Appendice online
Approfondimenti: la maturazione del follicolo ovarico
Se seguiamo lo sviluppo dell’unico follicolo che arriverà all’ovulazione, osserviamo che questo passa da
follicolo primordiale a follicolo primario (Fig. 1App.), a follicolo secondario (Fig. 2App.) cui
corrispondono ben 7 classi (dalla II^ alla VIII^) in base alle dimensioni via via crescenti che culminano
con il cosiddetto follicolo maturo o di Graaf (Fig. 3App. e Fig. 12 a pag. 8 delle dispense di
Embriologia). Quest’ultimo è la forma più sviluppata di follicolo di classe VIII. L’intero processo ha
una durata di circa un anno.
Fig. 1App. Follicolo primordiale e follicolo primario.
Fig. 2App. Follicolo secondario.
Fig. 3App. Follicolo di Graaf. Il follicolo arriverà a misurare
circa 20 mm e l’ovocito 100-150 µm
antro
follicolare
ovocito con
zona pellucida
Il follicolo primordiale è il tipo di follicolo predominante nell’ovaio ed è il più piccolo, avendo un
diametro di 25 µm. Esso è circondato da cellule follicolari appiattite (Fig. 1App.).
Gli ovociti dei follicoli primordiali si trovano in fase di arresto in diplotène della profase della prima
divisione meiotica fin dal periodo fetale. I follicoli primordiali che lasciano la fase di arresto, con la
ripresa dell’ovogenesi, diventano (in circa 150 giorni) follicoli primari. All’inizio il follicolo primario è
monostratificato (unilaminare), con uno strato singolo di cellule follicolari cuboidali (Fig. 1App.).
Queste, proliferando e stratificandosi, lo trasformano in pluristratificato (multilaminare) e verranno
chiamate anche cellule della granulosa (Fig. 1App.). Il follicolo primario cresce tra i 60 fino ai 100
µm di diametro, con l’ovocito che si accresce fino a 70 µm di diametro. Una lamina basale separa il
follicolo primario dallo stroma (connettivo e vasi) ovarico. Nel follicolo primario, l’ovocito primario
inizia a sintetizzare un rivestimento glicoproteico, la zona pellucida, che progressivamente separerà le
cellule follicolari dall’ovocito. La zona pellucida è attraversata da sottili prolungamenti citoplasmatici
delle cellule follicolari in contatto con microvilli dell’ovocito. Le zone di contatto sono sede di
giunzioni gap, attraverso le quali cellule follicolari e ovocito possono comunicare tra loro. Al follicolo
primario occorrono 4 mesi per raggiungere lo stadio di follicolo secondario di classe I (intorno ai
120-200 µm di diametro) e completare la fase cosiddetta pre-antrale perché non si è formato ancora
l’antro (vedi avanti). A questo stadio, il follicolo secondario va incontro a proliferazione continua
delle cellule follicolari ed ispessimento della zona pellucida. L’ovocito cresce ancora (ora siamo
intorno ai 100 µm di diametro). Le cellule stromali che circondano il follicolo vanno a disporsi a
formare una capsula cellulare, la teca. La teca si differenzia presto in due strati: 1. teca interna,
cellulare e 2. teca esterna, fibrosa. La teca interna, uno strato ben vascolarizzato adiacente alla lamina
basale del follicolo in sviluppo, è formata da cellule di natura connettivale ma ad attività endocrina. Tali
cellule, infatti, cominciano ad esprimere recettori per l’ormone luteinizzante (LH) ipofisario che le
stimola a produrre ormoni androgeni. Questi vengono traslocati alle adiacenti cellule della granulosa
che provvedono a trasformare gli androgeni in estrogeni stimolate dai recettori per l’altro importante
ormone ipofisario, l’ormone follicolo-stimolante (FSH). La teca esterna è uno strato di tessuto
connettivo simile a una capsula in continuità con lo stroma ovarico. Ben presto il follicolo secondario,
divenuto capace di rispondere agli stimoli ipofisari ed ora in grado di produrre ormoni, entra nella fase
antrale dove subisce una più veloce maturazione in cui si riconoscono 7 classi (dal follicolo secondario
di classe II fino a quello di classe VIII). Vediamo ora perché si chiama fase antrale: piccoli spazi
intercellulari contenenti fluido follicolare compaiono tra le cellule follicolari (Fig. 2App.), e
confluiranno in seguito per formare un ampio spazio, l’antro (Fig. 3 App.), entro il quale sporgerà
l’ovocito contornato da cellule della granulosa. Il follicolo secondario di classe VIII raggiungerà le
dimensioni di 20 mm in circa due mesi, che è la durata della fase antrale. Un solo follicolo di classe
VIII, infine, diventa follicolo maturo o di Graaf o preovulatorio, raggiungendo le dimensioni
massime pari a 25 mm (Fig. 3 App. e Fig. 12, pag. 8 delle dispense di Embriologia). Un follicolo
di Graaf è caratterizzato da un grande antro (che contiene fluido follicolare) in cui sporge l’ovocito,
circondato dalla zona pellucida, sormontata da un singolo strato di cellule follicolari che formano la
corona radiata (Fig. 4App.). A sua volta, l’ovocito è attaccato eccentricamente alla parete del follicolo
tramite un gruppo di cellule follicolari, chiamato cumulo ooforo (Fig. 4App.).
Di tutti i follicoli che ogni giorno iniziano a
maturare, solo dai 3 ai 10 follicoli antrali al mese
arrivano al punto di poter rispondere all’FSH ma,
come già detto, solo uno (il follicolo dominante o
di Graaf) giunge alla ovulazione, perché si trova ad
avere i recettori per gli ormoni ipofisari (FSH ed
LH) al momento giusto. Tutti gli altri follicoli
degenerano (atresìa) in ogni momento del loro
sviluppo.
Fig. 4App. Follicolo di Graaf.
Il ciclo ovarico (per un riassunto, vedi Fig. 8App.)
Tenendo bene in mente che occorre circa un anno affinché il follicolo primordiale riesca ad arrivare alla
ovulazione, tuttavia si può considerare un ciclo ovarico in parallelo al ciclo mestruale e di pari durata
(28±7,5 giorni). Il ciclo ovarico si può dividere in tre fasi: 1) fase follicolare; 2) fase ovulatoria; 3)
fase luteinica.
La fase follicolare. E’ la fase di circa 14 giorni che precede l’ovulazione, corrisponde al periodo
compreso tra il primo giorno di mestruazione e l’ovulazione stessa ed ovviamente, comprende l’ultimo
periodo di sviluppo del follicolo destinato ad ovulare. In fase follicolare precoce (durata 4 giorni),
l’evento fondamentale è la selezione di 1-5 follicoli secondari (che dalla classe V diventano follicoli di
classe VI). Essi evitano l’atresia forse perché le cellule della granulosa si dotano di un maggior numero
di recettori per FSH ed LH ed inoltre aumenta la vascolarizzazione della teca interna.
Nella fase follicolare intermedia (durata 8 giorni) si ha la dominanza precoce durante la quale i pochi
follicoli di classe VI diventano di classe VII e poi tutti, tranne uno, vanno incontro ad atresia. Tale
follicolo, definito dominante, diventerà di classe VIII.
Nella fase follicolare tardiva (pre-ovulatoria, durata 2 giorni) si ha la dominanza tardiva, in cui il
follicolo dominante di classe VIII diventa follicolo maturo (del diametro di 25 mm).
Nel follicolo maturo si assiste ai seguenti eventi:
1) distacco dell’ovocito e della relativa corona radiata dal cumulo ooforo, così che il complesso
ovocito/zona pellucida/corona radiata galleggia libero nel fluido follicolare;
2) completamento della prima divisione meiotica (diverse ore prima della ovulazione), con la
formazione di un ovocito secondario e del primo globulo polare, che rimane nello spazio compreso
tra la zona pellucida e l’ovocito (spazio perivitellino); la ripresa della meiosi nell’ovocito necessita della
produzione di progesterone da parte delle cellule della granulosa che iniziano a secernere tale ormone
poche ore prima dell’ovulazione. Subito, l’ovocito secondario inizia la seconda divisione meiotica che
prosegue fino ad arrestarsi di nuovo alla metafase. La meiosi si completerà solo in caso di fecondazione.
3) le cellule follicolari della granulosa esprimono i recettori per l’ormone LH (luteinizzante)
oltre ai già presenti recettori per l’ormone FSH (follicolo-stimolante).
Questo evento induce il follicolo a secernere progesterone ed è critico per lo sviluppo del corpo luteo
(vedi avanti).
La fase ovulatoria
Alla ovulazione, il follicolo maturo sporge alla superficie dell’ovaio formando lo stigma (Fig. 5App.). Il
picco di LH provoca l’induzione di una intensa attività di digestione enzimatica delle proteine della teca
esterna e della tunica albuginea. Così il follicolo di Graaf maturo si apre ed il gamete liberato entra nella
tuba uterina o ovidutto, molto vicino alla gonade. Poche ore dopo l’ovulazione, le cellule follicolari e la teca interna, rimaste nell’ovaio, iniziano a trasformarsi in corpo luteo.
epitelio di rivestimento
Fig. 5 App.
zona pellucida
corona radiata
stigma
ovocito secondario
globulo polare
lo strato delle cellule follicolari inizia
a ripiegarsi subito dopo l’ovulazione
la membrana basale si rompe e vasi della teca
interna invadono la granulosa. La cavità dell’antro si
riempie di sangue (corpo emorragico).
La fase luteinica: il corpo luteo
Dopo l’ovulazione, lo strato di cellule follicolari residue collassa e diviene parte del corpo luteo (Fig. 6
App.), una grossa ghiandola a secrezione ormonale.
vasi nel
corpo luteo
cellule
granulosaluteiniche
cellule tecaluteiniche
teca
esterna
Fig. 6App. Formazione del corpo luteo. A destra, cellule granulosa-luteiniche e cellule tecaluteiniche all’interno del corpo luteo.
Questa trasformazione è possibile dopo che è avvenuta:
1. La rottura della membrana basale del follicolo.
2. La formazione del coagulo e l’invasione di vasi sanguigni entro la massa di cellule follicolari dove
prima i vasi erano assenti. Nel coagulo penetrano poi vasi neoformati, fibroblasti e fibre collagene.
3. La trasformazione delle cellule follicolari e della teca interna (Fig. 6App.). Le cellule follicolari
della granulosa si trasformano in cellule granulosa-luteiniche e quelle della teca interna in cellule
teca-luteiniche. Il corpo luteo secerne estrogeni e progesterone stimolato dagli ormoni ipofisari FSH
ed LH e continua ad ingrandirsi per poi iniziare la fase involutiva circa 14 giorni dopo l’ovulazione, a
meno che non si abbia la fecondazione dell’ovocito ovulato. Se ciò avviene, il trofoblasto
dell’embrione impiantato inizia a secernere un ormone glicoproteico, la gonadotropina corionica
umana (hCG), che stimola il corpo luteo ad ingrandirsi ed a produrre progesterone ed estrogeni,
necessari per mantenere l’endometrio, prolungando la vita del corpo luteo almeno fino alla 7^ settimana
di gestazione. La placenta, la corteccia surrenale fetale e il fegato sostituiranno nella sua funzione il
corpo luteo producendo gli ormoni steroidei necessari a mantenere la gravidanza. La regressione del
corpo luteo (luteolisi) porta alla formazione del corpus albicans dove il connettivo stromale ha
sostituito le cellule in degenerazione. Il corpus albicans resta nell’ovaio; esso diminuisce in dimensioni ma
non scompare mai.
Per inciso, va detto che verso la fine della fase luteinica (ultimi 6 giorni), quando la caduta di
estrogeni e di progesterone stimola l’aumento di FSH ed LH, ha luogo il reclutamento dei follicoli
che andranno a costituire il pool di follicoli tra cui sarà scelto quello destinato ad ovulare nel
successivo ciclo ovarico. Durante il reclutamento, quei follicoli di classe IV che non vanno in
atresia maturano a follicoli di classe V tra i quali avverrà la selezione di cui abbiamo parlato.
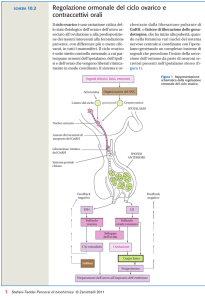
Regolazione ormonale della ovulazione e del corpo luteo
Due ormoni dell’ipofisi anteriore regolano la crescita follicolare (Fig. 7App.):
1. L’ormone follicolo-stimolante (FSH) che stimola la follicologenesi, l’ovulazione e la produzione di
estrogeni;
2. L’ormone luteinizzante (LH) che, con un’impennata, induce l’ovulazione.
La secrezione continua di LH induce la luteinizzazione dello strato di cellule follicolari che residuano
dopo l’ovulazione e poi stimola la secrezione di progesterone da parte del corpo luteo. I livelli elevati di
progesterone ed estrogeni nella fase luteinica inibiscono l’ipofisi e la produzione di FSH ed LH cessa.
Pertanto, nella fase luteinica del ciclo ovarico non è possibile che altri follicoli maturino fino alla
ovulazione. Questo meccanismo è utilizzato nella cosiddetta pillola anticoncezionale che, appunto, è
una miscela di progesterone d estrogeni che agisce come un corpo luteo artificiale. La diminuzione di
LH contribuisce a far degenerare il corpo luteo. All’inizio della mestruazione, i livelli di estrogeni e di
progesterone sono bassi e crescono gradualmente durante il periodo pre-ovulatorio.
Fig. 7App. Il ciclo
ovarico.
Gli estrogeni raggiungono i massimi livelli appena prima che il picco di LH preceda l’ovulazione. La
sintesi FSH-dipendente di estrogeni da parte delle cellule follicolari stimola la proliferazione delle
ghiandole dell’endometrio. La sintesi LH-dipendente di progesterone da parte del corpo luteo inizia
e mantiene l’attività secretoria delle ghiandole dell’endometrio. La Fig. 8App. riassume il ciclo
ovarico.
Fig. 8App. Riassunto del ciclo ovarico.
Dal follicolo primordiale si passa a quello primario, secondario ed, infine, a quello maturo che ovulerà liberando
l’ovocito maturo contornato dalla zona pellucida e dalle cellule della corona radiata. Ciò che resta del follicolo
dopo l’ovulazione (le cellule della granulosa e della teca interna) subirà l’invasione di vasi e tessuto connettivo
trasformandosi in corpo luteo, una nuova ghiandola a secrezione steroidea (produce progesterone ed estrogeni),
vitale per 14 giorni, dopo di che si trasformerà in corpus albicans, una struttura cicatriziale che non scomparirà mai
dall’ovaio; in caso di fecondazione, il corpo luteo (detto corpo luteo gravidico) sopravvivrà per diverse settimane
dopo di che cesserà di funzionare, sostituito dalla placenta che già alla sesta settimana inizia a produrre il
progesterone sufficiente a mantenere la gravidanza.
Nel 6-7 % delle gravidanze possono manifestarsi perdite ematiche (anche a carattere ritmico fino alla sedicesima
settimana) che possono essere scambiate per una regolare mestruazione. Varie le cause, come l’annidamento
dell’embrione o anomalie della gravidanza o dell’apparato genitale.
Il progesterone è necessario a sostenere la gravidanza fino al parto perché impedisce la mestruazione e mantiene
vitale l’endometrio (che si trasforma in decidua durante la gravidanza). Il farmaco RU-486 (pillola abortiva) agisce
come antagonista del progesterone legandosi ai suoi recettori ed impedendogli di funzionare comportandosi così
come farmaco abortivo.