LEZIONE DI FISIOLOGIA DEL 08/04/2013
Francesca Parisi
FISIOLOGIA DELLA RIPRODUZIONE
Vorrei terminare la fisiologia delle ghiandole endocrine. Dobbiamo parlare delle gonadi e anche degli
ormoni che intervengono nell’omeostasi calcica, cioè paratormone, tireocalcitonina e ormone D, ma
non credo di aver tempo di parlare di questo capitolo.
Il discorso riguardante le gonadi è un processo molto descrittivo, perché le gonadi fanno parte del
processo della riproduzione dunque cercherò di dare qualche cenno della fisiologia della riproduzione
che comporta:
-lo studio degli organi della riproduzione;
-dei meccanismi della differenziazione sessuale;
-funzione delle gonadi, testicoli e ovaie;
-funzione delle vie genitali;
-la regolazione ormonale delle gonadi.
Si distinguono gli organi sessuali primari e secondari che insieme costituiscono i caratteri sessuali
primari, dunque non confondete il concetto di organo sessuale con carattere sessuale.
Gli organi sessuali primari sono ovviamente le gonadi, che hanno una duplice funzione:
-una gametogentica, che potremo definire esocrina;
- e una endocrina che è appunto la produzione di ormoni.
La funzione esocrina assicura il mantenimento della specie, mentre la funzione endocrina assicura il
mantenimento della funzione riproduttiva.
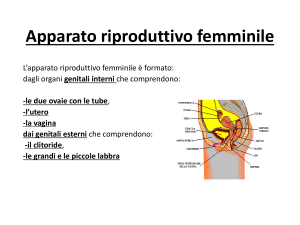
Gli organi sessuali secondari sono le vie genitali interne ed esterne. Nel maschio, gli organi genitali
interni sono l’epididimo, le vie spermatiche, le veschichette seminali, la prostata e le ghiandole
bulbouretrali. Gli organi genitali esterni sono il pene e lo scroto.
Nella femmina, gli organi genitali interni sono le tube di Falloppio, l’utero, la vagina e le ghiandole
annesse. Quelli esterni sono invece la cervice e le ghiandole mammarie.
I caratteri sessuali secondari che contribuiscono alla differenziazione del sesso e che dipendono dagli
ormoni prodotti dalle gonadi sono:
-la distribuzione del sistema pilifero;
-lo sviluppo della massa muscolare, maggiore nel maschio teoricamente;
-il maggiore o minore sviluppo dell’accumulo di grasso, maggiore nella donna;
-e infine la forma di alcune aree come il bacino e i cingoli scapolari diversi in lunghezza.
DIFFERENZAZIONE SESSUALE
Si fa riferimento al:
-Sesso genetico
-sesso gonadico, cioè differenziazione delle gonadi,
-fenotipo sessuale o sesso genitale, che consiste nella differenziazione degli organi genitali interni ed
esterni;
-differenziazione del cervello sessuale.
Il sesso genetico dipende dalla combinazione dei cromosomi sessuali . Se si forma un embrione con
cromosomi sessuali X Y si ha un soggetto di sesso maschile. Ovviamente la mancanza del cromosoma
Y orienterà verso il sesso genetico femminile. Ciò è importante perchè se il soggetto è geneticamente
maschio il gene del cromosoma Y codifica per un fattore determinante per i testicoli( TDF in sigla),
cioè un fattore che orienta la gonade primitiva verso il testicolo. Se invece manca il cromosoma Y, e
non viene prodotto di conseguenza questo fattore, non si svilupperà la gonade maschile, bensì l’ovaio.
Il TDF dirige dunque la differenziazione delle gonadi in testicoli. È una proteina.
I testicoli produrranno il testosterone e porteranno altri fattori che provvederanno al sesso fenotipico.
Immaginate una gonade X Y , quindi si differenzierà in testicolo che produrrà testosterone che
ovviamente stimolerà il dotto di Wolff. Contemporaneamente al testosterone, che è prodotto dalle
cellule del Leyding, le cellule del Sertoli produrranno un fattore inibente il dotto di Muller che andrà
incontro a regressione. Vedremo che poi questo dotto di Muller servirà invece alla differenziazione
delle vie genitali femminili.
Dunque il testicolo produce testosterone che agisce sul dotto di Wolff che permette la differenziazione
delle vie genitali interne ed esterne in senso maschile quindi il testicolo, l’epididimo, le veschichette
seminali, i vasi deferenti e il dotto eiaculatore.
Nel contempo il testosterone che passa in circolo verrà trasformato dalla reduttasi in
diidrotestosterone che agirà sull’abbozzo genitale embrionale e permetterà lo sviluppo e
differenziazione della prostata, del pene e dell’uretra.
Quindi abbiamo il dotto di Wolff che porterà alla differenziazione di queste componenti morfologiche e
invece il testosterone, o meglio il diidrotestosterone, che agisce sull’abbozzo genitale embrionale.
Se invece la gonade è differenziata in senso femminile, l’assenza del testosterone farà regredire il dotto
di Wolff. Inoltre non venendo prodotto dalle cellule del Sertoli il fattore inibente il dotto di Muller,
questo potrà agire e differenziare.
L’assenza di testosterone potrà agire sull’abbozzo embrionale producendo alcune componenti come il
clitoride, le piccole e grandi labbra e il seno urogenitale. Invece per quanto riguardo il dotto di Muller
si avranno le tube di Falloppio, l’utero, la vagina nella sua parte superiore.
Nel maschio le cellule germinali produrranno gli spermatogoni, l’epitelio celomico produrrà le cellule
del Sertoli, e le cellule mesenchimali produrranno le cellule di Leyding.
Nella femmina le cellule germinali saranno gli ovogoni, l’epitelo celomico produrrà le cellule della
granulosa e le cellule mesenchimali daranno vita alle cellule tecali.
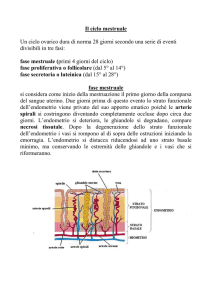
CICLO MESTRUALE
La funzione riproduttiva ha un ritmo circadiano, circa mensile e si chiama ciclo mestruale. Questo
rappresenta l’evento fondamentale per l’attuarsi della funzione riproduttiva e non si deve confondere
con le mestruazioni. Questo lo dico perché ci sono studenti che identificano il ciclo mestruale con il
flusso mestruale.
Il ciclo mestruale è il complesso degli eventi che porteranno, a un ritmo medio di 28 giorni, alla
produzione di un'ovocellula fecondabile e un terreno endometriale pronto all’eventuale annidamento
dell’uovo fecondato. Se non è avvenuta la fecondazione si ha la perdita dell’ovocellula e
dell’endometrio secretivo e dunque si ha il flusso mestruale.
Quali sono i momenti del ciclo mestruale?
Abbiamo innanzitutto una componente a carico del sistema nervoso, in questo caso lo possiamo
identificare con l’ipotalamo, l’ipofisi o meglio adenoipofisi e le ovaie.
La componente ipotalamica è correlata alla produzione di una liberina per due ormoni e non due
liberine per due ormoni, cioè l’ormone follicolo stimolante e luteinizzante. L’adenoipofisi produrrà due
gonadotropine LH e FSH.
Le ovaie hanno funzione gametogenetiche ed endocrine. Producono gli ormoni sessuali femminili che
sono l’estradiolo, e l’estrone in modo particolare, il progesterone e anche gli androgeni.
L’ipotalamo è un generatore e coordinatore di segnali. L’adenoipofisi traduce questi segnali in eventi
che porteranno all’attività delle ovaie. Quest’ultime saranno capaci di generare segnali a feedback che
poi sono responsabili del ciclo mestruale. Dunque chi detta il ciclo mestruale sono le ovaie, anche se
sotto il controlla dell’ipotalamo e dell'ipofisi. Qui abbiamo un feedback particolarissimo perché
solitamente in tutte le ghiandole abbiamo feedback esclusivamente negativi, qui invece avremo
feedback positivi e negativi in rapporto al periodo del ciclo mestruale.
La durata è di 28 giorni in media, nel ritmo fisiologico e inizia a partire dal primo giorno della
mestruazione precedente. Ovviamente in condizioni fisiologiche ogni ciclo è ovulatorio cioè produce
un ovocita pronto alla fecondazione.
Di norma, come fanno i ginecologi, si indica con 0 il giorno del picco pre-ovulatorio di LH che provoca
l’ovulazione, dunque coincide in pratica con l’ovulazione. Con +1, +2 e così via si indicano i giorni che
seguono e con -1,-2 i giorni che lo precedono.
Distinguiamo una fase pre-ovulatorio o follicolare e una fase post-ovulatoria o luteale. La prima, la fase
follicolare consiste nella maturazione del follicolo. La seconda invece consiste nella formazione e
attività del corpo luteo.
La fase pre- ovulatoria o follicolare va dal giorno M2 del ciclo al giorno -1 invece la fase postovulatoria dal giorno +1 al giorno M1, dove M significa il giorno dell’inizio della mestruazione del
ciclo precedente.
La fase follicolare è la più variabile mentre la fase luteale è la più costante. Se fosse stato l’inverso
avremmo potuto facilmente capire il momento dell’ovulazione.
Qui nello schema, si indica il ruolo dell’ipotalamo ed è come se esistessero due fattori liberanti invece
è sbagliato perché ne esiste solo una gonadoliberina, sia per FSH che per l’ LH.
L’ipofisi invece produce due gonadotropine, l’ FSH e l’LH. Ricordate che ne esiste una terza di origine
trofoblastica e non ipofisaria ed è la gonadotropina corionica, molto importante per il mantenimento del
corpo luteo nel corso della gravidanza.
Poi abbiamo l’ovaio che segnala a feedback perché si possa avere la fase pre-ovulatoria, il picco preovulatorio e la fase post- ovulatoria. Qui si studiano le modificazioni morfogenetiche che
consisteranno:
-nella maturazione follicolare,
-nel processo dell’ovulazione,
-nella formazione del corpo luteo
-le modificazioni funzionali che produrranno gli ormoni steroidei, cioè estrogeni, progesterone e
androgeni.
Poi abbiamo le tube dove avviene il processo dell’eventuale fecondazione. Anche queste vanno
incontro a modifiche cicliche, a livello sia della muscolatura liscia sia a livello dell’epitelio tubarico.
Poi abbiamo le classiche funzioni a livello dell’endometrio che sono la fase proliferativa o prima fase
che è la fase estrogenica, e la fase secretiva post-ovulatoria che è la fase luteale che dipende dal
progesterone.
Io aggiungerei che durante il ciclo mestruale ci sono modificazione morfologiche e funzionali a carico
del miometrio. Anche qui abbiamo una fase estrogenica che comporterà un aumento del trofismo del
miometrio e della sua eccitabilità. Invece durante la fase post – ovulatoria o progestinica gli effetti sul
miometrio saranno più che morfologico, che qui ci pensano gli estrogeni, ma riguarderà la riduzione
dell’eccitabilità del miometrio.
Saranno importanti le modificazione cicliche a carico della cervice, soprattutto del muco cervicale che
avrà caratteristiche chimico-fisiche diverse in fase estrogenica e progestinica.
E poi le modificazioni a carico della vagina dove si hanno modifiche cicliche a carico dell’epitelio
vaginale che tende a cheratinizzarsi.
Vedete che pur variando la durata del ciclo, la fase che permane pressochè costante è la fase luteale
mentre varia quella follicolare.
CICLO OVARICO
Adesso parlerò:
-dell’aspetto morfologico dell’ovaie,
-la biologia dell’ovogenesi, lo sviluppo dei follicoli ovarici che possiamo dividere in 3 parti,
-il ruolo degli ormoni ovarici,
-la regolazione ormonale nelle fasi dello sviluppo dei follicoli,
-il profilo temporale della secrezione ormanale durante il ciclo mestruale
-i meccanismi a feedback.
Fermo restando di quelle modificazioni a carico delle vie genitali importanti per il processo della
fertilizzazione della donna, dunqueun’ eventuale fecondazione e gestazione.
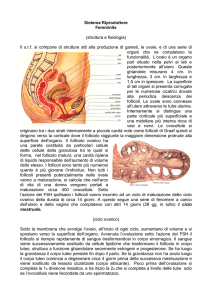
Il ciclo ovarico è caratterizzato da modificazioni morfologiche e funzionali che si manifestano a carico
dell’unità fondamentale riproduttiva dell’ovaio che è il follicolo ovarico che comprende le cellule della
granulosa, le cellule tecali con funzione endocrina e secretiva, e l’ovocita.
Gli aspetti morfologici: se prendiamo l’ovaio distinguiamo una parte corticale ,più rappresentata
rispetto al testicolo, la parte midollare che invece è meno rappresentata rispetto al testicolo, il tessuto
stromale e le cellule dell’ilo.
Se noi prendiamo la corticale vedremo i follicoli nei diversi gradi di maturazione: dal folliclo
primordiale, a quella primario, al follicolo secondario con formazione dell’antro, al follicolo maturo o
dominate o follicolo di Graff, la formazione del corpo luteo e quindi la regressione del corpo luteo se
non è avvenuta la fecondazione e quindi si forma il corpo albicante e poi troveremo anche dei follicoli
atresici.
Lo stroma è interposto tra i follicoli ed è importante perché dallo stroma si formeranno le cellule tecali.
Quindi ricapitolando nella corticale troveremo i follicoli ovarici che qui ho indicato come:
-follicoli primordiali
-follicoli primari o solidi
-follicoli secondari o cavi
-follicolo maturo o dominante o di Graff
-follicoli atresici e gli organi tecali
-corpi lutei
-corpi albicanti
Poi abbiamo la midollare che è un tessuto connettivo
lasso ricco di vasi.
Il tessuto stromale che è interposto tra i follicoli. E poi si hanno le cellule dell’ilo.
Tessuto stromale e cellule dell’ilo avranno attività endocrina perché produrranno gli androgeni.
Il follicolo di Graff o maturo è costitutioda un antro, una cavità follicolare con liquor follicoli, l’epitelo
pluristratificato, la membrana granulosa, il cumulo ooforo, la teca interna ed esterna.
OVOGENESI
Inizia durante la 4-5 settimana fetale quando le cellule embrionali si vanno a depositare lungo la cresta
genitale. Qui vanno incontro a mitosi e si trasformano in ovogoni e vanno incontro a mitosi. Durante la
gravidanza inizia la meiosi che però è incompleta. Prima avremo la proliferazione mitotica e poi inizia
la profase della meiosi e si ferma qui a livello di ovociti primari. È importante che durante la 20-25
settimana già i due terzi degli ovogoni si trasformati in ovociti primari e si fermano in profase della
meiosi I. Però sei mesi dopo la gravidanza tutti gli ovogoni si trasformano in ovociti primari. Nei
maschi gli spermatogoni corrispondono agli ovogoni, maturano e proliferano fino alla morte
dell’individuo, invece gli ovogoni rimarranno fino a metà della vita fetale e poi si avranno gli ovociti
primari. Quindi alla fine, la proliferazione degli ovogoni è tipicamente fetale e poi cessa e al 6 mese
dopo la gravidanza tutti gli ovogoni si sono trasformati in ovociti primari e poi alla pubertà riprende il
processo meiotico e durante l’ovulazione si completerà la prima meiosi e l’ovocita primario si è
trasformato in ovocita secondario. La seconda meiosi si avrà soltanto con la fecondazione.
Nella femmina la cellula germinale primordiale si trasforma in ovogonio e quindi abbiamo la mitosi
embrionale poi inizia la meiosi e si ha l’arresto in diplotene, si avrà l’ovocita primario. Alla pubertà si
riattiverà la meiosi si formerà l’ovocita secondario e poi il completamento della meiosi II si avrà
quando l’uovo è fecondato.
Nel maschio invece la cellula germinale primordiale, lo spermatogonio va incontro a mitosi durante la
vita fetale e si arresta e la successiva maturazione si avrà nella fase post-puberale.
PROCESSO DI MATURAZIONE DEL FOLLICOLO E DELL’OVOCITA CONTENUTO
Il follicolo primario che ha l’ovocita primario unilamellare, cioè c’è un unico strato delle cellule della
granulosa. Poi abbiamo l’ovocita primario multilamellare, quando le cellule della granulosa proliferano
e si comincia a formare a poco a poco l’antro e poi si forma il cumulo ooforo, il follicolo maturo e
l’ovocita secondario se avviene l’ovulazione.
La maturazione si divide in 3 fasi ed è importante per il controllo.
Dunque la prima fase comprende la formazione del follicolo primordiale, si ha l’ovocita, le cellule
follicolari e lo stroma, poi si passa al follicolo primario unilamellare quando abbiamo l’ovocita e un
unico strato di cellule della granulosa, si comincia a formare anche la zona pellucida che delimita
l’ovocita e comincia a formarsi anche la lamina basale che separa le cellule della granulosa dalle cellule
stromali. Poi abbiamo il follicolo primario multilamellare perchè le cellule della granulosa sono
aumentate di numero, si completa la zona pellucida che delimita l’ovocita dalle cellule della granulosa
e comincia formarsi la teca interna.
Poi si ha il follicolo secondario o vescicolare in cui si comincia a secernere liquido follicolare e si
comincia a formare l’antro. Tutte queste fasi iniziano nella vita fetale e poi cessano. Alla nascita
avremo circa un milione e mezzo, ma nella vita puberale ce ne saranno solo 400000.
La fase del follicolo maturo o di Graff è la seconda fase. Come si chiama l’unico follicolo che
raggiunge la completa maturazione e perché solo uno è il follicolo che matura completamente?
Si chiama follicolo dominante ed è questo che impedirà la maturazione completa di altri follicoli e
quindi altre ovulazioni. Ovviamente si possono avere anche due follicoli maturi o 3 quindi poi si hanno
le gravidanze bigemellari o trigemellari. Di norma quando si hanno le gravidanze multiple dipende da
un fenomeno farmacologico, ad esempio soggetti che sono stati trattati con gonadotropine.
La terza fase corrisponde all’ovulazione e formazione del corpo luteo.
La prima fase è importante perché non dipende da gonadotropine. La maturazione da follicolo
primordiale a primario è un controllo di tipo paracrino, cioè sono fattori ormonali prodotti in situ che
agiscono in situ. Invece le successive fasi dipendono dalle gonadotropine FSH e LH.
Quando il follicolo primario comincia a maturare e si forma il follicolo multilamellare e poi il
secondario dipende dall’ FSH (soprattutto), dall’LH e dagli estrogeni.
L’ovulazione dipenderà soprattutto dall’LH, che agirà sia sulle cellule della granulosa che sulle cellule
tecali. Mentre l’FSH agirà esclusivamente sulle cellule della granulosa.
Infatti quando il follicolo primario comincia a maturare è sufficiente l’FSH, poi l’ulteriore maturazione
dipenderà dalla presenza contemporanea di FSH, LH ed estrogeni. La seconda fase dipende sia
dall’FSH sia dall’LH. L’ovulazione, dipende soprattutto dall’LH, e il mantenimento del corpo luteo
dipende dall’LH.
PRODUZIONE DI ORMONI
Sono di natura steroidea e derivano dal colesterolo che può essere di origine endogena. Se invece
deriva dal mondo esterno, soprattutto dalle LDL,c’ è una proteina , la START, che è capace di
veicolare il colesterolo all’interno della cellula. In quale componente della cellula avverrà l’iniziale
sintesi degli ormoni steroidei? Nei mitocondri. Quindi abbiamo bisogno del passaggio di colesterolo a
livello mitocondriale. La proteina START che lo trasporta all’interno dei mitocondri è fondamentale
per la sintesi degli ormoni steroidei e per la steroidogenesi.
Qualidi questi ormoni ci interessano? L’androstenedione, ormone sessuale maschile, l’estradiolo che di
norma è chiamato E1, l’strone E2, l’estriolo. E poi ovviamente il progesterone.
La sintesi degli ormoni avviene in due fasi: una fase tecale e una fase che si opera a livello delle cellule
della granulosa.
Le cellule tecali, quelle interne in modo particolare, esprimono recettori solo per LH. Infatti l’LH sarà
riconosciuto da un recettore specifico che è correlato all’AMP ciclico che permette la trasformazione
del colesterolo, attraverso diverse vie, in androstenedione che è un ormone sessuale maschile. E’ un
androgeno in poche parole. L’androstenedione è prodotto anche a livello delle cellule dell’ilo.
L’androstenedione, prodotto nella teca interna sotto l’influenza dell’LH, attraverso la via ematica
supera la lamina basale, che separa le cellule della granulosa dalle cellule tecali, e nelle cellule della
granulosa verrà trasformato, grazie alla presenza di un aromatasi, in estrogeno, in modo particolare in
estradiolo. Dunque l’estradiolo verrà prodotto all‘interno delle cellule della granulosa ad opera di
un’aromatasi indotta dall’ LH.
L’LH attiverà l’aromatasi che convertirà l’androstenedione di origine tecale o ilare in estrogeno, che è
l’ormone sessuale femminile, che è importante sia per la maturazione dei follicoli sia per tutti quei
processi che si verificano al livello delle vie genitali per la fertilizzazione. Gli estrogeni infatti
passeranno in circolo o nel fluido follicolare. Nel fluido in particolare svolgerà un ruolo importante per
la maturazione follicolare.
Quindi ricapitolando abbiamo la teoria della duplicità della sintesi degli ormoni estrogeni.
Agli estrogeni in circolo sono dovuti tutti gli effetti generalizzati degli estrogeni, mentre quelli che
passerranno nel liquor follicolare parteciperanno alla trasformazione di follicolo vescicolare in follicolo
dominante.
Ricordate che anche la cellula della granulosa potrà trasformare il colesterolo in pregnenolone e in
progesterone che passerà nella cellula tecale e contribuirà alla produzione di androstenedione. Quando
si forma il corpo il luteo, nelle cellule tecali si produrranno le quantità di progesterone indispensabili
agli effetti luteali del ciclo mestruale.
RUOLO DELL’IPOTALAMO
La produzione degli ormoni follicolo stimolante e luteinizzante è sotto l’influenza dell’ipotalamo.
Questo produce una liberina che è detta gonadoliberinao ormone liberante le gonadotropine. È un
decapeptide, in posizione 2 e 3 della molecola ci stanno l’istidina e il triptofano e questi due residui
costituiscono il centro attivo della molecola. Invece l’acido piroglutammico in posizione 1 e la glicina
in posizione 6 e 10 sono fondamentali per la stabilità della molecola e per il suo legame con il recettore
ipofisario.
Due sono le sedi di produzione: una è l’ipotalamo medio basale e in particolare il nucleo arcuato,
responsabile della secrezione continua, pulsatile della gonadoliberina.
(immagine secrezione pulsatile nel corso del ciclo)
C’ è un picco in fase pre-ovulatoria, poi nella fase lueale ci sarà sempre un andamento pulsatile della
gonadoliberine.
Dunque il nucleo arcuato e in parte quello paraventricolare sono responsabili della secrezione tonica,
continua, pulsatile della gonadoliberina. Invece l’area pre-ottica è responsabile della secrezione ciclica
cioè di quel picco pre-ovulatorio di gonadoliberina.
Vedete (indica un lucido) questo è il profilo della gonadoliberina questo dell’FSH e questo dell’LH.
Tutti i profili delle 3 secrezioni sono pulsatili e hanno tutti un picco pre-ovulatorio. Se vedete
nell’immagine i tre picchi non sono sincroni. Il picco pre-ovulatorio della gonadoliberina precede i
picchi pre-ovulatori di FSH e di LH. Chi ha i profili maggiori tra FSH e LH? Il picco di LH è
notevolmente più alto di quello di FSH.
Si dice normalmente che è questo picco di LH il responsabile dell’ovulazione. Sarà così ma se il
follicolo dominante non è preparato dal picco di FSH allora non si potrà mai avere l’ovulazione.
Dunque abbiamo bisogno di entrambi i picchi. Uno è preparatorio, ed è quello dell’FSH, e quello che
invece scatena l’ovulazione è quello dell’LH. I picchi tra FSH e LH sono perfettamente sincroni,
mentre quello della gonadoliberina precede i due picchi.
Il nucleo arcuato è responsabile della secrezione continua, l’ipotalamo anteriore è invece responsabile
del picco pre-ovulatorio.
Attività fisiologia della gonadoliberina: stimola la liberazione e la sintesi adeno-ipofisaria
dell’ormone follicolo stimolante e dell’ormone luteinizzante.
Vedete che però alcune fibre di questi neuroni che producono la gonadoliberina si porteranno in
strutture extra ipotalamiche, soprattutto al livello del sistema limbico. Queste fibre peptidergiche
portano la gonadoliberina a centri nervosi sia intra che extra ipotalamici e questo è importante ai fini
del comportamento sessuale. La gonadoliberina prodotta da queste fibre peptidergiche fungerà non da
ormone ma da neuromodulatore. Il meccanismo d’azione è legato all’AMP ciclico, ma non
approfondiremo.
CONTROLLO DELLA SECREZIONE DI GONADOLIBERINA
Perché si ha un andamento pulsatile? E perché c’è il picco? Perché i livelli di gonadoliberina sono
maggiori in fase luteale che in fase pre-ovulatoria?
C’è un controllo nervoso. I segnali inibitori e falicitatori che provengono all’ipotalamo attraverso vie
discententi che originano dalla neocorteccia e dal sistema limbico, amigdala e ippocampo,e delle vie
nervose ascendenti che provengono dal midollo spinale e dal tronco dell’encefalo. Questi sono
resposabili delle sollecitazioni psico-sensoriali che possono influenzare la produzione di liberina, per
esempio anticiparla quindi si avrà un‘ovulazione precoce.
Molto importante è anche il ritmo circadiano, buio-luce, attraverso un’altra ghiandola endocrina che
forse mi è sfuggita nell’introduzione iniziale che è l’epifisi. Questa produce una sostanza che è la
melatonina. Di norma la melatonina inibisce la sintesi di liberina e tutto è legato al ritmo circadiano
buio-luce. Infatti vedete la luce colpisce la retina e si inibisce il ganglio cervicale superiore. Se si
inibisce questo ganglio che fa parte del sistema nervoso simpatico si ha un’inibizione di noradrenalina,
quindi un riduzione di produzione di AMP ciclico. Quindi si inibisce l’enzima che produce melatonina.
Se si produce meno melatonina, che ha effetti inibitori per la liberina, allora i livelli di gonadoliberina
tenderenno ad aumentare. Se invece il soggetto è esposto al buio si ha l’efetto opposto. Si attiva il
ganglio cervicale superiore, si attiva l’enzima che orta alla formazione di melatonina e di ocnsguenza
avremo una maggiore sintesi di melatonina. Ciò porterà ad un minore rilascio di gonadoliberina.
Sono interessanti anche i controlli ormonali a feedback positivi e negativi, corti, ultracorti e lunghi.
In fase pre-ovulatoria, il comportamento è diverso se prendiamo la prima metà di questa fase (fino ale 5
giorno) o la seconda metà ( 6- 13 giorno).
È chiaro che all’inizio del ciclo mestruale i livelli di estrogeni e progesterone sono bassi perché stiamo
partendo dalla fine della precedente mestruazione in cui si è verificato una riduzione della produzione
di estrogeni e progesterone.
In questa prima metà del ciclo, modesti livelli di estrogeni e di progesterone mediante un feedback
positivo aumenteranno i livelli di gonadoliberina che a sua volta provocherà un aumento dei livelli di
FSH e di LH. L’aumento di FSH e di LH aumenterà la produzione di estrogeni. Nella seconda parte
della fase pre-ovuatoria abbiamo alti livelli di estrogeni che provocheranno mediante feedback positivo
un ulteriore aumento di gonadoliberina. Questa da un lato aumenta la produzione di LH, ma non di
FSH. Vedremo perché avviene ciò. È chiaro che un aumento di LH provocherà un aumento di estrogeni.
Questo provocherà un picco pre-ovulatorio di gonadoliberina dunque i picchi pre-ovulatori di FSHe LH.
Dunque chi è che detta questo andamento ciclico? Le ovaie con gli estrogeni con feedback positivi e
negativi. Fino a qui sono tutti positivi perché diminuiscono gli estrogeni ed aumentano gli ormoni. In
questa fase aumenta l’LH, aumentano gli estogeni e questo porta i picchi pre-ovuatori di FSH e di LH e
si giunge all’ovulazione.
Nella fase post-ovulatoria ci sarà un aumento di LH che provocherà un aumento di estrogeni e
progesterone. Qui ci sarà un feedback negativo. Infatti l’aumento di estrogeni e progesterone, tipici
della fase post-ovulatoria( soprattutto il progesterone), porterà alla diminuzione della produzione di
liberina,di FSH e di LH, di estrogeni e progesterone e questo porterà al flusso mestruale. A meno che
non ci sia la fecondazione e si produce la gonadotropina corionica, così si mantiene il corpo luteo e i
livelli di estrogeni e progesterone aumentano.
La produzione di gonadoliberina dipende dall’ovaio, mediante i meccanismi a feedback che sono molto
complessi perché io posso averlo positivo o negativo. Se tutto procede in questo modo e il ciclo dura 28
giorni, io avrò una fase pre-ovulatoria in cui si verificano i feedback che porteranno al picco di LH e
quindi allì’ovulazione e poi all’evento post-ovulatorio.
Se tutto dipendesse dall’ovaio e dagli ormoni ovarici i problemi di controllo della gravidanza
fisiologica non ci sarebbero. Però ci sono altri stimoli nervosi di origine psichica, corticale,limbica e
influenze periferiche, psico-sensoriali.
Sull’ipotalamo interferiscono influenze che derivano dal midollo spinale, dal tronco dell’encefalo,
quindi sono psico-sensoriali, ad esempio la vista di qualcosa di gradevole. Oppure la componente
emotiva, come l’aggressività sessuale. Per cui anche le sollecitazioni della corteccia possono
modificare il ritmo ovarico legato agli ormoni estrogeni e progesterone.
RUOLO DELL’IPOFISI
Due gonadotropine ipofisarie:
-FSH o follitropina che stimola il follicolo e soprattutto le cellule della granulosa;
-LH o ormone luteinizzante o luteotropina, che invece permette l’ovulazione e quindi l’attività del
corpo luteo. Per LH ci sono recettori specifici nelle cellule della granulosa, soprattutto nella fase
tardiva della maturazione follicolare e anche a livello delle cellule tecali.
Che ruolo hanno queste gonadotropine?
-Crescita delle gonadi
- La loro maturazione al momento della pubertà
-Il ciclo riproduttivo
-La secrezione di ormoni steroidei
Esiste la gonadotropina corionica che è un ormone fisiologico solo per la donna gravida.
Sono glicoproteine, se vi ricordate infatti sono 3 le glicoproteine prodotte dall’adeno-ipofisi: il TSH e
le due gonadotropine. Sono costituite da due subunità sia le gonatropine che il TSH. Una subunità α
non specifica, cioè è simile per i tre ormoni. Invece la subinità β è specifica. È chiaro che è questa la
componente che determina gli effetti biologici diversi dei 3 ormoni.
FSH e LH sono sintetizzate dalle cellule basofile, cioè le celluleδ.
Invece la gonadotropina corionica è sintetizzata dal sincizio trofoblasto della placenta e anche dal
citotrofoblasto da cui derivano queste cellule.
Come la secrezione di gonadoliberina, questa è una secrezione pulsatile.
Le secrezioni episodiche di FSH e di LH sono asincrone. I picchi secretori di LH sono più alti di quelli
di FSH. Nelle 24 ore ci sono 4 picchi secretori di LH. E’ parte fissa. Non è riconoscibile un ritmo
circadiano.
I livelli ematici di FSH e di LH sono variabili nel corso della vita. I valori sono molto bassi
nell’infanzia, ma è l‘unico periodo di FSH sono maggiori di quelli dell’ LH. Nella pubertà i livelli di
FSH aumenta prima dei livelli ematici di LH, ma alla fine quest’ultimi sono sempre maggiori di quelli
dell’ FSH. Nell’età adulta si fanno evidenti le differenze tra i sessi.
Nella donna le oscillazioni delle gonadotropine diventano ultradiane, dunque mensili, e sono più
evidenti per l’LH.
Vediamo che succede durante il ciclo mestruale vedendo il loro profilo.
Nella prima metà dela fase pre-ovulatoria, quindi i primi 5 giorni, aumentano prima i livelli ematici di
FSH e poi quelli di LH. I livelli di FSH sono più alti di quelli di LH.
Poi invece nella seconda metà della fase pre-ovulatoria i livelli di FSH tendono a diminuire mentre
quelli di LH tendono ad aumentare. Se voi notate queste variazioni dipendono dai livelli di estrogeni
che nella fase pre-ovulatoria tendono progressivamente ad aumentare. Ecco che a metà ciclo abbiamo il
picco di LH. C’è pure il picco di FSH che sarà minore.
Ma chi è che detta il maggiore picco di LH? L’estradiolo che raggiunge i massimi livelli in questa fase
è responsabile dei picchi pre-ovulatori di FSH e di LH.
Durante la fase post-ovulatoria diminuiscono progressivamente i livelli di LH e di FSH.
Nella menopausa aumentano i livelli delle gonadotropine perchè in questa fase le ovaie non funzionano,
non si producono estrogeni e progesteorne e per feedback negativo tendono ad aumentare.
Notate che in fase pre-ovulatoria mentre i livelli di estrogeni tendono ad aumentare, i livelli di
progesterone si mantengono molto bassi. Cominciano ad aumentare quando si ha il picco preovulatorio quindi ad opera dell’LH. Infatti in fase post-ovulatoria gli estrogeni tendono a diminuire e il
progesterone tende ad aumentare. È un feedback negativo.
L’ipotalamo quando rilascia la gonadoliberina, lo fa nel circolo portale ipotalamo-ipofisario. Le
gonadotropine saranno immesse nel circolo generale per raggiungere i tessuti bersaglio.
L’FSH trova i recettori specifici lungo la membrana delle cellule della granulosa, ma non nelle cellule
della teca interna e questo favorisce la crescita e la maturazione dei follicoli ovarici. Permette l'
ipertrofia e la proliferazione delle cellule della granulosa, la differenziazione delle cellule tecali e la
formazione del liquido follicolare. In più stimola la produzione di estrogeni da parte delle cellule della
granulosa.
Invece LH, trova soprattutto i suoi recettori specifici lungo la membrana tecale ma nella fase terminale
della maturazione deifollicoli, soprattutto quando si comincia a forma il follicolo dominante, allora
avremo come conseguenza anche l’azione dell’LH sulle cellule della granulosa. I recettori specifici per
LH si trovano anche nelle cellule dell’ilo dell’ovaio.
Gli effetti fisiologici dell’LH risultano completi e ottimali solo quando concomita l’attività dell’FSH.
Senza un’attività preparatoria dell’FSH non si può avere una completa maturazione del follicolo e
dunque neanche l’ovulazione.
LH stimola le cellule tecali alla sintesi degli androgeni, androstenedione che passerà nelle cellule della
granulosa dove verrà aromatizzato in estrogeni.
Favorisce la sintesi degli androgeni anche nelle cellule stromali e dell’ilo. Provoca lo scoppio del
follicolo ovarico giunto a completa maturazione e provoca dunque l’ovulazione. Favorisce a scoppio
follicolare avvenuto, la formazione del corpo luteo e stimola la produzione di estrogeni, ma soprattutto
di progesterone nelle cellule luteali del corpo luteo.
CONTROLLO DELLA SECREZIONE
Si ha da parte della liberina da parte dell’ipotalamo e mediante feedback diretti e indiretti dall’ovaio o
dall’ipofisi sull’ipotalamo.
Vediamo ora i feedback diretti sull’ipofisi da parte degli ormoni ovarici sul rilascio di gonadotropina.
Anche qui distiguiamo una fase pre-ovulatoria o follicolare e una post-ovulatoria o luteale.
Per quanto riguarda la fase pre-ovulatoria si suddivide la prima metà dalla seconda metà.
Nei primi 5 giorni, che segue la mestruazione precedente, i livelli di estrogeni e progesterone saranno
bassi e a feedback positivi aumenteranno i livelli di FSH e di LH. L’aumento di FSH e di LH
provocherà un ulteriore aumento degli estrogeni. Ci spieghiamo perché nella prima fase aumentano i
livelli di FSHe di LH, anche se quelli dell’FSH sono più alti.
Nella seconda metà della fase pre-ovulatoria si verificava una caduta dell’FSH e un aumento dell’LH.
Nella seconda metà i livelli di FSH diminuiscono. Perché? Se pensiamo che esistano due liberine, una
per l’FSH e una per l’LH, si potrebbe ipotizzare che l’aumento degli estrogeni inibisca a livello
ipotalamico una sola delle due liberine. Ma sappiamo che non è così perché abbiamo una sola liberina
per le due gonadotropine dunque come ci spieghiamo la caduta dell’FSH? È dovuto all’azione delle
cellule della granulosa che producono inibina. Questainibina diminuisce i livelli di FSH perché è
specifica per questo ormone e non agisce sull’LH.
L’aumento degli estrogeni mediante feedback porta all’aumento dell’LH che a sua volta aumenterà i
livelli degli estrogeni. In questa fase infatti abbiamo il più alto livello di estradiolo per un feedback
positivo.
L’aumento degli estrogeni però ad un certo punto provocherà l’aumento dei picchi pre-ovulatori di
FSH e di LH e si avrà l’ovulazione.
Poco prima dell’ovulazione aumentano gli estrogeni e questo comporta sia un aumento dell’FSH sia
dell’LH e l’inibina non viene più prodotta. Viene prodotta un’altra proteina, sempre dalle cellule della
granulosa, che si chiama attivina che aumenta i valori di FSH.
Dunque l’aumento dell’LH è dovuto ad un aumento degli estrogeni per un feedback positivo, ma
l’aumento di FSH è dovuto a tre motivi:
- diminuisce la produzione di inibina;
- aumenta la produzione di estrogeni, per un feedback positivo;
- aumenta la produzione di attivina.
E alla fine avremo i due picchi che produrranno l’ovulazione.
Poi segue la fase post-ovulatoria: dove avviene un aumento dell’LH mentre l’FSH tende a diminuire.
Ciò porterà a un aumento degli estrogeni, soprattutto del progesterone. Per feedback negativo
diminuiscono entrambi e diminuisco estrogeni e progesterone e si ha la regressione del corpo luteo.
Se avviene la fecondazione, si produce la gonodotropina corionica che nonostante i bassi livelli di FSH
e di LH permetterà la formazione del corpo luteo in forma gravidica.
L’inibina è prodotta dalle cellule della granulosa e diminuisce la secrezione di FSH agendo soprattutto
a livello ipofisario e non dell’ipotalamo. La sua sintesi è attivata dagli estrogeni e inibita dal
progesterone. Ciò causa una diminuzione dell’ FSH durante la maturazione follicolare. Il picco preovulatorio di FSH generato dalla gonadoliberina è consentito dal venir bene del blocco dell’inibina.
Il follicolo primario risente esclusivamente dell’ormone follicolo stimolante che porterà alla
proliferazione delle cellule della granulosa che sintetizzeranno estrogeni che in parte passeranno in
circolo e in parte passano nel liquor follicolare, aumentando così i recettori degli estrogeni e dell’FSH
nelle cellule della granulosa e così potenzieranno l’effetto dell’FSH.
Ma le cellule della granulosa produrranno anche fattori di crescita, che permetteranno l’ulteriore
sviluppo del follicolo da primario a secondario, sino al follicolo dominante, l’unico che ovulerà. Infatti
su questo follicolo saranno espressi i recettori per l’FSH nelle cellule della granulosa, e recettori per
l’LH, sia nelle cellule della granulosa sia nelle cellule tecali. Vengono prodotti gli androgeni e quantità
elevante degli estrogeni. L’elevato aumento di estrogeni, per il feedback positivo, provocherà il picco
pre-ovulatorio di FSH e LH e avverrà l’ovulazione.
LH agisce a livello delle cellule della granulosa stimolando la produzione di progesterone che poi
passerà a livello delle cellule tecali in cui si produrrà androstenedione.
Perché soltanto uno matura completamente e gli altri vanno incontro ad atresia? Il follicolo dominante
produce estrogeni e inibina, che ha effetto a feedback sull’adeno-ipofisi, ma agisce anche localmente
con gli estrogeni che sono aumentati e inibiranno la crescita di altri follicoli.
Gli estrogeni sono molto importanti per la maturazione del follicolo perché aumentano i recettori per
l’FSH nelle cellule della granulosa. Gli estrogeni in eccesso assieme all’inibina, inibiranno la crescita
di altri follicoli. Poi ci saranno gli effetti degli estrogeni a livello dell’endometrio che si preparerà
all’accoglimento dell’eventuale cellula uovo fecondata. E poi avremo gli effetti degli estrogeni anche a
livello delle tube, dell’epitelio vaginale e del muco cervicale.
A cosa è dovuto l’ovulazione? Una volta si pensava che quando il follicolo maturo raggiungeva valori
elevati di pressione intrafollicolare si aveva l’ovulazione, per la rottura della parte sottile del follicolo.
Ma non è così perché è stata misurata la pressione intrafollicolare a livello antrale e si è visto che in
realtà nell’emisfero dominante questa pressione raggiunge valori elevati ma da soli non possono
spiegare il processo ovulatorio. Questo infatti avviene anche a causa della proteolisi delle proteine della
parte di follicolo che va in contro a rottura. L’aumento del progesterone porta ad un aumento delle
prostaglandine, principalmente le PGF, che attiveranno gli enzimi proteolitici che attiveranno la lisi di
proteine e quindi l’ovulazione.
Dunque ricapitalando l’ovulazione non è solo legata all’aumento della pressione all’interno dell’antro
ma anche a causa una lisi delle proteine e la parete diviene più fragile. La lisi è dovuta ad enzimi come
la collagenasi e sotto l’influenza delle PGF aumentate a seguito dell’azione del progesterone.
L’inibina tende ad aumentare e poi diminuisce nella fase luteale perché deve contribuire all’inibizione
della produzione di FSH e di LH.
IL COMPORTAMENTO DELLA TEMPERATURA CORPOREA
Durante il picco pre-ovulatorio si ha l’aumento della temperatura corporea della donna. Aumenta di
poco. L’aumento della temperatura corporea è dovuto all’iniziale aumento del progesterone durante la
fase pre-ovulatoria. Durante la fase post-ovulatoria ci sarà un ulteriore aumento della temperatura.
La tecnica di Ogino Knaus cerca di prevedere il momento dell’ovulazione in seguito all’aumento della
temperatura corporea della donna. Questa tecnica è molto importane per valutare l’ovulazione in un
soggetto scarsamente fertile per favorire la fecondazione, soprattuto nei soggetti che hanno un numero
di spermatozoi non adeguati. Così si aumentano le probabilità della fecondazione. La misurazione deve
essere molto precisa, con termometri particolari e deve avvenire né a livello buccale, né
sottoascellare ,nè rettale ma bensì vaginale.
MODIFICAZIONI A LIVELLO DEL MUCO CERVICALE
Parliamo di muco E e muco G.
Il muco E è quello correlato agli estrogeni e precede l’ovulazione.
Il muco G è correlato ad un’ eventuale gestazione, quindi al progesterone.
Il muco E è liquido, filante cioè poco viscoso, e cristallizza sottoforma di foglie di felce. È dovuto agli
estrogeni. Perché ha queste caratteristiche? Perché deve favorire la risalita degli spermatozoi.
Poi si modifica e diviene più denso, meno filante e non cristallizza a forma di felce. Anche qui la
tecnica Ogino Knaus valutava anche la filanza del muco.