Patologia generale
Prof. Ezio Laconi
Lezione n°30 parte 2
08/01/2013
Beatrice Onali
In questa lezione parleremo di fisiologia dell’emopoiesi,di anemia mentre non parleremo delle
piastrinopenie.Per quanto riguarda i linfomi basti quello che abbiamo detto sulle neoplasie in generale(sono
argomenti dell’ematologia clinica) però se qualcuno vuole fare un seminario è benvenuto.
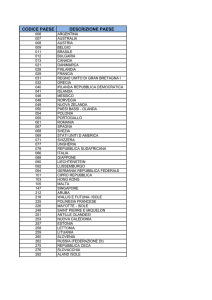
In questa tabella si possono osservare i riferimenti normali per i leucociti.
1 μl =1 mm³
1cm³=1 ml
Vedete c’è un range di normalità,non valori fissi.
Per quanto riguarda gli eritrociti ,di cui ci occuperemo prevalentemente parlando di anemie, avete tutta
un’altra serie di parametri e sigle che è opportuno fissare.
1
I parametri sono leggermente diversi tra uomo e donna.
I livelli di emoglobina sono un pochino più bassi per la donna rispetto all’uomo.
Alterazioni della dimensione:
Microciti :globuli rossi più piccoli, si hanno in alcune forme di anemia associate a carenza di ferro o
difetto di sintesi dell’emoglobina.
Macrociti:globuli rossi più grandi,li troviamo in anemie macrocitiche , sono dovuti a carenza di
vitamina B 12 o di folati.
Alterazioni della forma:
Sferociti:eritrociti a forma di sfera,li troviamo in anemie sferocitiche in cui è alterata la membrana
eritrocitaria. La forma a sfera è una forma normale per le cellule circolanti come i leucociti,invece
per gli eritrociti è una forma anomala, perché la loro forma normale è a lente biconcava. L’eritrocita
per mantenere questa forma biconcava deve avere una certa struttura. Alterazioni della struttura
per default fanno acquisire la forma a sfera.
Drepanociti:eritrociti a forma di falce,associati all’anemia falciforme.
Poichilociti:globuli rossi a forma di lacrima,anemie di diverso tipo. Non hanno la forma di una
lacrima classica,è dovuta a danni nel microcircolo.
2
Queste cellule viaggiano molto, vivono 120 giorni e sono soggette a traumi come nessun’ altro tipo di
cellula .Quindi nonostante sembrino “indifese”,hanno una sola membrana e non hanno nucleo, invece
devono essere molto resistenti ai traumi di natura fisica.
Le anemie sono caratterizzate da una diminuita concentrazione di emoglobina nel sangue a prescindere
dal fatto che i globuli rossi siano diminuiti o meno.
Nei maschi si parla di anemia a valori inferiori a 12.5-13.5 ,nelle femmine a valori inferiori a 11.5-12.5.
Anche qui c’è un range che dipende dall’età, dall’etnia ecc.
Nel concetto di malattia i limiti sono sfumati, bisogna avere del buon senso e vedere di volta in volta
quando intervenire e se è necessario intraprendere un percorso diagnostico.
Questo perché ,mentre l’esame dell’emoglobina si effettua con un semplice emocromo ,gli altri esami non
sono semplicissimi né economicissimi,quindi è una decisione importante.
Nel sesso femminile valori inferiori hanno delle implicazioni ulteriori.
L’eritropoiesi
Le cellule del sangue vengono generate a livello del midollo osseo,tutte le cellule circolanti,nello specifico i
globuli rossi,attraverso una serie di tappe ben studiate.
Il modello di una cellula staminale del midollo osseo è un “caposcuola” per i modelli di
differenziamento,quasi tutta la biologia del differenziamento cellulare ha preso l’avvio dallo studio di
questo modello.
3
La serie rossa inizia dal BFU E (burst forming unit) che continua nelle varie tappe differenziative . A livello
delle cellule CFU E compare il recettore per l’eritropoietina,che è uno degli ormoni o citochine che ha un
potere stimolatorio sull’eritropoiesi.
L’eritropoietina viene rilasciata prevalentemente a livello renale ed agisce su questi due tipi cellulari(nella
figura sono indicati CFU E e pronormoblasti) durante il processo di formazione degli eritrociti.
In seguito il recettore scompare nuovamente.
CFU E è a questo punto una cellula unipotente che darà luogo alla serie eritrocitaria (non solo l’eritrocita
avrà luogo da questa cellula).
Un’ulteriore fase maturativa darà origine al pro normoblasto ,il normoblasto basofilo, ancora qui c’è il
nucleo che occupa gran parte del volume cellulare.
Il normoblasto policromatofilo diventa normoblasto ortocromatofilo che smette di dividersi.
Da questo punto in poi la cellula subisce solo processi di differenziamento fino ad acquisire il fenotipo finale
dell’eritrocita.
Il reticolocita viene rilasciato nel sangue periferico,questa cellula ha nel suo citoplasma dei residui di
materiale citoplasmatico essenzialmente rna ,ribosomi ed organuli.
L’eritropoiesi è in condizioni normali inefficace per circa il 10 % ,ciò significa che non produce cellule
utilizzabili ed inviabili in periferia per svolgere il loro lavoro.
Questo fatto è motivato dall’estrema complessità del processo differenziativo che include l’eliminazione del
nucleo dalla cellula.
4
L’eritropoiesi inizia nel sacco vitellino, continua nel fegato e nella milza fino al sesto mese per poi prendere
l’avvio nel midollo osseo, che diventerà nella vita post natale ,la sede definitiva dell’eritropoiesi.
Secondo un’acquisizione di una 50ina di anni,l’eritropoiesi è organizzata nelle cosiddette isole
eritropoietiche.
Il numero delle isole determina l’intensità dell’eritropoiesi.
Sono stati fatti degli esperimenti, nei quali sono state messe a confronto tre situazioni: se si sottrae del
sangue a degli animali (simulando un’ emorragia) l’eritropoiesi viene stimolata ,se si trasfonde del sangue
ad un animale , l’eritropoiesi viene soppressa però non completamente, oppure si tiene l’animale in
condizioni normali non avremo alterazioni.
Quando si sottrae sangue l’eritropoiesi aumenta ed aumentano le isole eritropoietiche.
Queste isole sono costituite da cellule chiamate Nurse(bambinaia),una cellula che si prende cura degli
eritrociti che si stanno formando e li segue nel loro percorso maturativo.
Questa cellula è ritenuta un macrofago,in realtà il suo fenotipo non lo conosciamo molto bene,si tratta di
cellule derivate dalla serie mieloide del midollo.
Chiamiamo queste cellule macrofagi anche se la loro unica funzione è quella di aiutare l’eritropoiesi.
Non sappiamo la loro funzione ben precisa,aiutano soprattutto la parte intermedia del processo
maturativo.
Sappiamo che le isole eritropoietiche non sono presenti negli uccelli e i globuli rossi degli uccelli sono
nucleati.
5
Questo ci da un’indicazione su almeno una funzione delle isole ,quella di aiutare nel processo di
enucleazione dei globuli rossi.
Gli eritroblasti prima di entrare nei sinusoidi del midollo emettono l’intero nucleo,che viene
immediatamente fagocitato proprio dai macrofagi.
Questo processo è estremamente controllato,produce
2 000 000 di eritrociti al secondo,quindi è un processo che va avanti a ritmi elevatissimi.
L’attività midollare da questi livelli può aumentare ,in condizioni di maggiore richiesta funzionale fino a 8-10
volte ,qualcuno parla anche di 15 volte.
E’ importante che venga ripulito il microambiente midollare da questi detriti (i nuclei degli
eritroblasti),pieni di DNA che può diventare uno stimolo per il processo infiammatorio.
Questi nuclei hanno sulla loro membrana la fosfatidilserina che è un segnale di “mangiami” per il fagocita.
Il fagocita deve digerire il DNA infatti è ricchissimo di DNAasi. Se c’è un deficit della DNAasi (forse di tipo 2)
il processo dell’eritropoiesi si blocca.
Eritroblasti e macrofagi si inviano segnali reciproci attraverso recettori e ligandi che sono molto importanti
(alcuni dei quali vedete illustrati, ma non sono da sapere i nomi).Mancando qualcuna di queste componenti
viene messo in crisi l’intero processo.
Le cellule Nurse seguono per un lungo periodo gli eritroblasti. Si è visto che le isole di eritropoiesi più vicine
ai sinusoidi ( più vicine al sito in cui questi reticolo citi dovranno essere immessi) hanno degli eritroblasti più
vicini alla fase terminale di maturazione.
6
Questo fatto ha suggerito due ipotesi:
Le isole si muovono da zone più lontane ai sinusoidi a zone più vicine,mano a mano che gli eritrociti
che hanno deciso di seguire maturano.
Oppure se pensiamo che le isole non si muovano saranno gli eritroblasti a spostarsi da un’isola
all’altra ,mano a mano che acquisiranno caratteristiche di maturazione più avanzate.
Le due ipotesi sono ancora entrambe valide perché non è stata trovata ancora quella corretta,possono
essere tutte e due valide.
Per quanto riguarda il processo di eliminazione degli organuli citoplasmatici è stato considerato sia il
fenomeno di esocitosi sia il fenomeno di autofagia.
L’autofagia è l’autodigestione da parte della stessa cellula di organuli endocellulari,importante per la
maturazione finale dei globuli rossi perché alterazioni di questo processo sono associate ad alterazioni
dell’eritropoiesi.
Nell’eritropoiesi intervengono fenomeni di regolazione positiva(che l’aumentano) e fenomeni di
regolazione negativa (che la diminuiscono).
Più un processo è veloce,come questo,più necessita di essere controllato. Se voi state guidando una
macchina a 100 km/h con il volante dovete stare molto più attenti che quando andate a 10km/h.
Un errore anche dell’1% può portare un incremento di cellule (che erano 2000 000/s) di 40 000 cellule in
più al secondo che se moltiplicate per un giorno diventano parecchi miliardi di cellule in più. Quindi è un
fenomeno che deve essere finemente controllato.
Regolazione positiva
7
Tra i regolatori positivi un ruolo fondamentale ha l’eritropoietina che è un ormone indotto
dall’ipossia(situazione in cui l’ossigeno è diminuito) non è l’unico ma è uno dei principali,il mediatore è
sempre l’HIF(Hypoxia inducible factor).
L’eritropoietina è prodotta soprattutto dal rene ma anche dal fegato e dal SNC.
In condizioni d’ insufficienza renale possiamo avere anemia anche legata al fatto che viene prodotta meno
eritropoietina.
L’eritropoietina stimola la proliferazione e protegge i pro eritroblasti dall’ apoptosi indotta dagli stessi
eritroblasti maturi.
Gli eritroblasti maturi regolano la produzione dei loro precursori (proeritroblasti).
Queste cellule nascono con una sorta di “marchio” ,la possibilità di essere eliminate.
I pro eritroblasti posseggono sulla loro superficie il recettore per il Fas ligando ,dei recettori che
opportunamente stimolati possono portare queste cellule ad apoptosi.
Sono gli eritroblasti a mandare i segnali di apoptosi.
In assenza di macrofagi gli eritroblasti in vitro proliferano molto poco ,se aggiungiamo altri macrofagi si ha
un aumento fino a 3 volte dell’eritropoiesi in vitro. Inoltre forniscono un supporto meccanico importante
per la trasmissione dei segnali.
Adesione, grado di tensione tra matrice e cellule stesse si è visto ultimamente che sono fondamentali anche
per le neoplasie.
Il ruolo dei messaggi chimici anche da parte dei macrofagi fu ipotizzato dallo stesso studioso che descrisse
le isole eritropoietiche e che diede il nome di cellule Nurse,il quale ipotizzò che i macrofagi fossero anche i
donatori di ferro.
Il ferro è un elemento fondamentale per l’eritropoiesi poiché viene utilizzato dagli eritrociti per la
produzione dell’emoglobina ,della quale sono ricchi.
Il macrofago secerne(secondo questa ipotesi) una proteina, la ferritina,che lega il ferro,questa sarebbe poi
captata dagli eritroblasti per la cattura del ferro e la sintesi dell’emoglobina.
Le anemie sono classificate anche in quelle che rispondono all’eritropoietina e in quelle che non
rispondono.
Se non funziona il meccanismo del passaggio del ferro anche dando molta eritropoietina l’anemia
continuerà ad esistere. Questo può succedere anche durante processi di infiammazione cronica e neoplasie
,l’IL6 può essere responsabile di queste anemie.
Fattori solubili secreti da macrofagi ed eritroblasti,ad esempio la proteina Gas 6 (growth arrest specific 6)
può essere un risvolto terapeutico poiché stimolerebbe l’eritropoiesi ,favorendo il contatto tra macrofagi
ed eritroblasti,potrebbe bypassare l’effetto dell’eritropoietina in anemie che non rispondono a questa.
8