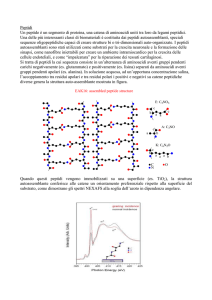
PROGRAMMA DI RICERCA
TITOLO: “ NUOVI FATTORI CARDIO-ATTIVI : I NEUROPEPTIDI ENDOGENI AD AZIONE ANTIADRENERGICA ”
PROPONENTE: DR. TOMMASO ANGELONE
BASE DI PARTENZA
SCIENTIFICA
Il cervello e il sistema cardiovascolare sono coinvolti funzionalmente in importanti interazioni
incentrate sul mantenimento dell'omeostasi. Due sistemi, conosciuti da oltre un secolo, collegano il
cervello e il sistema cardiovascolare: il sistema nervoso autonomo (SNA) attraverso dirette azioni
neurali, e il sistema neuroendocrino umorale attraverso l'asse ipotalamo-ipofisi-surrene (asse HPA).
Le conoscenze su queste interazioni sia in condizioni normali che, soprattutto, in condizioni di
stress, si sono evolute velocemente lungo memorabili tappe. Nel 1930 il ruolo emergente del
sistema nervoso simpatico (SNS) nella risposta allo stress fu ampiamente chiarito da Walter Cannon
and Hans Selye. In particolare, Cannon studiò la risposta allo stress, da lui indicata come reazione
di "lotta o fuga", e riconobbe la "saggezza del corpo" nella risposta "generalizzata" del sistema
simpatico che agisce durante lo stress, mentre allo stesso tempo mise in risalto le più "discrete"
influenze esercitate dal sistema nervoso parasimpatico (SNP) (Chrousos and Gold, 1992).
Successivamente, il concetto di "sindrome di adattamento" o di "risposta allo stress", sviluppato da
Selye (1936), svelò il ruolo cruciale del SNS e dell'asse HPA nel mantenimento dell'omeostasi in
risposta allo stress contrassegnata dall'aumento dei livelli periferici di catecolammine (CAs) e di
glucocorticoidi.
Da allora il cuore e il sistema vascolare sono diventati un paradigma classico di organi minacciati
dallo stress. In particolare, il cuore è ora visto come interfaccia di integrazione tra le terminazioni
nervose noradrenergiche, che rilasciano il loro principale neurotrasmettitore norepinefrina (NE), e
le CAs circolanti, l’adrenalina (E) e, in minor misura, NE, secrete dalla midollare del surrene.
E' da tempo risaputo che elevati livelli di CAs inducono necrosi cardiaca (Raab, 1963; Benjamin et
al., 1989; Teerlink et al., 1994; Tan et al., 2003; Goldspink et al., 2004). E' anche noto che la
risposta iniziale del cuore ad uno stress eccessivo e prolungato è un aumento di volume della massa
cardiaca fino allo stato di ipertrofia cardiaca, cioè un adattamento compensatorio che previene il
progressivo deterioramento della funzione di pompa (Grossman et al., 1975; Chein, 1991). Tuttavia,
come abbiamo appreso negli ultimi decenni, spesso lo stress ha il sopravvento sul compenso
omeostatico, rendendo insufficiente il cuore ipertrofico. Allo stesso tempo, vi è una elevata e
sostenuta attivazione del SNS e del RAS, nel tentativo di controllare sia la gittata cardiaca che la
pressione sanguigna sistemica (Esler et al., 1997). Dati recenti a livello cardiaco indicano che nei
processi di deterioramento, queste cascate eccitatorie, se lasciate incontrollate, potrebbero essere più
deleterie dello stesso stress. E' importante notare che, nello scompenso cardiaco umano, la sostenuta
attivazione cronica del SNS e l'associato aumento del segnale adrenergico attraverso le CAs,
assume un significato prognostico avverso, spesso accelerando i processi patologici (Cohn et al.,
1984). Chiaramente, il risultato degli studi in questo ambito della ricerca cardiovascolare è stato di
grande rilevanza clinica, fornendo le basi per una terapia farmacologica anti-adrenergica, che
include i beta-bloccanti, ancora oggi tra i farmaci più usati nel mondo.
Il nostro progetto suggerisce come obiettivo futuro la ricerca di alcuni meccanismi neuroendocrini
di base capaci di interagire con l'iperattivazione del sistema adrenergico e che potrebbero avere una
forte influenza sull'inizio e il decorso delle condizioni fisiopatologiche.
I granuli secretori delle cellule cromaffini, cioè le cellule derivate dalla cresta neurale che
costituiscono la midollare del surrene e il sistema neuroendocrino diffuso, contengono CAs e una
grande varietà di proteine e peptidi (Goumon et al., 1998).
Questi ultimi includono alcuni neuropeptidi (per esempio il neuropeptide Y e il peptide
vasointestinale) e alcuni pro-ormoni. In seguito ad una adeguata stimolazione, questi peptidi
insieme con le CAs sono rilasciati nel circolo sanguigno dalla ghiandola surrenale o a livello
d'organo da cellule neuroendocrine, neuronali ed endocrine locali, partecipando quindi a molti e
importanti processi omeostatici. Nell'ultimo decennio maggiore enfasi è stata data alla
caratterizzazione biochimica e funzionale dei principali costituenti dei granuli cromaffini, cioè
quelle proteine idrosolubili ad alto peso molecolare costituite dalla famiglia delle Cromogranine e
dalla PEBP (phosphatidylethanolamine-binding protein),.
La famiglia delle Cromogranine comprende la Cromogranina A (CGA) e B (CGB) (Aunis e MetzBoutigue, 2000). La CGA, proteina ubiquitaria, distribuita in molti tessuti di mammifero, incluso il
cuore (Steiner et al., 1990), è rilasciata mediante esocitosi e rapidamente processata da una proormone convertasi e da altre proteasi in maniera tessuto-specifica (Metz-Boutigue et al., 1993),
dando origine a diversi peptidi. I peptidi CGA-derivati, cioè catestatina, pancreastatina, parastatina,
vasostatine 1 e 2, sono stati proposti come nuovi ormoni regolatori coinvolti nel rilascio di CAs
dalla midollare del surrene, nel bilancio plasmatico di glucosio e calcio, nella immunità innata e
nella motilità vascolare (Helle et al., 2001; Helle 2004). In particolare, la pancreastatina e la
catestatina sono stati ipotizzati come componenti del sistema omeostatico "zero steady-state error",
secondo cui, due fattori regolatori agiscono in contrapposizione con risultato integrato (Koeslag et
al., 1999; Koeslag e Saunders, 2004). Pertanto, in seguito ad una perturbazione, tali componenti
riportano la variabile al valore di controllo (Koeslag et al., 1997; Saunders et al., 1998). La
Vasostatina-1 (VS-1) e la vasostatina-2 (VS-2) corrispondono rispettivamente ai polipeptidi CGA
1-76 e CGA 1-113 derivati dal clivaggio del primo e secondo paio di a.a. basici del dominio Nterminale della CGA. Essi sono stati chiamati vasostatine (VSs) per la loro capacità di regolare la
pressione sanguigna sia a livello arterioso che venoso, riducendo la vasocostrizione indotta da
concentrazioni elevate di potassio e da agonisti come noradrenalina ed endotelina (Aardal e Helle,
1992; Angeletti et al., 1994). Di conseguenza, è stato suggerito per essi un ruolo protettivo sul
sistema vascolare rispetto a intensi stimoli eccitatori, come nella risposta da stress (Helle e
Angeletti, 1998). La scoperta che elevati livelli di CGA in pazienti con insufficienza cardiaca sono
un indice di mortalità e gravità della malattia, sottolinea l'importanza delle VSs nell'omeostasi
cardio-circolatoria (Ceconi et al., 2002). Più recentemente, in assenza di dati circa le loro azioni sul
cuore, alcuni di noi hanno studiato gli effetti della VS-1 e VS-2 esogene sulla performance cardiaca
in vitro nella rana, nell'anguilla e nel ratto, paradigmi di tipici cuori di vertebrati.
Tali ricerche hanno dimostrato che anche il cuore può essere importante bersaglio per le VSs.
Infatti, entrambi i peptidi deprimono la contrazione miocardica (inotropismo negativo) in condizioni
basali, e antagonizzano l'incremento della contrattilità (inotropismo positivo) indotto dalla
stimolazione adrenergica (Corti et al., 2002; Imbrogno et al., 2004; Tota et al., 2004). Inoltre,
studiando il rapporto struttura-funzione, è stato visto che l'inotropismo beta-adrenergico è
antagonizzato dal frammento STA-CGA 1-78, dal frammento CGA7-57 e dal frammento Nterminale CGA1-40 con ponte disolfuro intatto (Tota et al., 2003). Nell'insieme, questi risultati per
la prima volta indicano le VSs come possibili modulatori cardioinibitori, capaci di proteggere il
cuore da intense stimolazioni, come nella risposta da stress (Tota et al., 2004).
Recentemente, usando tecniche di proteomica, è stato dimostrato che la PEBP
(phosphatidylethanolamine-binding protein), l'altra proteina idrosolubile processata nella matrice
intragranulare dei granuli cromaffini, è anche precursore di un nuovo prodotto di maturazione, lo
HCNP (Hippocampal Cholinergic Neurostimulatory Peptide) (Goumon et al., 2000; Kieffer et al.,
2003). Il PEBP, proteina di 21-kDa, è ubiquitaria, essendo stata identificata in molti organismi quali
batteri, lieviti, piante, nematodi, drosophila e mammiferi, (uomo, ratto, topo e bue) (Shoentgen e
Jollès, 1995), così come in molti tessuti (fegato, rene, testicoli, cervello, ghiandola surrenale di ratto
e piastrine umane) (Frayne et al., 1999; Ojika et al., 2000). Il PEBP è stato anche chiamato RKIP
(Raf-1Kinase Inhibitor Protein) (Yeung et al., 1999). Membri di questa famiglia di proteine sono
coinvolti in diversi sistemi-segnale come dimostrato per il PEBP di mammifero, che modula
l'azione delle proteine G eterometriche e che inibisce proteasi seriniche come la MAP kinasi e la
NF-kB (Vallée et al., 2003). Particolare attenzione è stata rivolta alla porzione N-terminale del
PEBP di mammifero, un frammento di undici a.a. denominato HCNP (Hippocampal Cholinergic
Neurostimulatory Peptide). Lo HCNP, così chiamato poiché è stato purificato dall'ippocampo di
ratto, aumenta la sintesi di acetilcolina (ACh) in nuclei del setto mediale in coltura (Ojika e Appel,
1984; Ojika et al., 2000). Nei neuroni colinergici, lo HCNP produce un aumento dose- e tempodipendente dell'attività e della Vmax dell'enzima acetilcolintransferasi (ChoATase), influenzando lo
sviluppo di fenotipi neuronali colinergici (Ojika et al., 1994). Relazioni tra caratteristiche strutturali
e fisicochimiche dello HCNP e del suo precursore hanno evidenziato una importante e duplice
funzione di questo peptide (Vallée et al., 2003). In primo luogo, essendo lo HCNP la regione Nterminale del PEBP, dotata di lipofilia, esso interagisce con le membrane cariche negativamente e,
al tempo stesso, attraverso la sua capacità di formare legami idrogeno, partecipa alla conformazione
e alla stabilità del sito attivo del PEBP. In secondo luogo, lo HCNP, libero in soluzione, appare ben
strutturato, almeno per quanto riguarda la regione centrale (D3LSKW7), nella quale la
conformazione è simile a quella osservata nel PEBP cristallizzato (Vallée et al., 2003). Pertanto, lo
HCNP libero può agire come segnale in cellule e tessuti bersaglio contribuendo in tal modo ad
alcune delle funzioni biologiche del PEBP (Vallée et al., 2003). A parte la loro presenza nel tessuto
nervoso, sia il PEBP (per esempio, nel ratto adulto: Frayne et al., 1999) che lo HCNP (per esempio
nel ratto giovane: Katada et al., 1996) sono stati immunolocalizzati in altri organi e tessuti, quali la
parete dei vasi sanguigni, la ghiandola surrenale, l'intestino e il rene. Recentemente, usando la
midollare del surrene bovino, Goumon e collaboratori (2004) hanno dimostrato che il PEBP e lo
HCNP sono presenti anche nelle cellule cromaffini (matrice intragranulare) da cui vengono secreti
nel circolo sanguigno insieme alle CAs. Inoltre, essi sono stati ritrovati anche nelle piastrine e nel
siero (Goumon et al., 2004), nel liquido cerebrospinale dei pazienti affetti da Alzheimer (Tsugu et
al., 1998) e negli adipociti di topo (Kratchmarova et al., 2002). L'ampia distribuzione dello HCNP e
del PEBP a livello d'organo e tissutale, unitamente alla loro co-localizzazione nei granuli secretori
delle cellule cromaffini con altri neuropeptidi e cromogranine (Kieffer et al., 2003), suggeriscono
che entrambi potrebbero rappresentare nuovi fattori endocrino/paracrini.
Usando come modello un preparato di cuore isolato e perfuso di rana, alcuni di noi hanno
dimostrato per la prima volta che lo HCNP esercita in condizioni basali inotropismo negativo da 1011
a 10-7 M e si oppone all'inotropismo positivo dell'isoproterenolo (ISO), classico agonista
adrenergico (Goumon et al., 2004). E' di rilievo che questi risultati sono stati confermati anche sul
cuore di ratto isolato e perfuso alla Langendorff (Goumon et al., 2004; Angelone et al., 2005). Ciò
suggerisce che lo HCNP, al pari delle VSs, possa essere un nuovo potenziale modulatore cardioinibitore.
DESCRIZIONE DEL PROGRAMMA DI RICERCA
La recente scoperta del ruolo cardioinibitorio ed antiadrenergico delle VSs e dello HCNP
suggerisce il loro possibile coinvolgimento nella modulazione dei segnali adrenergici. Le seguenti
fasi di lavoro saranno effettuate per confermare la nostra ipotesi:
a) Interazione dei peptidi: sinergismo, sommazione, potenziamento o antagonismo;
b) possibile ruolo cardioprotettivo dei peptidi;
c) VSs e HCNP: effetti su modelli di cuore di ratto iperteso.
a)INTERAZIONE DEI PEPTIDI: SINERGISMO, SOMMAZIONE, POTENZIAMENTO O ANTAGONISMO
Considerato che le VSs e lo HCNP sono co-rilasciati in seguito a stimolazione adrenergica, il nostro
obiettivo è di evidenziare una loro possibile interazione qualitativa e quantitativa. Le interazioni
farmacodinamiche sono utili in terapia e rivestono una notevole importanza in quanto hanno la
finalità di ottenere effetti combinati benefici. Quindi, intendiamo verificare se i due peptidi possano
esercitare simili azioni con differenti meccanismi, e come queste azioni si integrano in un effetto
finale a livello cardiaco.
Analizzando su preparati cardiaci, sia in condizioni basali che sotto stimolazione adrenergica, i
profili cardiotropici di ciascun peptide con quelli ottenuti dalla contemporanea somministrazione
dei peptidi, sarà possibile ottenere informazioni sulle possibili interazioni tra VSs e HCNP quali ad
esempio sinergismo, sommazione, ecc. L'interazione tra VSs e HCNP sarà valutata: 1) mediante
curve dose-effetto delle VSs più HCNP; 2) mediante curve di competizione di ciascun peptide in
presenza di dosi fisse dell'altro peptide. Da queste curve saranno calcolati i rispettivi valori di EC50
che daranno chiare indicazioni dell'interazione farmaco-dinamica che i due peptidi esercitano a
livello cardiaco.
Non essendo presente in letteratura alcune evidenza sperimentale che dimostri la presenza di
recettori specifici per i due peptidi, sarà inoltre nostro obiettivo analizzare funzionalmente il
meccanismo d’azione con cui i due peptidi esercitano la loro azione cardiotropa. I nostri studi
condotti finora riguardo le VSs hanno dimostrato un'azione inotropa negativa sul cuore di ratto
secondo Langendorff in un range di concentrazioni nanomolari (Cerra et al., 2005). Poiché al
momento non esistono dati che evidenziano l'esistenza di recettori specifici per le VSs (Helle et al.,
2001), le nostre ricerche sono attualmente indirizzate ad esplorare se tale azione cardiotropica
coinvolge meccanismi mediati da recettori o interazioni con proteine inter- e/o intracellulari. Nel
cuore di ratto, l'inotropismo negativo della VS1 è abolito da antagonisti dei recettori adrenergici, ma
non di quelli colinergici (Angelone et al., 2004). Più che ad una azione diretta del peptide sul sito
attivo del recettore adrenergico, ciò fa supporre interazioni secondarie tra VSs e proteine regolatrici
agendo, per esempio, su siti allosterici dei recettori adrenergici. E' riportato che le VSs potrebbero
attivare pathways di trasduzione dei segnali inter- e/o intracellulari attraverso la perturbazione della
membrana cellulare causata dall'interazione della porzione lipofilica della VS1 con domini presenti
sul bilayer lipidici (caveole, recettori scavenger, etc.) (Maget-Dana et al., 2002). Inoltre, studi
sull'attività pro-adesiva delle VSs, in particolare della VS1, hanno suggerito un loro possibile
coinvolgimento nel modulare i meccanismi di adesione nello spazio extracellulare, per esempio
regolando la capacità o l'affinità delle integrine per i loro ligandi (Gasparri et al., 1997). Nei
miocardiociti la modulazione dei canali ionici mediante l'interazione del citoscheletro con i recettori
adrenergici potrebbe influenzare la performance contrattile (Malan et al., 2003). In questo contesto,
appare interessante esplorare le interazioni fra le VSs e le ERM (Ezrin-Radixin-Moesin), proteine
regolatrici dell'interazione membrana plasmatica-citoscheletro (Bretscher et al., 2000), e, allo stesso
tempo, tra le VSs e le fosfoproteine coinvolte nella comunicazione intercellulare, quali le
connessine (Sàez et al., 2003).
Nulla è riportato, invece, riguardo il meccanismo d'azione dello HCNP. La struttura cristallografica
del PEBP umano e bovino ha rivelato che lo HCNP potrebbe essere coinvolto in specifiche
interazioni con le membrane cellulari. In particolare, lo HCNP, studiato in soluzione attraverso
dicroismo circolare e spettrometria NMR, appare ben strutturato (Vallée et al., 2003). La
conformazione ad "uncino" dello HCNP, conferita dalla presenza di una serie di giri nella sua
struttura molecolare, gli permette di interagire con le cariche negative delle membrane (Vallée et
al., 2003). Perciò, è plausibile investigare se questo peptide, come le VSs, può attivare risposte
cellulari attraverso interazioni con specifici domini lipidici delle membrane cellulari.
Studi condotti sul cuore di rana hanno mostrato che lo HCNP induce effetti inotropi negativi già a
basse concentrazioni (10-12 M) ed è in grado di contrastare la stimolazione adrenergica (Goumon et
al., 2004). Anche nel cuore di ratto dati preliminari indicano un effetto inotropo negativo dosedipendente, rilevabile dalla significativa riduzione della pressione ventricolare sinistra (LVP) e
dello RPP (Rate Pressure Product), ma nessuna influenza è stata osservata sulla frequenza cardiaca
(HR) e sulla pressione coronarica (CP). Inoltre, lo HCNP è in grado di contrastare il cronotropismo
e l'inotropismo positivo e la vasodilazione coronarica mediati dalla stimolazione adrenergica
(Angelone et al., 2005). Al fine di corroborare i risultati preliminari sul ruolo cardiomodulatore e
antiadrenergico dello HCNP, la sua natura antagonistica sarà valutata mediante studi di
competizione fra la stimolazione adrenergica e quella indotta dallo HCNP da cui si ricavano i
relativi valori di EC50. L'identificazione delle VSs e dello HCNP come fattori cardioinibitori, cioè
stabilizzatori del tono adrenergico nell'omeostasi cardiocircolatoria, potrebbe sfidare nuovi studi
sulle interazioni mediate dai peptidi tra sistema adrenergico e cuore. Inoltre, l'uso di inibitori delle
connessine, delle integrine o del citoscheletro permetterà di rilevare possibili dirette interazioni
intracellulari dei due peptidi.
b) POSSIBILE RUOLO CARDIOPROTETTIVO DEI PEPTIDI
In collaborazione con il laboratorio di Fisiologia Cardiovascolare del Dipartimento di Neuroscienze
dell’Università di Torino verrà studiato il ruolo della VS1 e dello HCNP come agenti
cardioprotettivi in condizioni di precondizionamento ischemico.
Con il termine di precondizionamento ischemico si indica la protezione miocardica che
sperimentalmente può essere prodotta dalla transitoria ipossia ottenuta con uno o più brevi episodi
di occlusione di un grosso ramo coronarico. In generale la protezione da precondizionamento può
essere ottenuta con tecniche che limitano la disponibilità di ossigeno per il cuore (Shizikuda et al.,
1992). La protezione indotta dal precondizionamento ischemico consiste generalmente in una
limitazione delle lesioni causate da un successivo episodio di ischemia e di riperfusione, nel quale la
durata dell’ischemia sia sufficientemente lunga (30 min) da procurare infarto (Lawson e Downey,
1990). A causa di questa protezione si ha una limitazione dell’area di infarto, una minore durata
dell’ipocontrattilità post-ischemica, nota anche come stunning miocardico e una minore incidenza
delle aritmie soprattutto nel periodo di riperfusione (Bolli et al., 1997; Parratt e Vegh, 1994). La
minore incidenza delle aritmie comporta una protezione contro l’evenienza letale dovuta alla
fibrillazione ventricolare. Vari sono i meccanismi con i quali si ritiene possa attuarsi la protezione
miocardica da precondizionamento. Un ruolo determinante sembrerebbe essere dovuto ad
un’apertura di canali mitocondriali per il potassio ATP- dipendenti (Sasaki et al., 2000). Il
precondizionamento esercita anche un effetto a carico dei vasi coronarici. Nel corso di un
successivo severo episodio di ischemia-riperfusione (I/R), l’endotelio di questi vasi non vede ridotta
la sua capacità di liberare NO il quale media la protezione vascolare esercitata dal
precondizionamento ischemico (Gattullo et al., 1999;Pagliaro et al., 2001). Alla luce dei dati
presenti in letteratura che pongono la vasostatina come agente cardiotropico, il presente lavoro di
ricerca si pone l’obiettivo di indagare il ruolo della VS1 in presenza di precondizionamento
ischemico studiando perciò il peptide come probabile agente cardioprotettivo in grado di limitare i
danni derivanti da una I/R. In questo modo verrebbero esplorati i meccanismi biochimici e
farmacologici che sottendono l’eventuale effetto cardioprotettivo della VS1 e dello HCNP al fine di
chiarirne il potenziale terapeutico.
c) VSs E HCNP: EFFETTI SU MODELLI DI CUORE DI RATTO IPERTESO
Nella ricerca cardiovascolare, l'impiego di modelli animali si è rivelato particolarmente utile nello
studio delle malattie cardiovascolari nei primi stadi di sviluppo, dei meccanismi che sottendono le
condizioni patologiche, degli effetti e dei meccanismi di diversi farmaci. Per l'ipertensione, il
modello animale più comunemente usato è il ratto SHR (spontaneously hypertensive rat) mentre il
ratto Wistar Kyoto (WKY) rappresenta il controllo normoteso (Kurtz e Morris, 1987). Il maschio
SHR è anche utilizzato come modello per studiare i cambiamenti nei meccanismi di segnali indotti
dall'ipertensione umana (Takata e Kato, 1996). La perdita di variazioni inter-individuali è uno dei
maggiori vantaggi dell'uso degli SHRs (Lindpaintner et al., 1992). Un altro vantaggio legato all'uso
dello SHR è che le fasi ipertensive quali pre-ipertensione, sviluppo e ipertensione sostenuta sono
ben standardizzate, e ciascuna fase dura almeno per diverse settimane (Folkow, 1993). Usando lo
SHR come modello ben caratterizzato di una sostenuta attivazione del SNS e del segnale
adrenergico, intendiamo valutare se cambiamenti ipertensione-dipendenti nell'automatismo
cardiaco, nella meccanica cardiaca e nella funzione coronarica, possono essere ridotti o annullati dal
trattamento con vasostatina e/o con HCNP. Utilizzando cuori di ratto secondo la metodica
Langendorff, saranno condotti esperimenti paralleli su ratti normotesi e ipertesi. In un primo
momento saranno determinati gli effetti sui parametri cardiaci dei peptidi somministrati
singolarmente. Successivamente, i ratti ipertrofici saranno trattati somministrando
contemporaneamente i due peptidi allo scopo di valutare la possibilità di effetti sinergici e/o
additivi. L'analisi immunoistochimica permetterà di valutare l'eventuale over-espressione dei
peptidi nei cuori ipertesi. Si attendono dai risultati ottenuti sui modelli SHR importanti informazioni
preliminari riguardo il coinvolgimento delle VSs e dello HCNP negli stadi patologici, suscettibili di
aprire nuove strade per la progettazione di farmaci basati sulle strutture dei peptidi e per nuovi
approcci farmacologi.
MATERIALI E METODI
i) Preparati cardiaci di ratto secondo Langendorff (ratti normotesi e ipertesi al primo e terzo stadio della malattia).
Questa tecnica è estesamente utilizzata per lo studio della fisiologia e della fisio-patologia cardiaca (vedi per referenze
Sutherland e Hearse, 2000). Il cuore di ratto verrà incannulato a livello dell'aorta per iniziare la perfusione retrograda a
flusso costante (10-13,5 ml min-1g-1 di cuore fresco) e quindi collegato all'apparato di perfusione. Un trasduttore di
pressione collegato alla cannula aortica consentirà di misurare la pressione coronarica. All'interno del ventricolo sinistro
sarà inserita una sonda di pressione collegata ad un trasduttore di pressione. Entrambi i trasduttori saranno collegati con
il sistema di acquisizione McLab. Tale metodica consentirà di seguire nel tempo l'andamento dei parametri relativi
all'attività sistolica (pressione ventricolare sinistra, +(LVdP/dt)max, tempo impiegato per raggiungere il picco di
tensione, frequenza, lavoro), al recupero diastolico (pressione telediastolica sinistra, -(LVdP/dt)max, tempo per ottenere
il 50% del rilassamento) ed alla pressione coronarica. Dopo aver stabilizzato il preparato, questo sarà perfuso con il
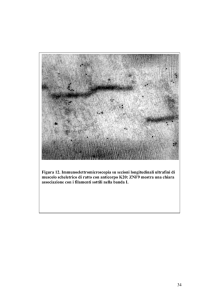
mezzo di perfusione arricchito della sostanza in esame. ii) Immunoistochimica. Per l'immunodeterminazione delle VSs e
dello PEBP/HCNP i cuori saranno rimossi, lavati in PBS, crioprotetti, fissati in azoto liquido e conservati a -80°C.
Sezioni cardiache saranno ottenute al criostato (Microm HM505E), postfissate con acetone e conservate a -20°C.
L'immunolocalizzazione sarà effettuata usando anticorpi primari monoclonali anti-VSs o anticorpi policlonali anti
PEBP/HCNP. Anticorpi secondari commerciali coniugati TRITC e FITC saranno usati per identificare l'antigene. Il
peptide PEBP/HCNP, gli anticorpi monoclonali e policlonali anti-PEBP/HCNP saranno prodotti e forniti dalla Prof.
Metz-Boutigue (INSERM di Strasburgo, Francia). Tale collaborazione è documentata da pubblicazioni (Tota et al.,
2003; Goumon et al., 2004), da un progetto VINCI di dottorato in co-tutela Italia-Francia e dal programma GALILEO
nell'ambito delle azioni integrate Italia-Francia. Le VSs e gli anticorpi monoclonali primari anti-VSs saranno fornite dal
Dr. Corti (Istituto San Raffaele, Milano). Tale collaborazione è documentata da recenti pubblicazioni (Corti et al., 2002;
Corti et al., 2004).
RISULTATI ATTESI
Sia le VSs che lo HCNP, cioè i rispettivi prodotti della CGA e del PEBP presenti nelle cellule
cromaffini e co-rilasciati con le CAs, sembrano agire come modulatori negativi e sono capaci di
opporsi agli stimoli adrenergici. Tenendo conto di tali effetti, il presente progetto di ricerca ha lo
scopo di verificare l'ipotesi che le VSs e lo HCNP possano agire in modo integrato per regolare
finemente a livello agire come controregolatori con i quali, a diversi livelli, l'organismo viene
protetto dalla iperstimolazione. Le azioni "anti-adrenergiche" delle VSs e dello HCNP, di recente
scoperta, potrebbero essere rilevanti nella risposta allo stress, allorché alcuni organi come il cuore
diventano bersaglio preferenziale delle CAs. L'uso di sequenze primarie delle VSs e dello HCNP
nella ricerca fisio-farmacologica potrebbe avere una forte ricaduta a livello clinico.
RIFERIMENTI BIBLIOGRAFICI
•
•
Aardal S. and Helle K. B., (1992). Reg. Pept. 41: 9-18.
Angeletti R. H., et al., (1994). Acta Physiol. Scand. 152: 11-19.
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
•
Angelone T., et al. Role of Vasostatin-1 on Cytoskeletal modulation in rat cardiac myocytes. XI Congresso
Nazionale della SocietàItaliana di Ricerche Cardiovascolari (SIRC), Latina, 23-25 settembre, 2004.
Angelone T., et al. The Hippocampal Cholinergic Neurostimulating Peptide (HCNP): a novel cardioregulatory
molecule? Data will be presented in SEB Annual Main Meeting, Barcellona 11-15 July, 2005.
Angelone T., et al., Cardiosuppressive action of vasostatins on the Langendorff isolated and perfused rat heart.
Proceedings of the 12th International symposium on chromaffin cell biology, 21-25 September, 2003.
Aunis D. and Metz-Boutigue M.H., (2000). Adv. Exp. Med. Biol. 482: 21-38.
Benjamin IJ et al (1989). Circ.Res. 65: 657-70.
Bolli R. et al Circ.Res. 1997; 81:42-52
Bretscher A et al (2000). Annu.Rev.Cell.Dev.Biol. 16: 113-143.
Brodde O.E. and Michel M.C., (1999). Pharm. Rev. 51: 651-690,
Ceconi C., et al., (2002). Eur. Heart J. 23: 967-974.
Cerra et al., (2005) Basic Res Cardiol 100:1-10.
Chein KR et al (1991).. FASEB 5: 3037-46.
Chrousos GP & Gold PW (1992). JAMA 267:1244-52.
Cohn JN et al (1984) N.Engl.J.Med. 311: 819-23.
Corti A., et al., (2002). Ann. N. Y.Acad. Sci. 971: 362-365.
Corti A., et al., (2004). Gen. Comp. End. 136: 217-224.
Depre C., (1998). Nucl. Med. Biol. 25: 711-713.
Esler M et al (1997). Am.J.Card. 80: 7L-14L.
Frayne J., et al., (1999). Cell. Tissue Res. 298(3): 415-23.
Gasparri A., et al., (1997). J. Biol. Chem. 272: 20835-20843.
Gattullo, D. et al (1999) Cardiovasc Res 42(1): 57-64.
Goldspink DF et al (2004). Exp.Physiol. 89: 407-16.
Goumon Y., et al., (1998). J. Biol. Chem. 273(45): 29847-56.
Goumon Y., et al., (2000). J. Biol. Chem. 275(49):38355-62.
Goumon Y., et al., (2004). J. Bio. Chem. 279: 13054-13064.
Grossman LA et al (1975). J.Tenn.Med.Assoc. 68: 903-4.
Helle K.B, et al., (2001). Curr . Med. Chem. 1: 119-140.
Helle K.B. and Angeletti R. H. (1998). In Kanno, T., et al (Eds) : The Adrenal Chromaffin Cell : Archetype
and Exemplar of Cellular Signaling in Secretory Control. Hokkaido University Press, Japan, pp 273-281.
Helle K.B., (2004). Biol. Rev. 79: 769-794.
Hengst U., et al., (2001). J. Biol. Chem. 276: 535-540.
Imbrogno S., et al., (2004). Gen. Comp. End. 139: 20-28.
Katada E., et al., (1996). Histochem. Cell. Biol. 105: 43-51.
Kieffer A.E., et al., (2003). FASEB J. 17: 776-778.
Koeslag, J.H., et al. (1997) J Endocrinol 154(2): 187-92.
Koeslag, J.H., et al. (1999) J Physiol 517 (3): 643-9.
Kratchmarova I., et al., (2002). Mol. Cell. Proteomics. 1(3):213-22.
Kroslak T., et al. (2001). J. Biol. Chem. 276: 39772-39778.
Kurtz TW & Morris RC (1987). Hypert. 10: 127-131.
Lawson CF, Downey JM, Card. Res. 1993; 27: 542-50
Lindpaintner K et al (1992). Hypert. 19: 428-430.
Lugardon K., et al., (2001). J. Biol. Chem. 276: 35875-35882.
Maget-Dana R., et al., (2002). Ann. N. Y. Acad. Sci. 971: 352-354.
Malan D., et al., (2003). J. Mol. Cell. Card. 35: 195-206.
Metz-Boutigue M.H., et al., (1993). Eur. J. Biochem. 217: 247-57.
Ojika K et al , (2000). Prog. Neurobiol. 60: 37-83.
Ojika K. and Appel S.H. (1984). Proc. Natl. Acad. Sci U S A. 81(8): 2567-71.
Ojika K., et al., (1994). Brain. Res. Dev. Brain. Res.79 (1): 1-9.
Pagliaro P. et al, Ital heart J. 2001; 2:660-8.
Parratt JR. , Vegh A. , Cardioscience1994; 5: 9-18
Raab W (1963)Rev.Can.Biol.22: 217-239.
Rockman M.V., et al., (2002). Nature 415: 206-212,
Saez JC et al (2003). Physiol.Rev. 83:1359-400.
Sasaki N. et al (2000) Circulation; 101:439-45
Saunders, B.D. et al (1998) Endocrinology 139(4):1835-43
Selye H (1936). J.Neuropsychiatry Clin.Neurosci. 10: 230-231.
•
•
•
•
•
•
•
•
•
•
•
•
•
•
Shizukuda Y. et al (1992) Card. Res.; 26:534-42
Shoentgen F. and Jollès P., (1995). FEBS Lett. 369(1): 22-6.
Steiner H. J, et al., (1990). J. Histochem. Cytochem. 38: 845-850.
Sutherland F. J. and Hearse D. J. (2000). Pharm. Res. 41: 613-27.
Takata Y & Kato H (1996). Life Sci. 58: 91-106.
Tan LB et al (2003). J. Cardiovasc. Pharmacol. 41:518-25.
Teerlink JR et al (1994). Circ.Res. 75: 105-13.
Tota B., et al., (2003). Reg. Pept. 114: 123-130.
Tota B., et al., (2004). Curr. Med. Chem.. 4: 195-201.
Tsugu Y., et al., (1998). Eur. J. Neurol. 5(6): 561-569.
Vallèe B. S., et al., (2003). J. Pept. Res. 61: 47-57.
Vegh A. et al Card. Res. 1991, 25:1051-3
Yamada et al. (1998). Pharmacol. Rev. 50: 723-760.
Yeung K., et al., (1999). Nature. 401(6749):173-7.
PIANO DI SPESA
Voce di spesa
Spesa in euro
Descrizione
Materiale
inventariabile
1000
COMPUTER per sistema di acquisizione dati (POWER LAB)
Materiale di consumo
e funzionamento
2200
Altro
1000
ANIMALI (RATTI NORMOTESI E IPERTESI), SOSTANZE CHIMICHE
E FARMACOLOGICHE, MATERIALE DI CONSUMO DI
LABORATORIO (VETRERIA, MISCELE GASSOSE, FERRI DA
DISSEZIONE, ETC.),
TRASFERTE NELL'AMBITO DEL PROGETTO, SPESA
PUBBLICAZIONE DATI,
PARTECIPAZIONE A CONGRESSO NAZIONALI E/O
INTERNAZIONALI PER LA DIVULAGAZIONE DEI RISULTATI
DELLA RICERCA
TOTALE
4200
Rende, lì 02/03/2006
Il Proponente del progetto
Dr. Tommaso Angelone
Il Direttore del Dip. Farmaco-Biologico
Prof. Marcello Maggiolini