopatogeni (es. Pseudomonas e Xanthomonas) vivono nell’apoplasto
e acquisiscono zuccheri dalle cellule vegetali infettate;
Tramite effettori proteici i batteri inducono nelle cellule vegetali
l’espressione del trasportatore di saccarosio SWEET e di CWIN;
SWEET aumenta l’efflusso di saccarosio nell’apoplasto;
Il saccarosio nell’apoplasto viene idrolizzato da CWIN e gli esosi vengono
trasportati alle cellule batteriche tramite Hex carrier
Tramite effettori proteici i batteri inducono nelle cellule vegetali
l’espressione del trasportatore di saccarosio SWEET aumentando
l’efflusso di saccarosio nell’apoplasto
Efficienza della conversione fotosintetica
dell’energia solare in energia chimica
Radiazione solare incidente (%)
Perdita di energia (%)
Riflessa e trasmessa (RT):
non PAR (IR e UV)
banda del verde
11.2
Radiazione solare incidente (%)
Perdita di energia (%)
Riflessa e trasmessa (RT):
- non PAR (IR e UV)
- banda del verde
- calore (fotoni blu)
11.2
Fotoni del blu-violetto hanno energia
doppia (300 kJ mole-1)
di quelli
del rosso (160 kJ mole-1)
ma la stessa efficacia fotosintetica
Il surplus di energia del fotone
blu-violetto viene rapidamente
dissipata come calore
Radiazione solare incidente (%)
Perdita di energia (%)
Riflessa e trasmessa (RT):
- non PAR (IR e UV)
- banda del verde
- calore (fotoni blu)
11.2
- limite termodinamico
Energia persa nel centro di
reazione per il trasferimento
dell’energia dalla Chl-trap
all’accettore primario
Radiazione solare incidente (%)
Perdita di energia (%)
Riflessa e trasmessa (RT):
- non PAR (IR e UV)
- banda del verde
- calore (fotoni blu)
11.2
- limite termodinamico
Consumo energetico per la
biosintesi carboidrati
C3: 3 ATP e 2 NADPH
C4: 5 ATP e 2 NADPH
Consumo energetico del ciclo C3
3 ATP
per fissare e ridurre
1 molecola di CO2
2 NADPH
Consumo energetico del metabolismo C4
Richiesta energetica per organicare 1 molecola di CO2:
2 NADPH
+
5 ATP
(3 + 2 da PPDK)
Le piante C4 sono energeticamente meno efficienti
delle piante C3 ?
No, perchè le C3 perdono una quota rilevante
di carbonio con la fotorespirazione
Radiazione solare incidente (%)
Perdita di energia (%)
Riflessa e trasmessa (RT):
- non PAR (IR e UV)
- banda del verde
- calore (fotoni blu)
11.2
- limite termodinamico
- Biosintesi carboidrati
- Fotorespirazione
- Respirazione
= c massimo teorico
C3 = 4.6 % ; C4 = 6.0 %
sono valori teorici
In realtà l’efficienza di conversione per le piante
C3 e C4 è più bassa
C3 = 2.4 % ; C4 = 3.7 %
Cosa è che abbassa l’efficienza di conversione (c )?
la foglia è incapace di utilizzare tutta la radiazione incidente
Identificazione delle diverse vie fotosintetiche: C3,
C4 e CAM
Misurando l’abbondanza nelle foglie
degli isotopi stabili del carbonio:
(Isotopi radioattivi del carbonio :
11C
12C
e
e
14C
13C
)
Il carbonio-13 (13C) è un isotopo non radioattivo del carbonio,
composto da 6 protoni e 7 neutroni.
Gli isotopi radioattivi
decadono nel tempo
Gli isotopi stabili di un elemento restano
costanti in abbondanza nel tempo
CO2 dell’atmosfera contiene gli isotopi del carbonio
con queste proporzioni
Le proprietà chimiche della 13CO2 e 12CO2 sono identiche
Durante la fotosintesi le piante discriminano nei
confronti dell’isotopo più pesante: 13CO2
Mass
44
45
46
Isotopologue
Abundance
s
12C16O
2
13C16O
2
12C18O16O
98.40%
1.19%
0.41%
Nei tessuti il rapporto isotopico si misura con lo spettrometro di massa
13CO
2
R=
12CO
2
Il rapporto isotopico del carbonio delle piante si misura con il: δ13C ‰
Standard di riferimento
Pee Dee Belemnite (PDB)
Rapporto isotopico misurato
nella calcite di un mollusco fossile
della Carolina del Sud (USA)
a cui è stato dato valore 0
δ13C ‰ della CO2 atmosferica = - 8 ‰
Significa che c’è meno 13C nella CO2 atmosferica
rispetto allo standard
E nelle piante ?
Le piante C3 hanno un valore medio = - 28 ‰
Le piante C4 hanno un valore medio = - 14 ‰
Quindi le piante C3 e C4 hanno meno
13C
della CO2 atmosferica
il che significa che i tessuti fogliari sono in grado di discriminare
per il 13C durante il processo fotosintetico
Sono stati determinati i valori di δ13C ‰ di un
gran numero di piante C3 e C4
Come appare chiaro dalla figura esiste una vasta
gamma di valori di δ13C ‰ per le piante C3 (da – 20 a – 37)
e per le piante C4 (da – 9 a – 17)
Queste variazioni riflettono i cambiamenti
di conduttanza stomatica all’interno
di ciascun gruppo di piante
Conduttanza stomatica ALTA
BASSA discriminazione
Conduttanza stomatica BASSA
ALTA discriminazione
Come si spiegano queste variazioni nel rapporto
isotopico del carbonio tra piante C3 e C4?
A) La 12CO2 è più leggera della 13CO2 e diffonde più
rapidamente verso il sito di carbossilazione.
B) La RUBISCO ha un valore intrinseco di discriminazione
contro la 13CO2 di – 30 ‰.
La PEPasi ha invece un effetto discriminatorio
inferiore – 2 ‰
Motivo principale
Nelle piante CAM obbligate il δ13C ‰ è lo stesso delle C4
Nelle piante CAM inducibili il δ13C ‰ è lo stesso delle C3