Introduzione
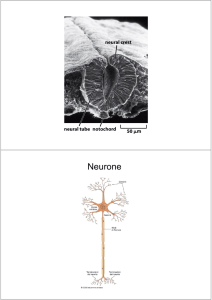
Fig.1 Immagine a immunofluorescenza.
Oligodendrocita
Gli oligodendrociti sono cellule gliali presenti esclusivamente nel sistema nervoso
centrale (SNC), adibiti alla formazione della guaina mielinica attorno agli assoni dei
neuroni. In tal modo garantiscono una maggiore velocità nella conduzione (saltatoria)
dell’ impulso.A differenza di quanto accade nel sistema nervoso periferico (dove tale
processo è effettuato dalle cellule di Schwann) un singolo oligodendrocita è in grado di
mielinizzare più assoni adiacenti per via dei numerosi prolungamenti che lo
caratterizzano (come mostrato in fig.1). Il processo che porta alla migrazione, alla
differenziazione degli oligodendrociti e dunque alla successiva mielinizzazione si basa su
modificazioni citoscheletriche continue, mediate da una serie di segnali intracellulari ed
extracelluari.
Discussione
La componente citoscheletrica degli oligodendrociti è caratterizzata da microfilamenti(MF) e microtubuli(MT), mentre sono
totalmente assenti i filamenti intermedi. Recentemente si è scoperto che tali cellule esprimono una particolare forma di
tubulina, la IV-tubulina, fino ad ora trovata espressa nel SNC solo in queste cellule. Seppur le specifiche funzioni della IVtubulina devono essere ancora identificate, il fatto che solo gli oligodendrociti esprimano tale
proteina sottolinea sicuramente l’ importanza che riveste il citoscheletro nell’ ambito della
mielinizzazione. Infatti l’ espressione di tale tubulina in altre popolazioni cellulari è
associata ad una maggiore stabilità dei MT. Nelle precoci fasi della mielinizzazione l’
oligodendrocita forma una serie di protusioni, i filopodi, dovuti a rapide polimerizzazioni di
MF nella zona sottostante la membrana. Il complesso formato da N-WASP e Arp2/3
determina ramificazioni dei MF e un conseguente allargamento dei filopodi, i quali tramite
la polimerizzazione dei MT che vi penetrano e l’ attivazione di altri complessi molecolari
come WAVE, diventano lamellipodi. L’ intero sistema, ovviamente, è regolato da segnali
extracellulari; fattori di crescita quali PDGF, bFGF, HBF e GGF, rilasciati da neuroni e
astrociti, sembrano essere coinvolti nei vari processi a cui va incontro l’ oligodendrocita sin
quando non stabilisce il contatto con l’ assone. Fondamentale risulta essere anche il ruolo
giocato dalla matrice extracellulare(ECM). Molecole quali fibronectina, laminina, collagene
(di cui la ECM è ricca) vanno a interagire con specifiche proteine di membrana. In
particolare il legame della laminina all’ 6 1 integrina determina una fosforilazione della
catena 1 di quest’ ultima ad opera della chinasi ILK del complesso IPP. È questo il complesso
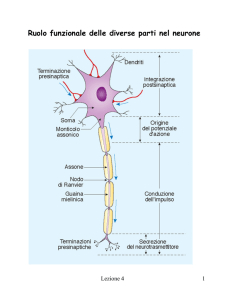
Fig.2 In primo piano: guaina
mielinica
che in altre popolazioni cellulari si trova legato intracellularmente all’ integrina. La
di rivestimento di un
fosforilazione
della catena 1 comporta il reclutamento di N-WASP e Arp 2/3, e dunque una rapida polimerizzazione della
assone.
actina. Tale meccanismo pare intervenire nelle prime fasi della differenziazione degli oligodendrociti. Durante la
formazione della guaina mielinica sembra invece avere un ruolo importante un altro recettore della laminina recentemente
individuato: il distroglicano. Ciò a testimonianza della necessità di diversi recettori durante diverse fasi e dunque della
complessità del fenomeno. Ma non dobbiamo dimenticare che la differenziazione comporta modificazioni a livello della
membrana plasmatica, che includono la formazione di zattere lipidiche, costituite essenzialmente da colesterolo,
glicosfingolipidi e fosfolipidi con catene degli acidi grassi saturi. Praticamente subito, a livello di tali zattere sono reclutati
recettori PDGF, a testimonianza dell’ importanza di tale fattore nell’ ambito del precoce differenziamento. Il contatto dell’
oligodendrocita con l’ assone determina modificazioni a livello delle zattere lipidiche, che risultano arricchite in tal
maniera soprattutto di 6 1 integrina e Fyn chinasi. Quest’ ultima in particolar modo è responsabile del legame con i MT,
nonché con tau, una MAP che stabilizza questi ultimi, fondamentale per il conseguente trasporto di vescicole contenenti
componenti mielinici al foglio mielinico in progressiva crescita e in fase di spiralizzazione attorno all’ assone. L’ importanza
di tale chinasi è testimoniata dal fatto che topi mancanti della proteina mostrano seri difetti nella mielinizzazione. Gli
eventi molecolari alla base della successiva e finale fase di compattazione della guaina mielinica con conseguente estrusione
del citoplasma
non sono ancora stati ben decifrati. Si suppone tuttavia siano determinati da disassemblaggio e
riorganizzazione del citoscheletro e in ciò può essere importante la statmina, proteina che destabilizza i MT.
Conclusione
È sicuramente evidente la complessità dei meccanismi che sono alla base della mielinizzazione, nonché l’ indispensabile
presenza del citoscheletro. Tuttavia ancora molti studi sono necessari per giungere ad una globale e dettagliata visione. Da
notare è l’ importanza di una precisa e fine regolazione di tali meccanismi. Si pensi solo ad una sovrespressione della
statmina nel momento in cui è richiesta una grande stabilità microtubulare per la formazione della guaina mielinica. Il
risultato sarebbe l’ incapacità di garantire un rivestimento all’ assone. Ed infatti in pazienti affetti da sclerosi multipla è
stata rilevata una sovrespressione di tale proteina.
References
Role of the oligodendroglial cytoskeleton in differentiation and myelination. Bauer NG, Richter-Landsberg C, Ffrench-Constant C.
Cristina Pizza